Changes in Fish Taxonomy Affect Freshwater Biogeographical Regionalisations: Insights from Greece



Abstract
:1. Introduction
1.1. Fish Species Taxonomy Is Changing in Europe
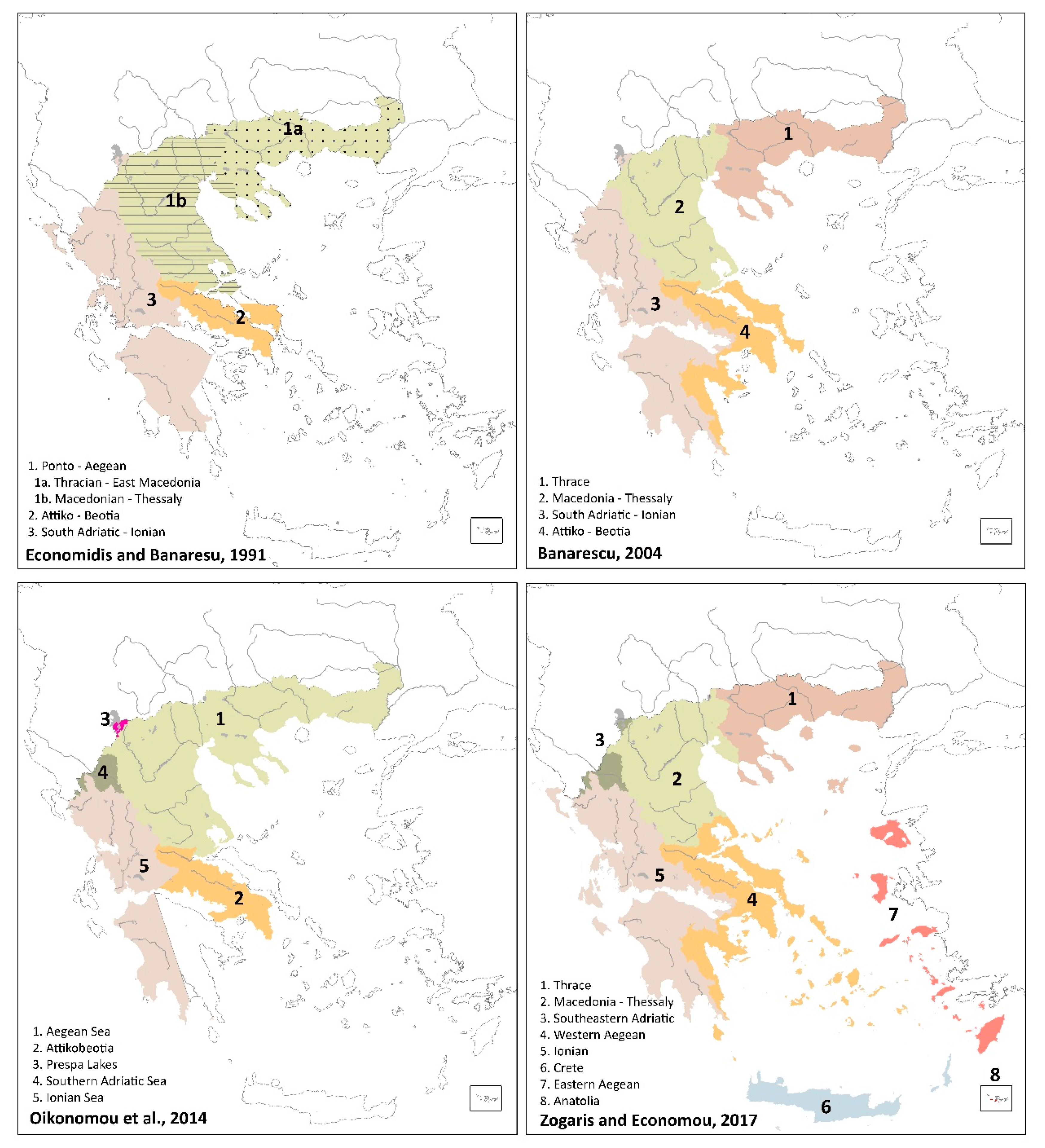
1.2. Biogeographical Delineations in Greece: Why Review Boundaries?
2. Materials and Methods
3. Results
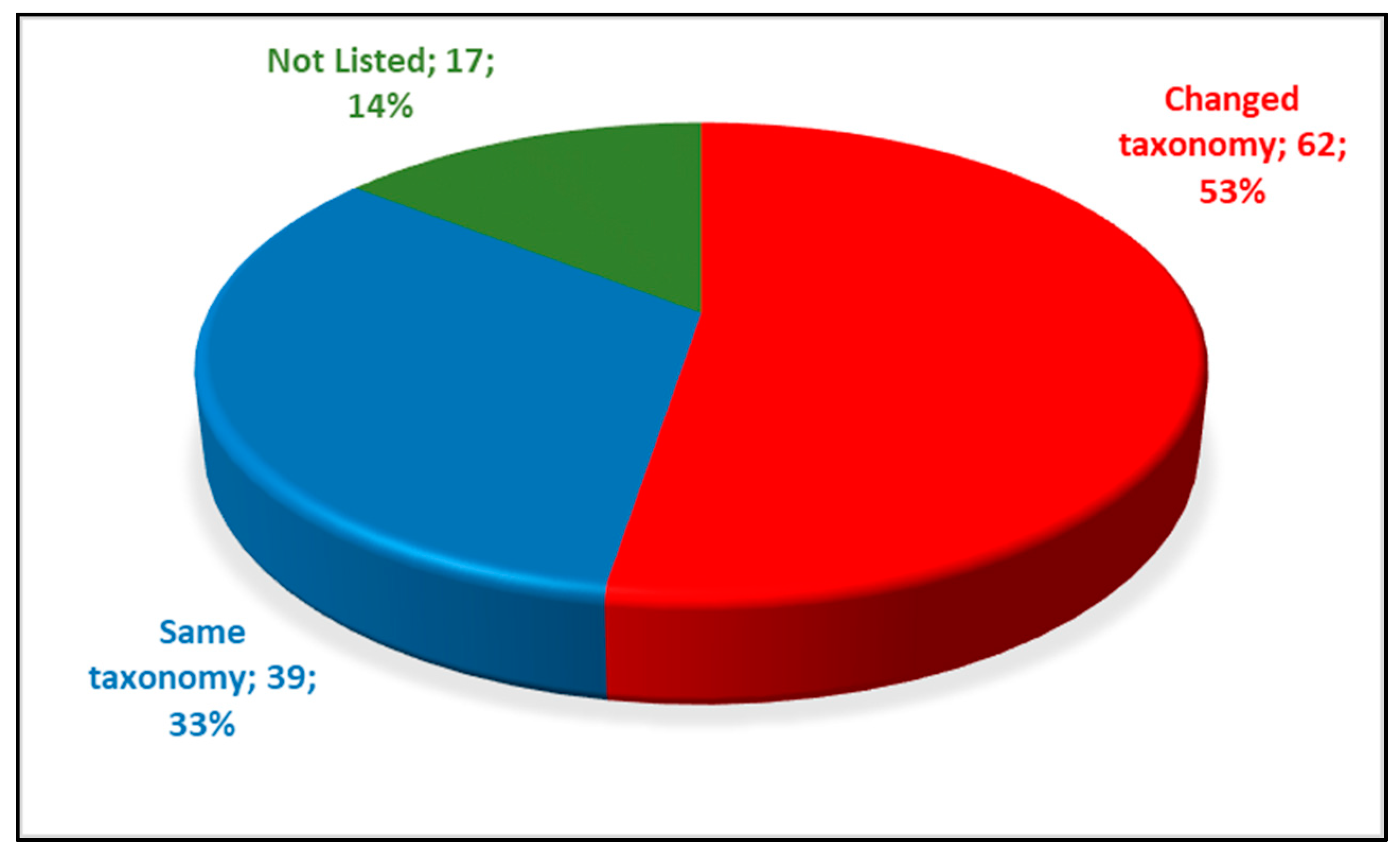
3.1. How Much Has the Taxonomy Changed?
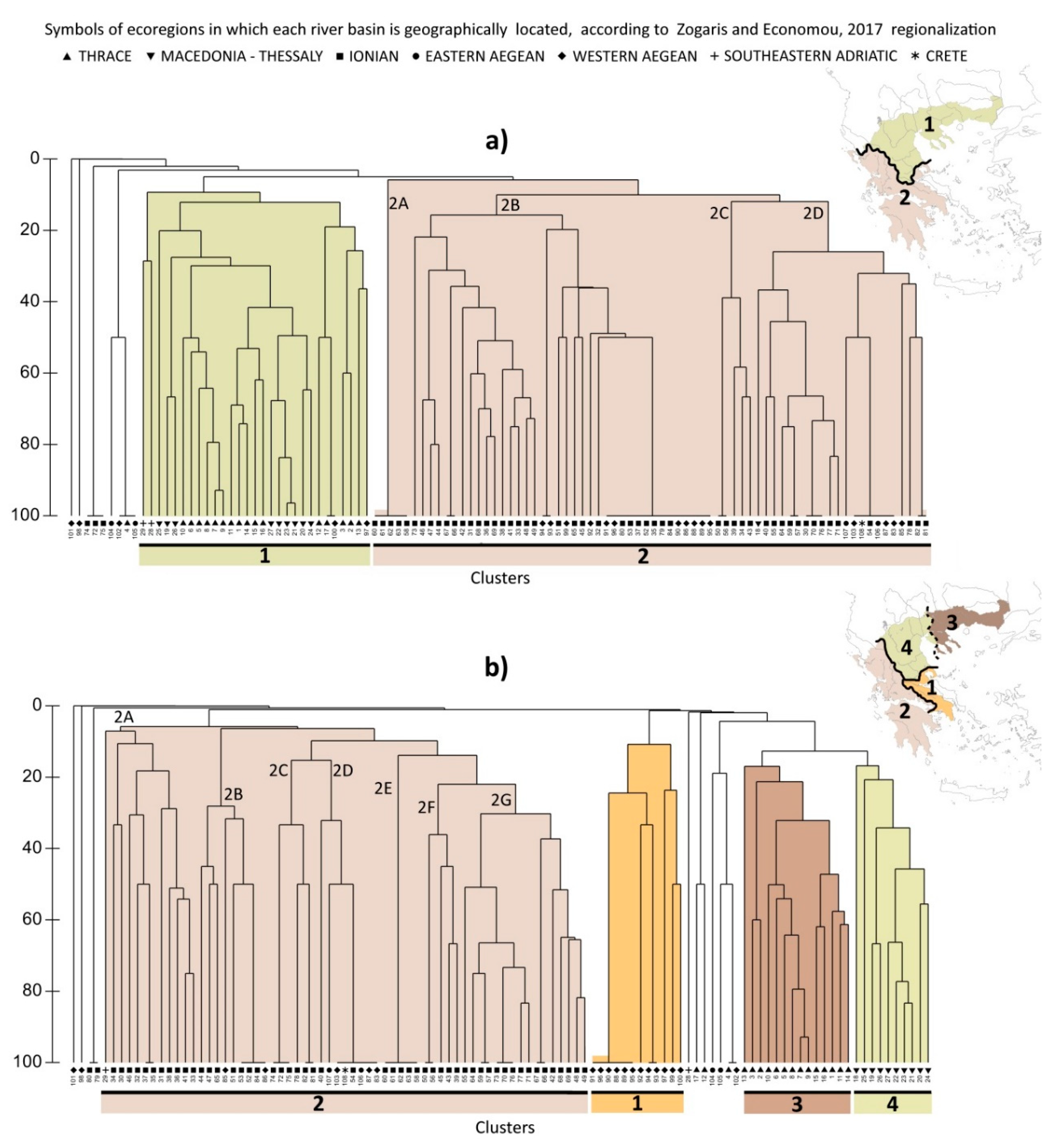
3.2. Different Dataset, Different Ichthyoregions
4. Discussion
4.1. Different Interpretations, Different Regionalisations
4.2. Biogeographical Numerical Taxonomy: The Devil Is in the Details
4.3. Beware of Anthropogenic Translocations and Faunal Homogenization
4.4. The Issue of Spatial Scale
4.5. Regionalisation Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Groves, C. Drafting a Conservation Blueprint: A Practitioner’s Guide to Planning for Biodiversity; Island Press: Washington, DC, USA, 2003; p. 480. [Google Scholar]
- Markovic, D.; Freyhof, J.; Wolter, C. Where Are All the Fish: Potential of Biogeographical Maps to Project Current and Future Distribution Patterns of Freshwater Species. PLoS ONE 2012, 7, 40530. [Google Scholar] [CrossRef] [PubMed]
- Elkins, D.; Sweat, S.C.; Kuhajda, B.R.; George, A.L.; Hill, K.S.; Wenger, S.J. Illuminating hotspots of imperiled aquatic biodiversity in the southeastern US. Glob. Ecol. Conserv. 2019, 19, e00654. [Google Scholar] [CrossRef]
- Von Humboldt, A. Essai sur la Géographie des Plantes: Accompagné d’un Tableau Physique des Régions Équinoxiales, Fondé sur des Mesures Exécutées, Depuis le Dixième Degré de Latitude Boréale Jusqu’au Dixième Degré de Latitude Australe, Pendant les Années 1799, 1800, 1801, 1802 et 1803 par Al. de Humboldt et A. Bonpland; Schoell: Paris, France, 1807; p. 155. [Google Scholar]
- Kreft, H.; Jetz, W. A framework for delineating biogeographical regions based on species distributions. J. Biogeogr. 2010, 37, 2029–2053. [Google Scholar] [CrossRef]
- Vilhena, D.A.; Antonelli, A. A network approach for identifying and delimiting biogeographical regions. Nat. Commun. 2015, 6, 6848. [Google Scholar] [CrossRef] [PubMed]
- Bernardo-Madrid, R.; Calatayud, J.; González-Suarez, M.; Rosvall, M.; Lucas, P.M.; Rueda, M.; Antonelli, A.; Revilla, E. Human activity is altering the world’s zoogeographical regions. Ecol. Lett. 2018, 287300. [Google Scholar] [CrossRef] [PubMed]
- Zogaris, S.; Economou, A.N.; Dimopoulos, P. Ecoregions in the Southern Balkans: Should Their Boundaries Be Revised? Environ. Manag. 2009, 43, 682–697. [Google Scholar] [CrossRef] [PubMed]
- Castro-Insua, A.; Gómez-Rodríguez, C.; Baselga, A. Dissimilarity measures affected by richness differences yield biased delimitations of biogeographic realms. Nat. Commun. 2018, 9, 5084. [Google Scholar] [CrossRef]
- Bunce, R.G.H.; Carey, P.D.; Elena-Rossello, R.; Orr, J.; Watkins, J.; Fuller, R. A comparison of different biogeographical classifications of Europe, Great Britain and Spain. J. Environ. Manag. 2002, 65, 121–134. [Google Scholar] [CrossRef]
- Petit, G. Remarques sur la distribution géographique des poissons d’eau douce des Balkans. C. R. Soc. Seances Soc. Biol. Fil. 1930, 59, 77–82. [Google Scholar]
- Reyjol, Y.; Hugueny, B.; Pont, D.; Bianco, P.G.; Beier, U.; Caiola, N.; Casals, F.; Cowx, I.; Economou, A.; Ferreira, T.; et al. Patterns in species richness and endemism of European freshwater fish. Glob. Ecol. Biogeogr. 2007, 16, 65–75. [Google Scholar] [CrossRef]
- Abell, R.; Thieme, M.L.; Revenga, C.; Bryer, M.; Kottelat, M.; Bogutskaya, N.; Coad, B.; Mandrak, N.; Balderas, S.C.; Bussing, W.; et al. Freshwater Ecoregions of the World: A New Map of Biogeographic Units for Freshwater Biodiversity Conservation. BioScience 2008, 58, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, C.R. Zoogeographic factors in relation to biological monitoring of fish. In Biological Monitoring of Fish; Hocutt, C.H., Stauffer, J.R., Eds.; D.C. Heath: Lexington, MA, USA, 1980; pp. 309–355. [Google Scholar]
- Bohlen, J.; Perdices, A.; Doadrio, I.; Economidis, P.S. Vicariance, colonisation, and fast local speciation in Asia Minor and the Balkans as revealed from the phylogeny of spined loaches (Osteichthyes; Cobitidae). Mol. Phylogenet. Evol. 2006, 39, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Matthews, W.J. Patterns in Freshwater Fish Ecology; Chapman & Hall: New York, NY, USA, 1998; p. 756. [Google Scholar]
- Leroy, B.; Dias, M.S.; Giraud, E.; Hugueny, B.; Jezequel, C.; Leprieur, F.; Oberdorff, T.; Tedesco, P.A. Global biogeographical regions of freshwater fish species. BioRxiv 2019, 319566. [Google Scholar] [CrossRef]
- Abell, R.A.; Olson, D.M.; Dinerstein, E.; Hurley, P.T.; Diggs, J.T.; Eichbaum, W.; Walters, S.; Wettengel, W.; Allnutt, T.; Loucks, C.J.; et al. Freshwater Ecoregions of North America: A Conservation Assessment; Island Press: Washington, DC, USA, 2000; p. 368. [Google Scholar]
- Ladle, R.; Whittaker, R.J. Conservation Biogeography; Wiley-Blackwell: Chichester, UK, 2011; p. 301. [Google Scholar]
- De Figueroa, J.M.T.; López-Rodríguez, M.J.; Fenoglio, S.; Sánchez-Castillo, P.; Fochetti, R. Freshwater biodiversity in the rivers of the Mediterranean Basin. Hydrobiologia 2013, 719, 137–186. [Google Scholar] [CrossRef]
- Tedesco, P.A.; Beauchard, O.; Bigorne, R.; Blanchet, S.; Buisson, L.; Conti, L.; Cornu, J.-F.; Dias, M.S.; Grenouillet, G.; Hugueny, B.; et al. A global database on freshwater fish species occurrence in drainage basins. Sci. Data 2017, 4, 170141. [Google Scholar] [CrossRef]
- Kottelat, M.; Freyhof, J. Handbook of European Freshwater Fishes; Kottelat: Cornol, Switzerland; Freyhof: Berlin, Germany, 2007; p. 646. [Google Scholar]
- Economou, A.; Giakoumi, S.; Vardakas, L.; Barbieri, R.; Stoumboudi, M.; Zogaris, S. The freshwater ichthyofauna of Greece—An update based on a hydrographic basin survey. Mediterr. Mar. Sci. 2007, 8, 91–166. [Google Scholar] [CrossRef]
- Kottelat, M. An Heuristic Checklist of the Freshwater Fishes of Europe (Exclusive of Former USSR): With an Introduction for Non-Systematists and Comments on Nomenclature and Conservation; Biologia: Bratislava, Slovakia, 1997; p. 271. [Google Scholar]
- Kottelat, M. Systematics, species concepts and the conservation of freshwater fish diversity in Europe. Ital. J. Zool. 1998, 65, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Maitland, P.S.; Linsell, K. Guide to Freshwater Fish of Britain and Europe; Hamlyn: London, UK, 2006; p. 256. [Google Scholar]
- Blanc, M.; Bănărescu, P.M.; Gaudet, J.; Hureau, J. European Inland Water Fish: A Multilingual Catalogue; Fishing News (Books) Ltd.: London, UK, 1971. [Google Scholar]
- Page, L.M. Handbook of European Freshwater Fishes. Copeia 2008, 2008, 725–727. [Google Scholar] [CrossRef]
- Freyhof, J.; Brooks, E. European Red List of Freshwater Fishes; Publications Office of the European Union: Luxembourg, 2011; p. 70. [Google Scholar]
- FEOW. Freshwater Ecoregions of the World—Conservation Science Program. Available online: http://www.feow.org (accessed on 21 June 2019).
- Barbieri, R.; Zogaris, S.; Kalogianni, E.; Stoumboudi, M.; Chatzinikolaou, Y.; Giakoumi, S.; Kapakos, Y.; Kommatas, D.; Koutsikos, N.; Tachos, V.; et al. Freshwater Fishes and Lampreys of Greece: An Annotated Checklist; Monographs on Marine Sciences No. 8; Hellenic Centre for Marine Research: Athens, Greece, 2015. [Google Scholar]
- Economou, A.N.; Zogaris, S.; Vardakas, L.; Koutsikos, N.; Chatzinikolaou, Y.; Kommatas, D.; Kapakos, Y.; Giakoumi, S.; Oikonomou, E.; Tachos, V. Developing policy-relevant river fish monitoring in Greece: Insights from a nation-wide survey. Mediterr. Mar. Sci. 2016, 17, 302. [Google Scholar] [CrossRef]
- Naseka, A. Zoogeographical freshwater divisions of the Caucasus as a part of the West Asian Transitional Region. Proc. ZIN 2010, 314, 469–492. [Google Scholar]
- Iliadou, E.; Kallimanis, A.S.; Dimopoulos, P.; Panitsa, M. Comparing the two Greek archipelagos plant species diversity and endemism patterns highlight the importance of isolation and precipitation as biodiversity drivers. J. Boil. Res. 2014, 21, 16. [Google Scholar] [CrossRef] [PubMed]
- Poulakakis, N.; Kapli, P.; Lymberakis, P.; Trichas, A.; Vardinoyiannis, K.; Sfenthourakis, S.; Mylonas, M. A review of phylogeographic analyses of animal taxa from the Aegean and surrounding regions. J. Zool. Syst. Evol. Res. 2015, 53, 18–32. [Google Scholar] [CrossRef]
- Sfenthourakis, S.; Triantis, K.A. The Aegean archipelago: A natural laboratory of evolution, ecology and civilisations. J. Boil. Res. 2017, 24, 373. [Google Scholar] [CrossRef] [PubMed]
- Triantis, K.A.; Kougioumoutzis, K.; Legakis, A.; Anastasiou, I.; Andriopoulos, P.; Georgiadis, C.; Lymberakis, P.; Oikonomou, A.; Probonas, N.; Proios, K.; et al. The zoogeographic regions of the Aegean Sea: A multi-taxon approach. In Biogeography and Biodiversity of the Aegean. In Honour of Prof. Moysis Mylonas; Sfenthourakis, S., Pafilis, P., Parmakelis, A., Poulakakis, N., Triantis, K.A., Eds.; Broken Hill Publishers Ltd.: Nicosia, Cyprus, 2018; pp. 279–290. [Google Scholar]
- Zogaris, S.; Economou, A.N. The Biogeographic Characteristics of the River Basins of Greece: Evolution, Current Status and Perspectives; Skoulikidis, N.T., Dimitriou, E., Karaouzas, I., Eds.; Springer-Verlag GmbH Germany: Athens, Greece, 2017; pp. 53–95. [Google Scholar]
- Buj, I.; Šanda, R.; Zogaris, S.; Freyhof, J.; Geiger, M.F.; Vukić, J. Cryptic diversity in Telestes pleurobipunctatus (Actinopterygii; Leuciscidae) as a consequence of historical biogeography in the Ionian Freshwater Ecoregion (Greece, Albania). Hydrobiologia 2019, 835, 147–163. [Google Scholar] [CrossRef]
- Stephanidis, A. Freshwater fish of Attiko-Beotia. Bull. Phys. Sci. Fasc. 1939, 50–51, 49–60. [Google Scholar]
- Economidis, P.S.; Banarescu, P.M. The Distribution and Origins of Freshwater Fishes in the Balkan Peninsula, Especially in Greece. Int. Rev. Hydrobiol. 1991, 76, 257–284. [Google Scholar] [CrossRef]
- Economidis, P.S. Check List of Freshwater Fishes of Greece: Recent Status of Threats and Protection; Hellenic Society for the Protection of Nature: Athens, Greece, 1991. [Google Scholar]
- Oikonomou, A.; Leprieur, F.; Leonardos, I.D. Biogeography of freshwater fishes of the Balkan Peninsula. Hydrobiologia 2014, 738, 205–220. [Google Scholar] [CrossRef]
- Illies, J. Limnofauna Europaea; Gustav Fischer Verlag: Stuttgart, Germany, 1978; p. 532. [Google Scholar]
- Griffiths, D. Pattern and process in the ecological biogeography of European freshwater fish. J. Anim. Ecol. 2006, 75, 734–751. [Google Scholar] [CrossRef]
- Maurakis, E.; Economidis, P.S. Reconstructing biogeographical relationships of river drainages in Peloponessos, Greece using distributions of freshwater cyprinid fishes. BIOS 2001, 6, 125–132. [Google Scholar]
- Maurakis, E.; Pritchard, M.; Economidis, P. Historical relationships of mainland river drainages in Greece. BIOS 2001, 6, 109–124. [Google Scholar]
- Carrizo, S.F.; Kapusi, F.; Szabolcs, M.; Kasperidus, H.D.; Scholz, M.; Cardoso, A.C.; Darwall, W.; Lengyel, S.; Markovic, D.; Freyhof, J.; et al. Critical catchments for freshwater biodiversity conservation in Europe: Identification, prioritisation and gap analysis. J. Appl. Ecol. 2017, 54, 1209–1218. [Google Scholar] [CrossRef]
- Koutsikos, N.; Zogaris, S.; Vardakas, L.; Tachos, V.; Kalogianni, E.; Sanda, R.; Chatzinikolaou, Y.; Giakoumi, S.; Economidis, P.; Economou, A. Recent contributions to the distribution of the freshwater ichthyofauna in Greece. Mediterr. Mar. Sci. 2012, 13, 268–277. [Google Scholar] [CrossRef]
- Zogaris, S.; Tachos, V.; Economou, A.; Chatzinikolaou, Y.; Koutsikos, N.; Schmutz, S. A model-based fish bioassessment index for Eastern Mediterranean rivers: Application in a biogeographically diverse area. Sci. Total Environ. 2018, 622, 676–689. [Google Scholar] [CrossRef]
- European Committee for Standardization (CEN). Water Quality–Sampling of Fish with Electricity. European Standard; European Committee for Standardization: Brussels, Belgium, 2003; p. 20. [Google Scholar]
- Koutsikos, N.; Zogaris, S.; Vardakas, L.; Kalantzi, O.-I.; Dimitriou, E.; Economou, A.N. Tracking non-indigenous fishes in lotic ecosystems: Invasive patterns at different spatial scales in Greece. Sci. Total Environ. 2019, 659, 384–400. [Google Scholar] [CrossRef]
- Barbieri, R.; Vukić, J.; Šanda, R.; Kapakos, Y.; Zogaris, S. Alburnoides economoui, a new species of spirlin from Central Greece and redescription of Alburnoides thessalicus (Actinopterygii: Cyprinidae). Boilogia 2017, 72, 1075–1088. [Google Scholar] [CrossRef]
- Economidis, P.S. Catalogue of the fish of Greece (In Greek). Hell. Oceanol. Limnol. 1973, 11, 421–598. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial, PRIMER—E: Plymouth; Plymouth Marine Laboratory: Plymouth, UK, 2006. [Google Scholar]
- Clarke, K.; Green, R. Statistical design and analysis for a ‘biological effects’ study. Mar. Ecol. Prog. Ser. 1988, 46, 213–226. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Austral Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Banarescu, P.M. Distribution Pattern of the Aquatic Fauna of the Balkan Peninsula. In Balkan Biodiversity: Pattern and Process in the European Hotspot; Griffiths, H.I., Krystufek, B., Reed, J.M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; pp. 203–217. [Google Scholar]
- Bianco, P.G. Potential role of the palaeohistory of the Mediterranean and Paratethys basins on the early dispersal of Euro-Mediterranean freshwater fishes. Ichthyol. Explor. Freshw. 1990, 1, 167–184. [Google Scholar]
- Skoulikidis, N.T.; Economou, A.; Gritzalis, K.; Zogaris, S. Rivers of the Balkans. In Rivers of Europe; Tockner, K., Uehlinger, U., Robinson, C.T., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2009; pp. 421–466. [Google Scholar]
- Ficetola, G.F.; Mazel, F.; Thuiller, W. Global determinants of zoogeographical boundaries. Nat. Ecol. Evol. 2017, 1, 89. [Google Scholar] [CrossRef]
- Bloomfield, N.J.; Knerr, N.; Encinas-Viso, F. A comparison of network and clustering methods to detect biogeographical regions. Ecography 2018, 41, 1–10. [Google Scholar] [CrossRef]
- Dagosta, F.C.P.; De Pinna, M. Biogeography of Amazonian fishes: Deconstructing river basins as biogeographic units. Neotropical Ichthyol. 2017, 15. [Google Scholar] [CrossRef]
- Geiger, M.F.; Herder, F.; Monaghan, M.T.; Almada, V.; Barbieri, R.; Bariche, M.; Berrebi, P.; Bohlen, J.; Casal-López, M.; Delmastro, G.B.; et al. Spatial heterogeneity in the Mediterranean Biodiversity Hotspot affects barcoding accuracy of its freshwater fishes. Mol. Ecol. Resour. 2014, 14, 1210–1221. [Google Scholar] [CrossRef] [Green Version]
- Nipperess, D.A.; Faith, D.P.; Barton, K. Resemblance in phylogenetic diversity among ecological assemblages. J. Veg. Sci. 2010, 21, 809–820. [Google Scholar] [CrossRef]
- Abell, R.; Thieme, M.; Dinerstein, E.; Olson, D. A Sourcebook for Conducting Biological Assessments and Developing Biodiversity Visions for Ecoregion Conservation. Volume II: Freshwater Ecoregions; World Wildlife Fund: Washington, DC, USA, 2002. [Google Scholar]
- Blondel, J. On humans and wildlife in Mediterranean islands. J. Biogeogr. 2008, 35, 509–518. [Google Scholar] [CrossRef]
- Viñuela Rodríguez, N. Phylogeny and Phylogeography of the Cyprinid Fish Genus Pelasgus (Teleostei: Cyprinidae). Master’s Thesis, Charles University in Prague, Prague, Czech Republic, 2016. [Google Scholar]
- Vanhove, M.P.M.; Economou, A.N.; Zogaris, S.; Larmuseau, M.H.D.; Giakoumi, S.; Kalogianni, E.; Volckaert, F.A.M.; Huyse, T. Phylogenetics and biogeography of the Balkan ‘sand gobies’ (Teleostei: Gobiidae): Vulnerable species in need of taxonomic revision. Boil. J. Linn. Soc. 2011, 105, 73–91. [Google Scholar] [CrossRef]
- Zogaris, S.; Maclaine, J.; Koutsikos, N.; Chatzinikolaou, Y. Does the river blenny Salaria fluviatilis (Asso, 1801) (Actinopterygii: Perciformes) still survive on the Mediterranean island of Cyprus? J. Nat. Hist. 2015, 49, 347–357. [Google Scholar] [CrossRef]
- Foufopoulos, J.; Ives, A.R. Reptile Extinctions on Land-Bridge Islands: Life-History Attributes and Vulnerability to Extinction. Am. Nat. 1999, 153, 1–25. [Google Scholar] [CrossRef]
- Mylonas, M. The influence of man: A special problem in the study of the zoogeography of terrestrial molluscs on the Aegean islands. In World-Wide Snails: Biogeographical Studies on Non-Marine Mollusca. Brill/Backhuys, Leiden; Solem, A., van Bruggen, A.C., Eds.; E. J. Brill: Leiden, The Netherlands, 1984; pp. 249–259. [Google Scholar]
- Lymberakis, P.; Poulakakis, N. Internationale Revue der gesamten Hydrobiologie und Hydrographie. Diversity 2010, 2, 233–255. [Google Scholar] [CrossRef]
- Kyriazi, P.; Kornilios, P.; Nagy, Z.T.; Poulakakis, N.; Kumlutaş, Y.; Ilgaz, Ç.; Avcı, A.; Göçmen, B.; Lymberakis, P. Comparative phylogeography reveals distinct colonization patterns of Cretan snakes. J. Biogeogr. 2013, 40, 1143–1155. [Google Scholar] [CrossRef]
- Masseti, M. The long-time redefinition of the non-volant vertebrate horizons of the Eastern Mediterranean islands. In Biogeography and Biodiversity of the Aegean. In Honour of Prof. Moysis Mylonas; Pafilis, P., Parmakelis, A., Poulakakis, N., Triantis, K.A., Eds.; Broken Hill Publishers Ltd.: Nicosia, Cyprus, 2018; pp. 191–222. [Google Scholar]
- Olden, J.D.; Kennard, M.J.; Leprieur, F.; Tedesco, P.A.; Winemiller, K.O.; García-Berthou, E. Conservation biogeography of freshwater fishes: Recent progress and future challenges. Divers. Distrib. 2010, 16, 496–513. [Google Scholar] [CrossRef]
- Thieme, M.L.; Abell, R.; Burgess, N.; Lehner, B.; Dinerstein, E.; Olson, D. Freshwater Ecoregions of Africa and Madagascar: A Conservation Assessment; Island Press: Washington, DC, USA, 2005; p. 483. [Google Scholar]
- Higgins, J.V.; Bryer, M.T.; Khoury, M.L.; Fitzhugh, T.W. A Freshwater Classification Approach for Biodiversity Conservation Planning. Conserv. Boil. 2005, 19, 432–445. [Google Scholar] [CrossRef]
- Filipe, A.F.; Araújo, M.B.; Doadrio, I.; Angermeier, P.L.; Collares-Pereira, M.J.; Collares-Pereira, M.J.; Collares-Pereira, M.J. Biogeography of Iberian freshwater fishes revisited: The roles of historical versus contemporary constraints. J. Biogeogr. 2009, 36, 2096–2110. [Google Scholar] [CrossRef]
- Nel, J.L.; Roux, D.J.; Abell, R.; Ashton, P.J.; Cowling, R.M.; Higgins, J.V.; Thieme, M.; Viers, J.H. Progress and challenges in freshwater conservation planning. Aquat. Conserv. Mar. Freshw. Ecosyst. 2009, 19, 474–485. [Google Scholar] [CrossRef]
- Darwall, W.; Carrizo, S.; Numa, C.; Barrios, V.; Freyhof, J.; Smith, K. Freshwater Key Biodiversity Areas in the Mediterranean Basin Hotspot: Informing Species Conservation and Development Planning in Freshwater Ecosystems; IUCN: Cambridge, UK; Malaga, Spain, 2014; p. 100. [Google Scholar]
- Posadas, P.; Crisci, J.; Katinas, L. Historical biogeography: A review of its basic concepts and critical issues. J. Arid. Environ. 2006, 66, 389–403. [Google Scholar] [CrossRef]
- Carstensen, D.W.; Lessard, J.-P.; Holt, B.G.; Borregaard, M.K.; Rahbek, C. Introducing the biogeographic species pool. Ecography 2013, 36, 1310–1318. [Google Scholar] [CrossRef] [Green Version]
- Hoegh-Guldberg, O.; Hughes, L.; McIntyre, S.; Lindenmayer, D.B.; Parmesan, C.; Possingham, H.P.; Thomas, C.D. ECOLOGY: Assisted Colonization and Rapid Climate Change. Science 2008, 321, 345–346. [Google Scholar] [CrossRef]
- Tockner, K.; Pusch, M.; Gessner, J.; Wolter, C.; Gessner, J. Domesticated ecosystems and novel communities: Challenges for the management of large rivers. Ecohydrol. Hydrobiol. 2011, 11, 167–174. [Google Scholar] [CrossRef]
- Gozlan, R.E.; Karimov, B.K.; Zadereev, E.; Kuznetsova, D.; Brucet, S. Status, trends, and future dynamics of freshwater ecosystems in Europe and Central Asia. Inland Waters 2019, 9, 78–94. [Google Scholar] [CrossRef]
- Vanhove, M.P.; Kovačić, M.; Zogaris, S. A distinct island population of threatened freshwater fish: To split or lump? Hydrobiologia 2016, 777, 79–93. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
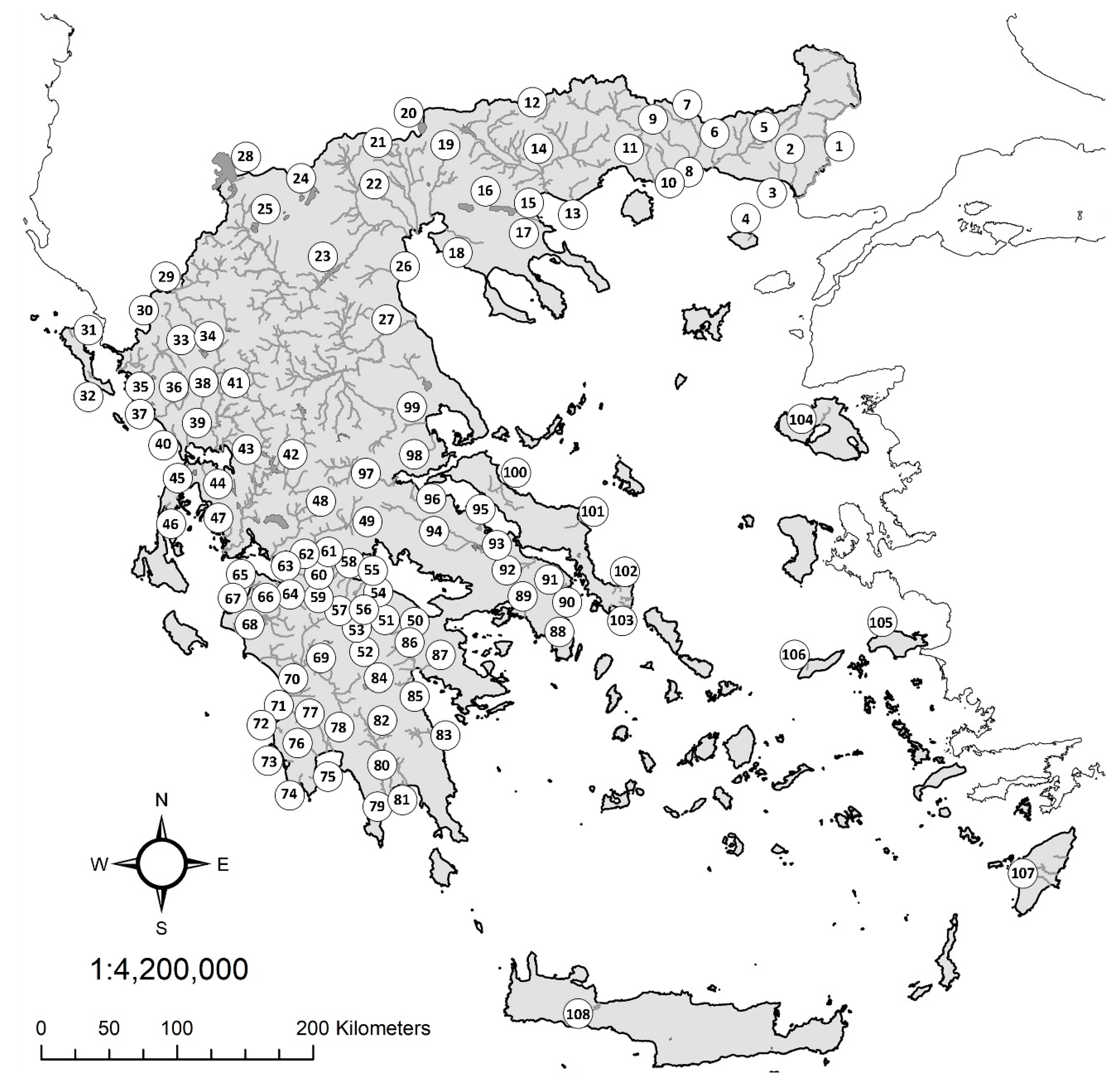
| 1. Evros | 23. Aliakmon | 45. Voulkaria | 67. Kotychi | 89. Kifissos (Attιki) |
| 2. Loutros | 24. Vegoritis | 46. Lefkada | 68. Pinios (Peloponnese) | 90. Rafina |
| 3. Apokrimno | 25. Kastoria | 47. Astakos | 69. Alfios | 91. Marathon |
| 4. Samothraki | 26. Mavroneri | 48. Evinos | 70. Neda | 92. Assopos (Beotia) |
| 5. Filiouri | 27. Pinios (Thessaly) | 49. Mornos | 71. Peristeras | 93. Yliki |
| 6. Bospos | 28. Prespa | 50. Assopos Pel | 72. Filiatrino | 94. Kifissos Beo |
| 7. Kompsatos | 29. Aoos | 51. Stymphalia | 73. Yannousagas | 95. Larymna |
| 8. Vistonis | 30. Zaravina | 52. Kandila | 74. Minagiotiko | 96. Platanias |
| 9. Kossinthos | 31. Kerkyra | 53. Feneos | 75. Kleisouraiiko | 97. Sperchios |
| 10. Laspias | 32. Messagis | 54. Krios | 76. Velika | 98. Sourporema |
| 11. Nestos | 33. Kalamas | 55. Krathis | 77. Pamissos | 99. Cholorema |
| 12. Nevrokopi | 34. Pamvotis | 56. Tsivlos | 78. Aris | 100. Kireas |
| 13. Marmaras | 35. Paramythia | 57. Vouraikos | 79. Smynous | 101. Manikiotiko |
| 14. Strymon | 36. Acheron | 58. Keronitis | 80. Ardeli | 102. Dimosaris |
| 15. Rihios | 37. Kalodiki | 59. Selinous | 81. Vassilopotamos | 103. Rigia |
| 16. Volvi | 38. Louros | 60. Meganitis | 82. Evrotas | 104. Lesvos |
| 17. Mavrolakas | 39. Ziros | 61. Phoenix | 83. Dafnonas | 105. Samos |
| 18. Anthemountas | 40. Arethoua | 62. Volinaios | 84. Taka | 106. Ikaria |
| 19. Gallikos | 41. Arachthos | 63. Glafkos | 85. Lerni | 107. Rhodos |
| 20. Doiran | 42. Acheloos | 64. Piros | 86. Erassinos Argolida | 108. Kourna |
| 21. Axios | 43. Vouvos | 65. Prokopos | 87. Nea Kios-Nafplion | |
| 22. Loudias | 44. Vlychos | 66. Vergas | 88. Erassinos Vravrona |
| Ichthyoregions | 3 | 4 | 2 | 1 |
|---|---|---|---|---|
| 3 | ||||
| 4 | 0.64 (0.1%) | |||
| 2 | 0.41 (0.1%) | 0.40 (0.1%) | ||
| 1 | 0.81 (0.1%) | 0.75 (0.1%) | 0.50 (0.1%) |
| Ionian Subclusters | 2G | 2A | 2D | 2C | 2F | 2B | 2E |
|---|---|---|---|---|---|---|---|
| 2G | |||||||
| 2A | 0.72 (0.1%) | ||||||
| 2D | 0.81 (0.1%) | 0.73 (0.1%) | |||||
| 2C | 0.82 (0.1%) | 0.83 (0.1%) | 0.82 (0.2%) | ||||
| 2F | 0.72 (0.1%) | 0.60 (0.1%) | 1.00 (0.3%) | 1.00 (0.2%) | |||
| 2B | 0.87 (0.1%) | 0.75 (0.1%) | 0.93 (0.1%) | 0.98 (0.1%) | 0.971 (0.1%) | ||
| 2E | 0.81 (0.1%) | 0.68 (0.1%) | 1.00 (0.1% | 1.00 (0.2%) | 1.00 (0.8%) | 1.00 (0.1%) |
| Old Taxonomy | Contrib% | Current Taxonomy | Contrib% |
|---|---|---|---|
| 1. “Northern Aegean” (31.70 %) | 1. Western Aegean (45.96%) | ||
| Leusiscus cephalus | 17.19 | Pelasgus marathonicus | 90.06 |
| Barbus cyclolepis | 12.00 | 2. Ionian (20.52%) | |
| Rhodeus sericeus | 9.40 | Salaria fluviatilis | 31.71 |
| Gobio gobio | 6.22 | Barbus peloponnesius | 17.60 |
| Cyprinus carpio | 5,76 | Squalius peloponensis | 14.49 |
| 2. “Greater Ionian” (27.88%) | Pelasgus stymphalicus | 13.51 | |
| Pseudophoxinus stymphalicus | 43.90 | Tropidophoxinellus spartiaticus | 5.29 |
| Salaria fluviatilis | 19.57 | 3. Macedonia–Thessaly (50.65%) | |
| Leusiscus cephalus | 14.87 | Squalius vardarensis | 15.01 |
| Barbus peloponnesius | 10.82 | Barbus balcanicus | 8.87 |
| Phoxinellus pleurobipunctatus | 3.07 | Cobitis vardarensis | 7.92 |
| Rhodeus meridionalis | 7.01 | ||
| Cyprinus carpio | 5.47 | ||
| 4. Thrace (48.82%) | |||
| Squalius orpheus | 16.25 | ||
| Cobitis strumicae | 13.09 | ||
| Rhodeus amarus | 10.30 | ||
| Gobio bulgaricus | 6.89 | ||
| Cyprinus carpio | 6.49 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vavalidis, T.; Zogaris, S.; Economou, A.N.; Kallimanis, A.S.; Bobori, D.C. Changes in Fish Taxonomy Affect Freshwater Biogeographical Regionalisations: Insights from Greece. Water 2019, 11, 1743. https://doi.org/10.3390/w11091743
Vavalidis T, Zogaris S, Economou AN, Kallimanis AS, Bobori DC. Changes in Fish Taxonomy Affect Freshwater Biogeographical Regionalisations: Insights from Greece. Water. 2019; 11(9):1743. https://doi.org/10.3390/w11091743
Chicago/Turabian StyleVavalidis, Theocharis, Stamatis Zogaris, Alcibiades N. Economou, Athanasios S. Kallimanis, and Dimitra C. Bobori. 2019. "Changes in Fish Taxonomy Affect Freshwater Biogeographical Regionalisations: Insights from Greece" Water 11, no. 9: 1743. https://doi.org/10.3390/w11091743
APA StyleVavalidis, T., Zogaris, S., Economou, A. N., Kallimanis, A. S., & Bobori, D. C. (2019). Changes in Fish Taxonomy Affect Freshwater Biogeographical Regionalisations: Insights from Greece. Water, 11(9), 1743. https://doi.org/10.3390/w11091743