Benthic Diatom Communities in Urban Streams and the Role of Riparian Buffers
 ,
,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- Can the influence of forested riparian buffers on benthic diatom communities be disentangled from other key drivers in urban streams? We hypothesized that the effect of riparian buffers would be weak due to overwhelming influences of catchment-wide urbanization causing degraded water quality and altered hydrogeomorphology.
- Can forested riparian buffers mitigate impacts of urbanization on diatom communities? We hypothesized that the provision of shading might have positive influences on biodiversity by reducing abundances of dominant, stress-tolerant diatoms and increasing the presence of rare species with ecological preferences for shaded conditions.
- When do forested riparian buffers contribute the most to improved stream ecological status? Here, we hypothesized that any change in diatom communities conferred by improved riparian conditions (i.e., forested buffers) would be contingent on the level of upstream impacts. Specifically, we predicted that positive effects would weaken with increasing catchment degradation, reflecting limits induced by poor water quality and hydrogeomorphological changes.
2. Materials and Methods
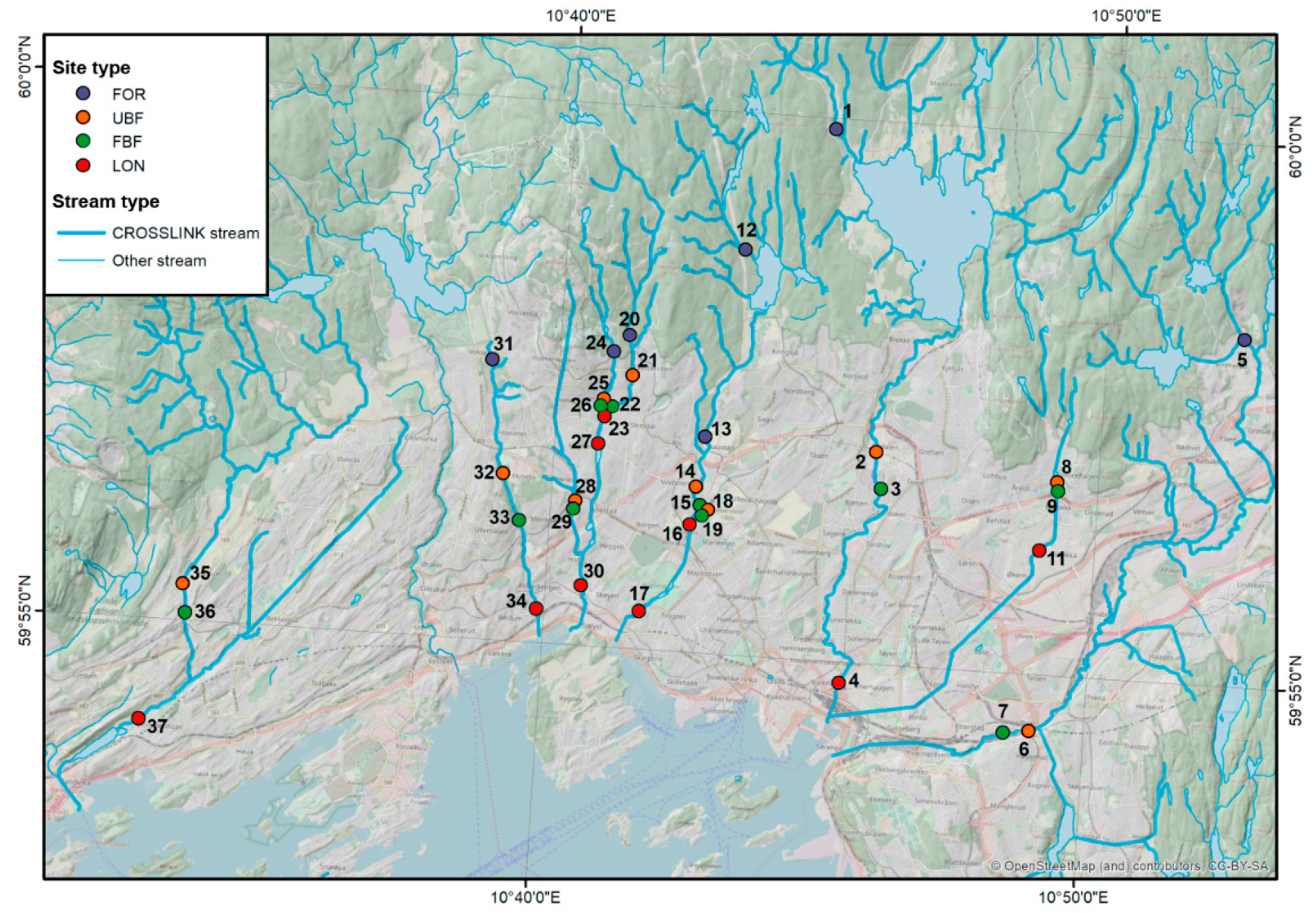
2.1. Study Design and Sites
2.2. Water Quality
2.3. Instream Habitat
2.4. Riparian Habitat and Catchment Land Uses
2.5. Diatoms
2.6. Data Analysis
3. Results
3.1. Description of Diatom Communities
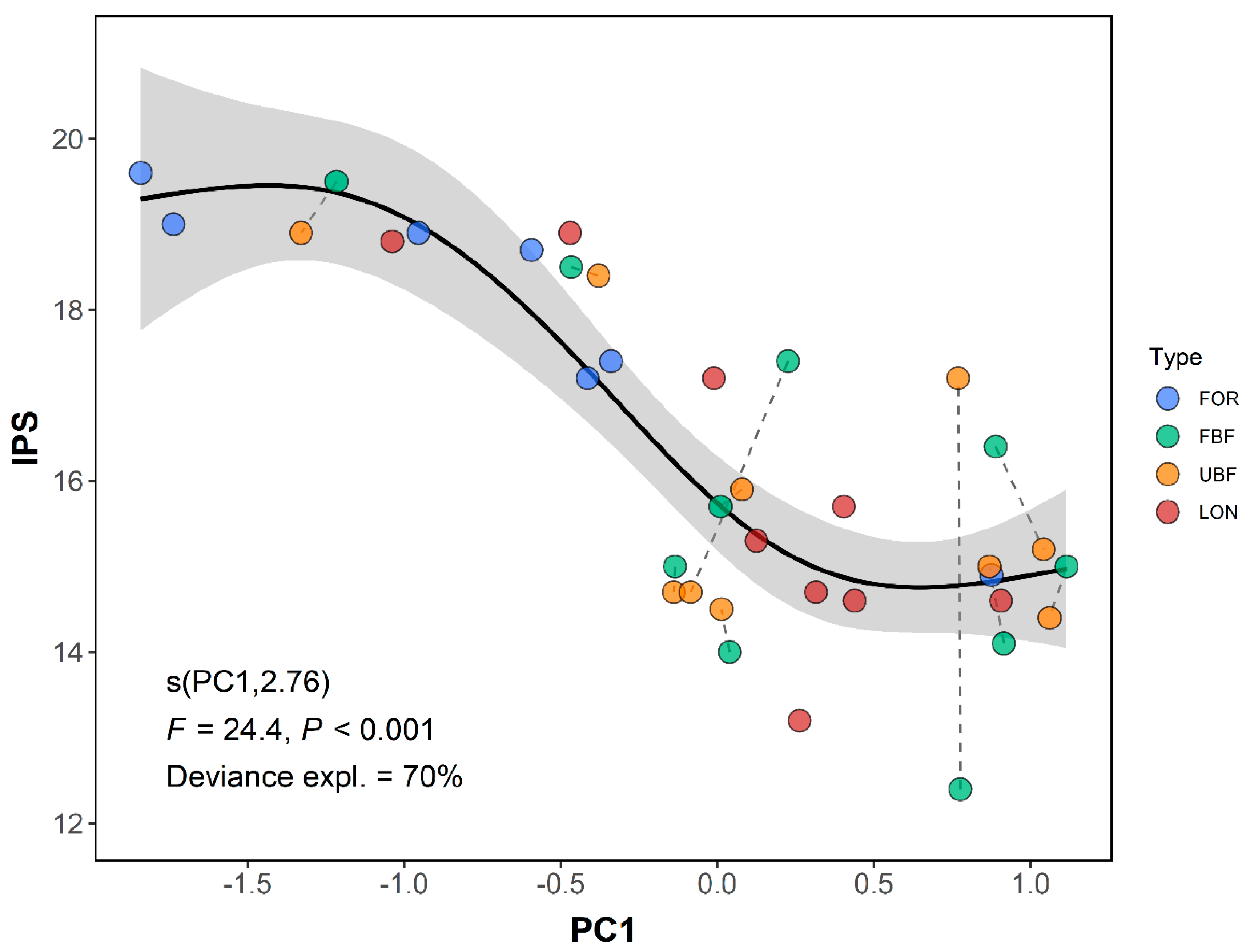
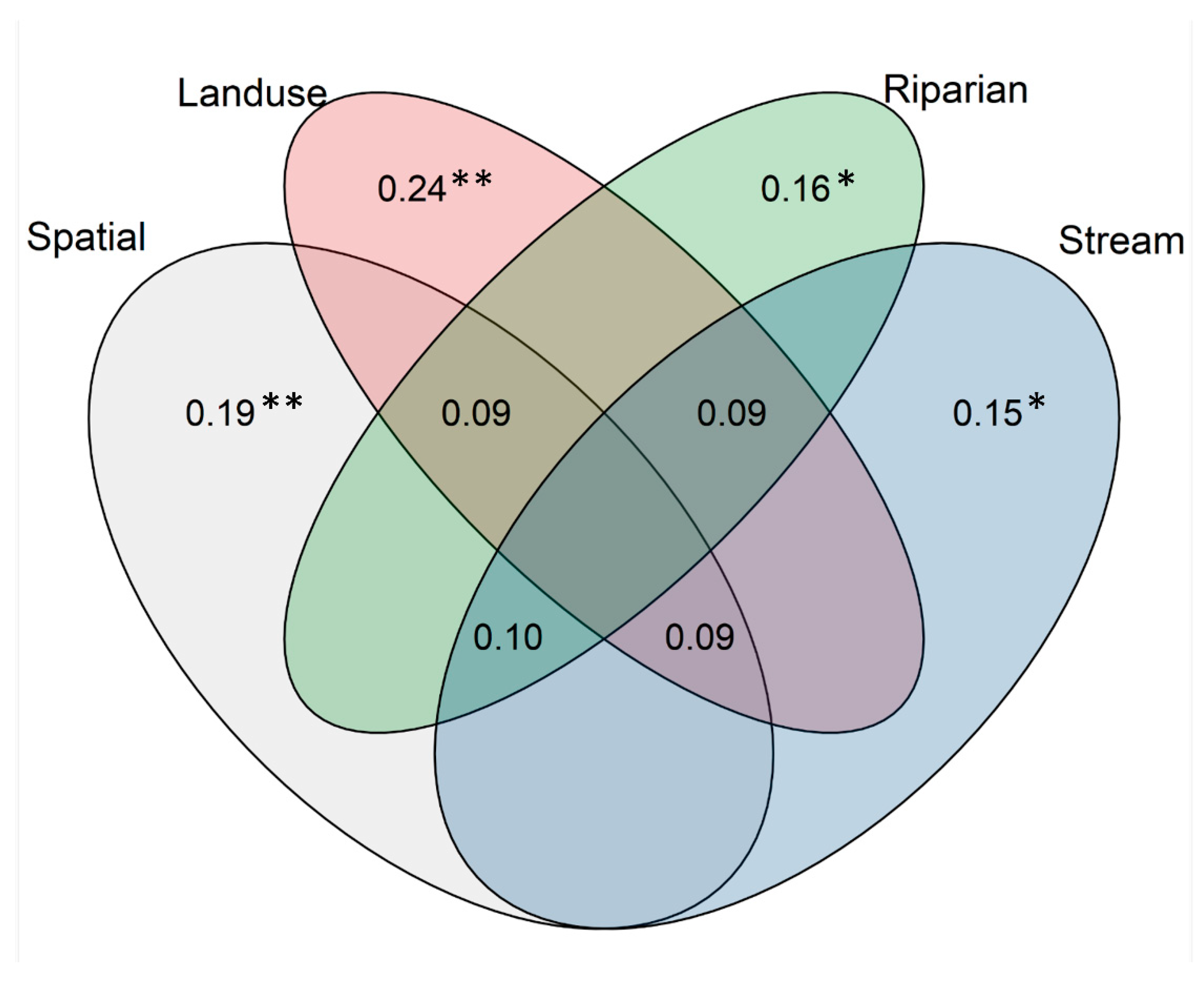
3.2. Drivers of Diatom Community Structure in the Oslo Basin
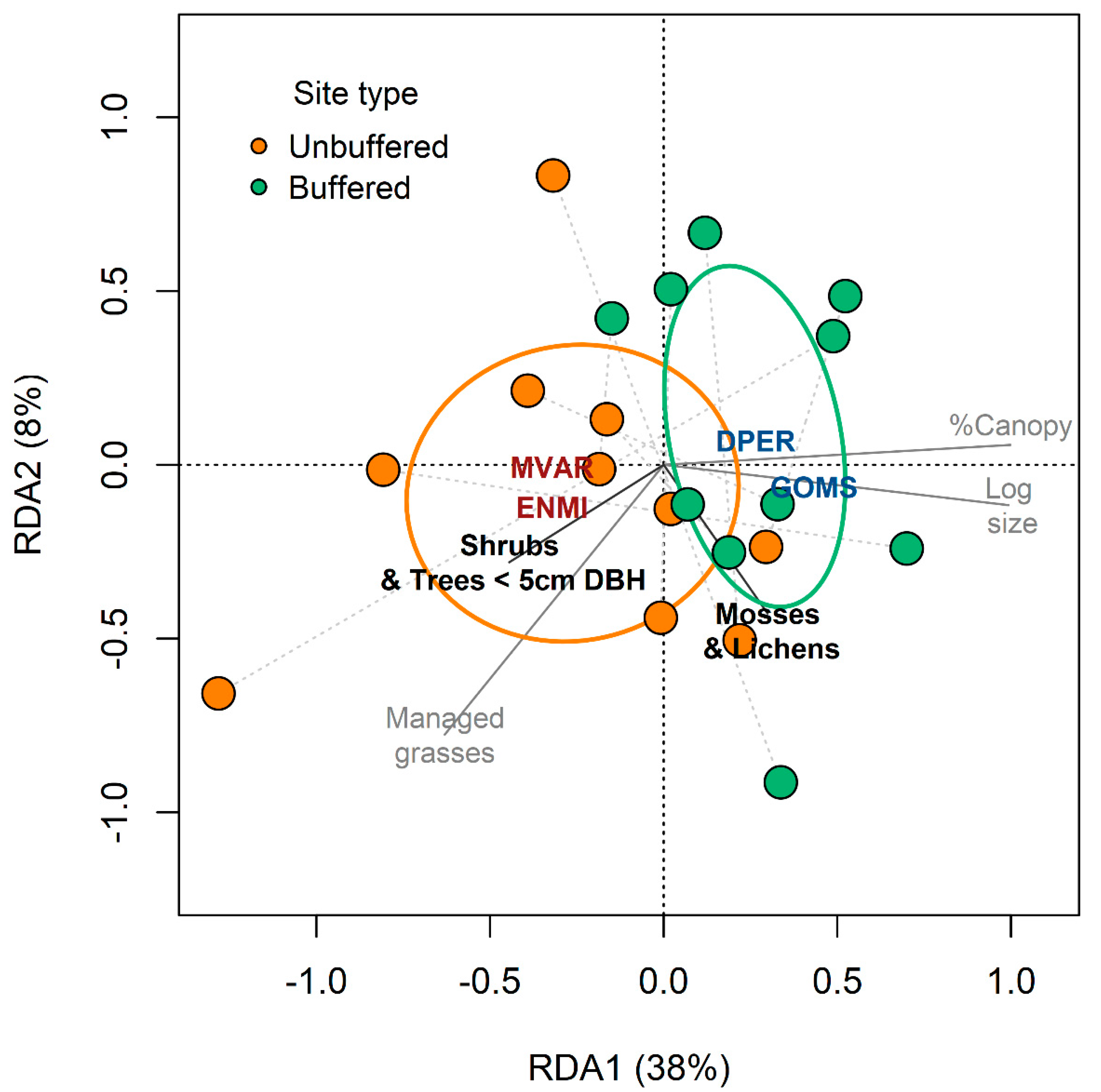
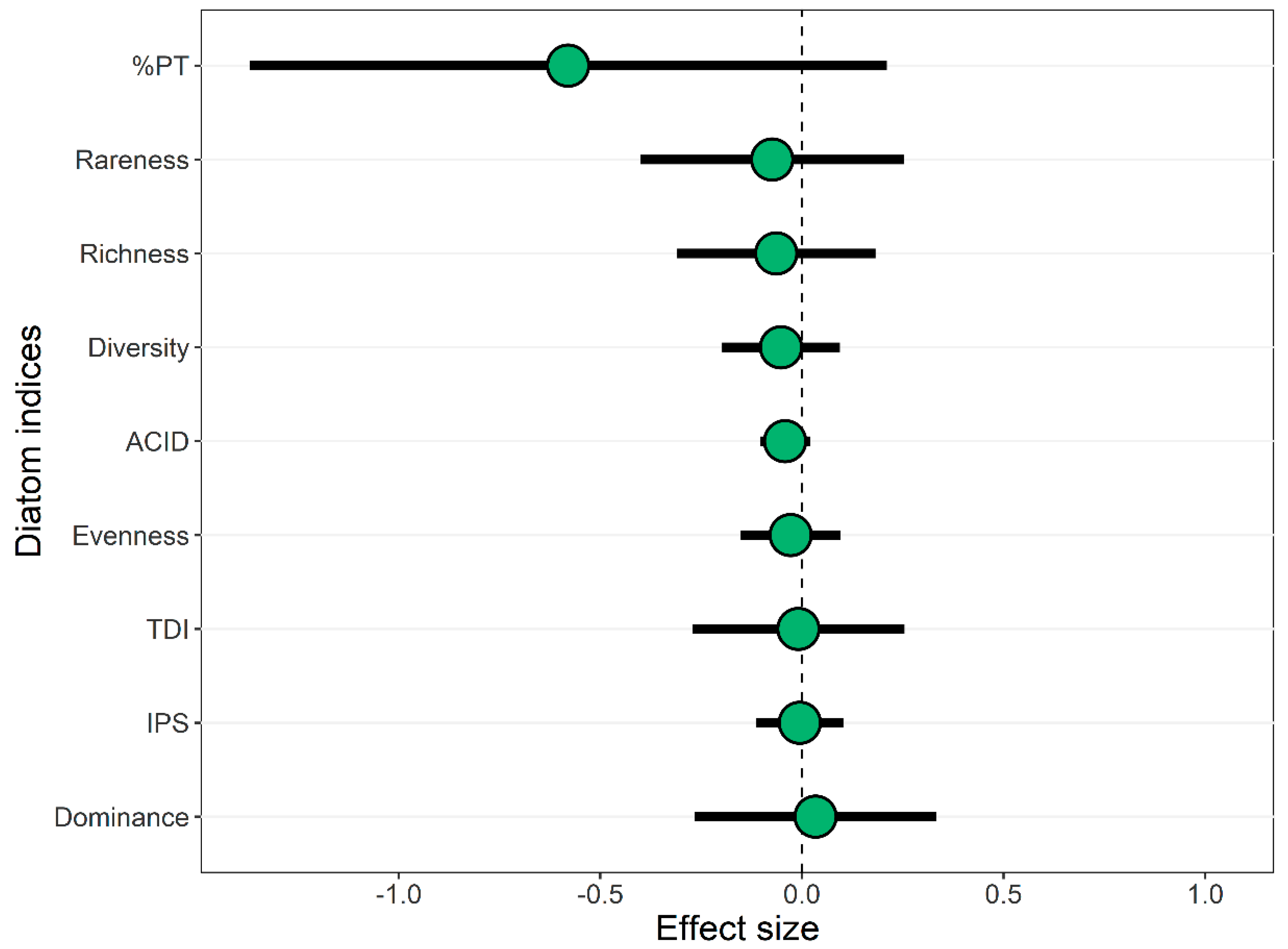
3.3. Forested Buffers and Diatoms in Urban Streams
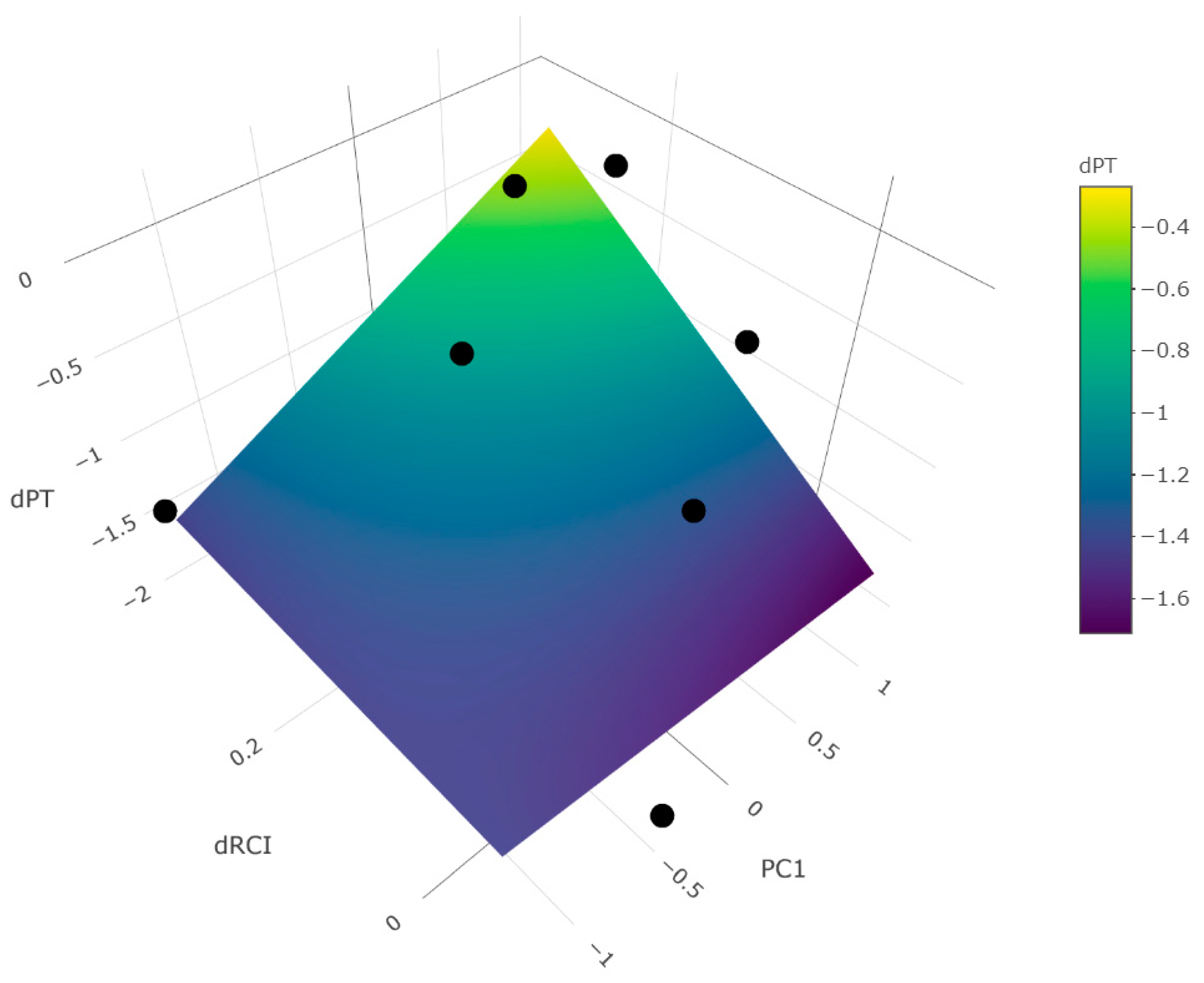
3.4. Context Dependent Influences of Forested Buffers on Diatoms
4. Discussion
4.1. Diatom Responses to Urbanization
4.2. Forest Buffers and Diatoms in Urban Streams
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A–Additional Site Information
Appendix B–Additional Results Complementing the Main Text
References
- Vörösmarty, C.J.; McIntyre, P.B.; Gessner, M.O.; Dudgeon, D.; Prusevich, A.; Green, P.; Glidden, S.; Bunn, S.E.; Sullivan, C.; Liermann, C.R.; et al. Global threats to human water security and river biodiversity. Nature 2010, 467, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Grimm, N.; Faeth, S.H.; Golubiewski, N.E.; Redman, C.L.; Wu, J.; Bai, X.; Briggs, J.M. Global Change and the Ecology of Cities. Science 2008, 319, 756–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, C.J.; Roy, A.H.; Feminella, J.W.; Cottingham, P.D.; Groffman, P.M.; Morgan, R.P. The urban stream syndrome: Current knowledge and the search for a cure. J. North Am. Benthol. Soc. 2005, 24, 706–723. [Google Scholar] [CrossRef]
- Wiederkehr, F.; Wilkinson, C.L.; Zeng, Y.; Yeo, D.C.; Ewers, R.M.; O’Gorman, E.J. Urbanisation affects ecosystem functioning more than structure in tropical streams. Biol. Conserv. 2020, 249, 108634. [Google Scholar] [CrossRef]
- King, R.S.; Baker, M.E.; Kazyak, P.F.; E Weller, D. How novel is too novel? Stream community thresholds at exceptionally low levels of catchment urbanization. Ecol. Appl. 2011, 21, 1659–1678. [Google Scholar] [CrossRef]
- Burdon, F.J.; Bai, Y.; Reyes, M.; Tamminen, M.; Staudacher, P.; Mangold, S.; Singer, H.; Räsänen, K.; Joss, A.; Tiegs, S.D.; et al. Stream microbial communities and ecosystem functioning show complex responses to multiple stressors in wastewater. Glob. Chang. Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Allan, J.D. Landscapes and Riverscapes: The Influence of Land Use on Stream Ecosystems. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 257–284. [Google Scholar] [CrossRef] [Green Version]
- Taylor, S.L.; Roberts, S.C.; Walsh, C.J.; Hatt, B.E. Catchment urbanisation and increased benthic algal biomass in streams: Linking mechanisms to management. Freshw. Biol. 2004, 49, 835–851. [Google Scholar] [CrossRef]
- Burdon, F.; Munz, N.; Reyes, M.; Focks, A.; Joss, A.; Räsänen, K.; Altermatt, F.; Eggen, R.; Stamm, C. Agriculture versus wastewater pollution as drivers of macroinvertebrate community structure in streams. Sci. Total. Environ. 2018, 659, 1256–1265. [Google Scholar] [CrossRef]
- Tonkin, J.D.; Stoll, S.; Jähnig, S.C.; Haase, P. Anthropogenic land-use stress alters community concordance at the river-riparian interface. Ecol. Indic. 2016, 65, 133–141. [Google Scholar] [CrossRef]
- Walker, C.E.; Pan, Y. Using Diatom Assemblages to Assess Urban Stream Conditions. Hydrobiology 2006, 561, 179–189. [Google Scholar] [CrossRef]
- Sudduth, E.; Hassett, B.A.; Cada, P.; Bernhardt, E.S. Testing the field of dreams hypothesis: Functional responses to urbanization and restoration in stream ecosystems. Ecol. Appl. 2011, 21, 1972–1988. [Google Scholar] [CrossRef] [PubMed]
- Coelho, D.; Hughes, S.J.; Varandas, S.; Cortes, R.M.V. Conservation benefits of riparian buffers in urban areas: The case of the Rio Corgo (north Portugal). Fundam. Appl. Limnol. Archiv Hydrobiol. 2014, 185, 55–70. [Google Scholar] [CrossRef]
- Burdon, F.J.; Ramberg, E.; Sargac, J.; Forio, M.A.E.; De Saeyer, N.; Mutinová, P.T.; Moe, T.F.; Pavelescu, M.O.; Dinu, V.; Cazacu, C.; et al. Assessing the Benefits of Forested Riparian Zones: A Qualitative Index of Riparian Integrity Is Positively Associated with Ecological Status in European Streams. Water 2020, 12, 1178. [Google Scholar] [CrossRef] [Green Version]
- Wenger, S. A Review of the Scientific Literature on Riparian Buffer Width, Extent and Vegetation; Office of Public Service & Outreach Institute of Ecology: Athens, Greece, 1999. [Google Scholar]
- Sabater, S.; Butturini, A.; Clement, J.-C.; Burt, T.; Dowrick, D.; Hefting, M.M.; Matre, V.; Pinay, G.; Postolache, C.; Rzepecki, M.; et al. Nitrogen Removal by Riparian Buffers along a European Climatic Gradient: Patterns and Factors of Variation. Ecosystems 2003, 6, 0020–0030. [Google Scholar] [CrossRef]
- Burdon, F.J.; McIntosh, A.R.; Harding, J.S. Mechanisms of trophic niche compression: Evidence from landscape disturbance. J. Anim. Ecol. 2019, 89, 730–744. [Google Scholar] [CrossRef]
- Pasquino, V.; Saulino, L.; Pelosi, A.; Allevato, E.; Rita, A.; Todaro, L.; Saracino, A.; Chirico, G. Hydrodynamic behaviour of European black poplar (Populus nigra L.) under coppice management along Mediterranean river ecosystems. River Res. Appl. 2018, 34, 586–594. [Google Scholar] [CrossRef]
- De Andrade, A.C.; Medeiros, W.; Adams, M. Urban forest fragments as unexpected sanctuaries for the rare endemic ghost butterfly from the Atlantic forest. Ecol. Evol. 2019, 9, 10767–10776. [Google Scholar] [CrossRef]
- Ives, C.D.; Lentini, P.E.; Threlfall, C.G.; Ikin, K.; Shanahan, D.F.; Garrard, G.E.; Bekessy, S.A.; Fuller, R.A.; Mumaw, L.M.; Rayner, L.; et al. Cities are hotspots for threatened species. Glob. Ecol. Biogeogr. 2015, 25, 117–126. [Google Scholar] [CrossRef]
- Kleerekoper, L.; Esch, M.P.-V.; Salcedo, T.B. How to make a city climate-proof, addressing the urban heat island effect. Resour. Conserv. Recycl. 2012, 64, 30–38. [Google Scholar] [CrossRef]
- Carter, J.; Cavan, G.; Connelly, A.; Guy, S.C.; Handley, J.; Kaźmierczak, A. Climate change and the city: Building capacity for urban adaptation. Prog. Plan. 2015, 95, 1–66. [Google Scholar] [CrossRef]
- Kenwick, R.A.; Shammin, R.; Sullivan, W.C. Preferences for riparian buffers. Landsc. Urban Plan. 2009, 91, 88–96. [Google Scholar] [CrossRef]
- Bernhardt, E.S.; Palmer, M.A. Restoring streams in an urbanizing world. Freshw. Biol. 2007, 52, 738–751. [Google Scholar] [CrossRef]
- Walsh, C.J.; Waller, K.A.; Gehling, J.; Mac Nally, R. Riverine invertebrate assemblages are degraded more by catchment urbanisation than by riparian deforestation. Freshw. Biol. 2007, 52, 574–587. [Google Scholar] [CrossRef]
- Bernhardt, E.S.; Palmer, M.A. River restoration: The fuzzy logic of repairing reaches to reverse catchment scale degradation. Ecol. Appl. 2011, 21, 1926–1931. [Google Scholar] [CrossRef] [PubMed]
- Aberle, J.; Järvelä, J. Flow resistance of emergent rigid and flexible floodplain vegetation. J. Hydraul. Res. 2013, 51, 33–45. [Google Scholar] [CrossRef]
- Lowe, R.; Stoermer, E.F.; Smol, J.P. The Diatoms: Applications for the Environmental and Earth Sciences. J. North Am. Benthol. Soc. 1999, 18, 567. [Google Scholar] [CrossRef]
- Kelly, M.; King, L.; Chatháin, B.N. The conceptual basis of ecological-status assessments using diatoms. Biol. Environ. Proc. R. Ir. Acad. 2009, 109, 175–189. [Google Scholar] [CrossRef]
- Kahlert, M.; Kelly, M.; Albert, R.-L.; Almeida, S.; Bešta, T.; Blanco, S.; Coste, M.; Denys, L.; Ector, L.; Fránková, M.; et al. Identification versus counting protocols as sources of uncertainty in diatom-based ecological status assessments. Hydrobiology 2012, 695, 109–124. [Google Scholar] [CrossRef]
- Kelly, M.G.; Whitton, B.A. The Trophic Diatom Index: A new index for monitoring eutrophication in rivers. Environ. Biol. Fishes 1995, 7, 433–444. [Google Scholar] [CrossRef]
- Andrén, C.; Jarlman, A. Benthic diatoms as indicators of acidity in streams. Fundam. Appl. Limnol. Archiv Hydrobiol. 2008, 173, 237–253. [Google Scholar] [CrossRef]
- Sonneman, J.A.; Walsh, C.J.; Breen, P.F.; Sharpe, A.K. Effects of urbanization on streams of the Melbourne region, Victoria, Australia. II. Benthic diatom communities. Freshw. Biol. 2001, 46, 553–565. [Google Scholar] [CrossRef]
- CEMAGREF. Etude des Méthodes Biologiques D’appréciation Quantitative de la Qualité des Eaux; Bassin Rhône-Mediterrannée-Corse, E.R.Q., Lyon, A.F.G.S., Eds.; EMAGREF/CSIRO: Lyon, France, 1982. [Google Scholar]
- Coste, M.; Boutry, S.; Tison, J.; Delmas, F. Improvements of the Biological Diatom Index (BDI): Description and efficiency of the new version (BDI-2006). Ecol. Indic. 2009, 9, 621–650. [Google Scholar] [CrossRef]
- Teittinen, A.; Taka, M.; Ruth, O.; Soininen, J. Variation in stream diatom communities in relation to water quality and catchment variables in a boreal, urbanized region. Sci. Total. Environ. 2015, 530, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Newall, P.; Walsh, C.J. Response of epilithic diatom assemblages to urbanization influences. Hydrobiology 2005, 532, 53–67. [Google Scholar] [CrossRef]
- Hill, B.H.; Stevenson, R.J.; Pan, Y.; Herlihy, A.T.; Kaufmann, P.R.; Johnson, C.B. Comparison of correlations between environmental characteristics and stream diatom assemblages characterized at genus and species levels. J. North Am. Benthol. Soc. 2001, 20, 299–310. [Google Scholar] [CrossRef] [Green Version]
- Johnson, R.K.; Almlöf, K. Adapting boreal streams to climate change: Effects of riparian vegetation on water temperature and biological assemblages. Freshw. Sci. 2016, 35, 984–997. [Google Scholar] [CrossRef]
- Kelly, M. Use of the Trophic Diatom Index to monitor eutrophication in rivers. Water Res. 1998, 32, 236–242. [Google Scholar] [CrossRef]
- Chen, X.; Zhou, W.; Pickett, S.T.; Li, W.; Han, L.; Ren, Y. Diatoms are better indicators of urban stream conditions: A case study in Beijing, China. Ecol. Indic. 2016, 60, 265–274. [Google Scholar] [CrossRef]
- Hlúbiková, D.; Novais, M.H.; Dohet, A.; Hoffmann, L.; Ector, L. Effect of riparian vegetation on diatom assemblages in headwater streams under different land uses. Sci. Total. Environ. 2014, 475, 234–247. [Google Scholar] [CrossRef]
- Allan, I.; Jenssen, M.T.S.; Braaten, H.F.V. Priority Substances and Emerging Contaminants in Selected Norwegian Rivers—The River Monitoring Programme 2018; Norwegian Institute for Water Research: Oslo, Norway, 2019. [Google Scholar]
- European Commission. Oslo—European Green Capital 2019; Publications Office of the European Union: Luxembourg, 2018; Available online: https://ec.europa.eu/environment/europeangreencapital/wp-content/uploads/2019/Oslo%20Brochure_EGCA%202019.pdf (accessed on 9 October 2020).
- Kerouel, R.; Aminot, A. Fluorometric determination of ammonia in sea and estuarine waters by direct segmented flow analysis. Mar. Chem. 1997, 57, 265–275. [Google Scholar] [CrossRef]
- Wentworth, C.K. A scale of grade and class terms for clastic sediments. J. Geol. 1922, 30, 377–392. [Google Scholar] [CrossRef]
- Wolman, M.G. A method of sampling coarse river-bed material. Trans. Am. Geophys. Union 1954, 35, 951–956. [Google Scholar] [CrossRef]
- Harding, J.S.; Clapcott, J.; Quinn, J.; Hayes, J.; Joy, M.; Storey, R.; Greig, H.; Hay, J.; James, T.; Beech, M.; et al. Stream Habitat Assessment Protocols for Wadeable Rivers and Streams of New Zealand; School of Biological Sciences: Christchurch, New Zealand, 2009. [Google Scholar]
- CEN, NS-EN 13946:2014 Water Quality—Guidance for the Routine Sampling and Preparation of Benthic Diatoms from Rivers and Lakes. 2014. Available online: https://www.standard.no/en/webshop/ProductCatalog/ProductPresentation/?ProductID=698749 (accessed on 9 October 2020).
- CEN, NS-EN 14407:2014 Water Quality—Guidance for the Identification and Enumeration of Benthic Diatom Samples from Rivers and Lakes. 2014. Available online: https://www.standard.no/en/webshop/ProductCatalog/ProductPresentation/?ProductID=698752 (accessed on 9 October 2020).
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae 1. Teil: Naviculaceae. In Süßwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Stuttgart, Germany, 1986. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae 2. Teil: Bacillariaceae, Epithemiaceae, Surirellaceae. In Süßwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Stuttgart, Germany, 1988. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae 3. Teil: Centrales, Fragilariaceae, Eunotiaceae. In Süßwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Stuttgart, Germany, 1991. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae 4. Teil: Achnanthaceae, Kritische Ergänzungen zu Navicula (Lineolatae) und Gomphonema. In Süßwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Stuttgart, Germany, 1991. [Google Scholar]
- Krammer, K. Diatoms of Europe, Volume 1: The genus Pinnularia. In Diatoms of the European Inland Waters and Comparable Habitats; Lange-Bertalot, H., Ed.; Gantner Verlag: Ruggell, Liechtenstein, 2000. [Google Scholar]
- Krammer, K. Diatoms of Europe, Volume 4: Cymbopleura, Delicata, Navicymbula, Gomphocymbellopsis, Afrocymbella. In Diatoms of the European Inland Waters and Comparable Habitats; Lange-Bertalot, H., Ed.; Gantner Verlag: Ruggell, Liechtenstein, 2003. [Google Scholar]
- Krammer, K. Diatoms of Europe, Volume 1: The genus Cymbella. In Diatoms of the European Inland Waters and Comparable Habitats; Lange-Bertalot, H., Ed.; Gantner Verlag: Ruggell, Liechtenstein, 2002. [Google Scholar]
- Lange-Bertalot, H.; Metzeltin, D. Oligotrophie-Indikatoren: 800 Taxa repräsentativ für drei diverse Seen-Typen, kalkreich-oligodystroph-schwach gepuffertes Weichwasser. In Iconogr. Diatomol; Lange-Bertalot, H., Ed.; Koeltz Scientific Books, Schmitten: Oberreifenberg, Germany, 1996. [Google Scholar]
- Lange-Bertalot, H. Diatoms of Europe, Volume 2: Navicula sensu stricto, 10 genera separated from Navicula sensu lato, Frustulia. In Diatoms of the European Inland Waters and Comparable Habitats; Lange-Bertalot, H., Ed.; Ganthner Verlag: Ruggell, Liechtenstein, 2011. [Google Scholar]
- Lange-Bertalot, H.; Bak, M.; Witkowski, A.; Tagliaventi, N. Diatoms of Europe, Volume 6: Eunotia and some related genera. In Diatoms of the European Inland Water and Comparable Habitats; Krammer, K., Ed.; Ganthner Verlag: Ruggell, Liechtenstein, 2011. [Google Scholar]
- Lange-Bertalot, H.; Hofmann, G.; Werum, M.; Cantonati, M. Freshwater Benthic Diatoms of Central Europe: Over 800 Common Species Used in Ecological Assessment. In Koeltz Botanical Books; Cantonati, M., Kelly, M.G., Lange–Bertalot, H., Eds.; Schmitten: Oberreifenberg, Germany, 2017. [Google Scholar]
- Lecointe, C.; Coste, M.; Prygiel, J. “Omnidia”: Software for taxonomy, calculation of diatom indices and inventories management. Twelfth Int. Diatom Symp. 1993, 269, 509–513. [Google Scholar] [CrossRef]
- Burdon, F.J.; Reyes, M.; Alder, A.C.; Joss, A.; Ort, C.; Räsänen, K.; Jokela, J.; Eggen, R.I.; Stamm, C. Environmental context and magnitude of disturbance influence trait-mediated community responses to wastewater in streams. Ecol. Evol. 2016, 6, 3923–3939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burdon, F.J.; McIntosh, A.; Harding, J.S. Habitat loss drives threshold response of benthic invertebrate communities to deposited sediment in agricultural streams. Ecol. Appl. 2013, 23, 1036–1047. [Google Scholar] [CrossRef]
- Bray, J.; Kilroy, C.; Gerbeaux, P.; Burdon, F.J.; Harding, J.S. Ecological processes mediate the effects of the invasive bloom-forming diatom Didymosphenia geminata on stream algal and invertebrate assemblages. Hydrobiology 2019, 847, 1–14. [Google Scholar] [CrossRef]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package; R Foundation for Statistical Computing: Vienna, Austria, 2009. [Google Scholar]
- Blanchet, F.G.; Legendre, P.; Borcard, D. Forward selection of explanatory variables. Ecology 2008, 89, 2623–2632. [Google Scholar] [CrossRef]
- Dray, S.; Legendre, P.; Blanchet, G. Packfor: Forward Selection with Permutation (Canoco p.46). R Package Version 0.0-8/r136. Available online: https://R-Forge.R-project.org/projects/sedar (accessed on 9 October 2020).
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press (CUP): Cambridge, UK, 2002; p. 570. [Google Scholar]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2011; p. 306. [Google Scholar]
- Borcard, D.; Legendre, P. All-scale spatial analysis of ecological data by means of principal coordinates of neighbour matrices. Ecol. Model. 2002, 153, 51–68. [Google Scholar] [CrossRef]
- Borcard, D.; Legendre, P.; Avois-Jacquet, C.; Tuomisto, H. Dissecting the spatial structure of ecological data at multiple scales. Ecology 2004, 85, 1826–1832. [Google Scholar] [CrossRef] [Green Version]
- Dray, S.; Legendre, P.; Peres-Neto, P.R. Spatial modelling: A comprehensive framework for principal coordinate analysis of neighbour matrices (PCNM). Ecol. Model. 2006, 196, 483–493. [Google Scholar] [CrossRef]
- Peres-Neto, P.R.; Legendre, P.; Dray, S.; Borcard, D. Variation partitioning of species data matrices: Estimation and comparison of fractions. Ecology 2006, 87, 261425. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.N. Generalized Additive Models; Informa UK Limited: London, UK, 2017; p. 496. [Google Scholar]
- Bates, D.M.; Machler, M.B.; Bolker, B.M.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Pustejovsky, J.E.; Swan, D.M. SingleCaseES: A Calculator for Single-Case Effect Sizes. R Package Version 0.4.3. Available online: https://CRAN.R-project.org/package=SingleCaseES. (accessed on 9 October 2020).
- Sundberg, I.; Herlitz, E.; Jarlman, A.; Kahlert, M. Påväxt i sjöar och vattendrag—kiselalgsanalys. In Havs- och Vattenmyndigheten; Handledning för miljöövervakning: Göteborg, Sweden, 2016; p. 24, Version 3:2: 2016-01-20. [Google Scholar]
- Pan, Y.; Herlihy, A.T.; Kaufmann, P.; Wigington, J.; Van Sickle, J.; Moser, T. Linkages among land-use, water quality, physical habitat conditions and lotic diatom assemblages: A multi-spatial scale assessment. Hydrobiology 2004, 515, 59–73. [Google Scholar] [CrossRef]
- Dodds, W.K.; Clements, W.H.; Gido, K.; Hilderbrand, R.H.; King, R.S. Thresholds, breakpoints, and nonlinearity in freshwaters as related to management. J. North Am. Benthol. Soc. 2010, 29, 988–997. [Google Scholar] [CrossRef]
- Capon, S.J.; Lynch, A.J.J.; Bond, N.R.; Chessman, B.C.; Davis, J.; Davidson, N.; Finlayson, C.M.; Gell, P.; Hohnberg, D.; Humphrey, C.; et al. Regime shifts, thresholds and multiple stable states in freshwater ecosystems; a critical appraisal of the evidence. Sci. Total. Environ. 2015, 534, 122–130. [Google Scholar] [CrossRef] [Green Version]
- Taylor, J.M.; King, R.S.; Pease, A.A.; Winemiller, K.O. Nonlinear response of stream ecosystem structure to low-level phosphorus enrichment. Freshw. Biol. 2014, 59, 969–984. [Google Scholar] [CrossRef]
- Tang, T.; Tang, T.; Tan, L.; Gu, Y.; Jiang, W.; Cai, Q.-H. Identifying community thresholds for lotic benthic diatoms in response to human disturbance. Sci. Rep. 2017, 7, 4134. [Google Scholar] [CrossRef] [Green Version]
- Tibby, J.; Richards, J.; Tyler, J.J.; Barr, C.; Fluin, J.; Goonan, P. Diatom–water quality thresholds in South Australian streams indicate a need for more stringent water quality guidelines. Mar. Freshw. Res. 2019, 71, 942–952. [Google Scholar] [CrossRef]
- Finkel, Z.V.; Beardall, J.; Flynn, K.J.; Quigg, A.; Rees, T.A.V.; Raven, J.A. Phytoplankton in a changing world: Cell size and elemental stoichiometry. J. Plankton Res. 2009, 32, 119–137. [Google Scholar] [CrossRef] [Green Version]
- Walsh, C.J.; Sharpe, A.K.; Breen, P.F.; Sonneman, J.A. Effects of urbanization on streams of the Melbourne region, Victoria, Australia. I. Benthic macroinvertebrate communities. Freshw. Biol. 2001, 46, 535551. [Google Scholar] [CrossRef]
- Heino, J. The importance of metacommunity ecology for environmental assessment research in the freshwater realm. Biol. Rev. 2012, 88, 166–178. [Google Scholar] [CrossRef] [PubMed]
- McBride, M.; Booth, D.B. Urban Impacts On Physical Stream Condition: Effects Of Spatial Scale, Connectivity, And Longitudinal Trends. Jawra J. Am. Water Resour. Assoc. 2005, 41, 565–580. [Google Scholar] [CrossRef]
- Kaushal, S.S.; Belt, K.T. The urban watershed continuum: Evolving spatial and temporal dimensions. Urban Ecosyst. 2012, 15, 409–435. [Google Scholar] [CrossRef]
- Wenger, S.J.; Roy, A.H.; Jackson, C.R.; Bernhardt, E.S.; Carter, T.L.; Filoso, S.; Gibson, C.A.; Hession, W.C.; Kaushal, S.S.; Martí, E.; et al. Twenty-six key research questions in urban stream ecology: An assessment of the state of the science. J. North Am. Benthol. Soc. 2009, 28, 1080–1098. [Google Scholar] [CrossRef] [Green Version]
- Riis, T.; Kelly-Quinn, M.; Aguiar, F.C.; Manolaki, P.; Bruno, D.; Bejarano, M.D.; Clerici, N.; Fernandes, M.R.; Franco, J.C.; Pettit, N.; et al. Global overview of ecosystem services provided by riparian vegetation. BioScience 2020, 70, 501–514. [Google Scholar] [CrossRef]
- Lovett, G.M.; Likens, G.E.; Buso, D.C.; Driscoll, C.T.; Bailey, S. The biogeochemistry of chlorine at Hubbard Brook, New Hampshire, USA. Biogeochemistry 2005, 72, 191–232. [Google Scholar] [CrossRef]
- Kaushal, S.S.; Groffman, P.M.; Likens, G.E.; Belt, K.T.; Stack, W.P.; Kelly, V.R.; Band, L.E.; Fisher, G.T. Increased salinization of fresh water in the northeastern United States. Proc. Natl. Acad. Sci. USA 2005, 102, 13517–13520. [Google Scholar] [CrossRef] [Green Version]
- Porter-Goff, E.R.; Frost, P.C.; Xenopoulos, M.A. Changes in riverine benthic diatom community structure along a chloride gradient. Ecol. Indic. 2013, 32, 97–106. [Google Scholar] [CrossRef]
- Mitani, E.; Fumiyo, N.; Izumi, M.; Ichi, I.; Kawabata, A.; Kawachi, M.; Kato, M. Fatty acid composition profiles of 235 strains of three microalgal divisions within the NIES Microbial Culture Collection. Microb. Resour. Syst. 2017, 33, 1929. [Google Scholar]
- Twining, C.W.; Brenna, J.T.; Hairston, N.G.; Flecker, A.S. Highly unsaturated fatty acids in nature: What we know and what we need to learn. Oikos 2015, 125, 749–760. [Google Scholar] [CrossRef]
- Chari, L.; Richoux, N.; Moyo, S.; Villet, M. Dietary fatty acids of spiders reveal spatial and temporal variations in aquatic-terrestrial linkages. Food Webs 2020, 24, e00152. [Google Scholar] [CrossRef]
- Keck, F.; Vasselon, V.; Tapolczai, K.; Rimet, F.; Bouchez, A. Freshwater biomonitoring in the Information Age. Front. Ecol. Environ. 2017, 15, 266–274. [Google Scholar] [CrossRef]
- Zimmermann, J.; Glöckner, G.; Jahn, R.; Enke, N.; Gemeinholzer, B. Metabarcoding vs. morphological identification to assess diatom diversity in environmental studies. Mol. Ecol. Resour. 2014, 15, 526–542. [Google Scholar] [CrossRef] [PubMed]
- Hering, D.; Borja, A.; Jones, J.; Pont, D.; Boets, P.; Bouchez, A.; Bruce, K.; Drakare, S.; Hänfling, B.; Kahlert, M.; et al. Implementation options for DNA-based identification into ecological status assessment under the European Water Framework Directive. Water Res. 2018, 138, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Potapova, M.; Hamilton, P.B. Morphological and ecological variation within the Achnanthidium minutissimum (Bacillariophyceae) species complex. J. Phycol. 2007, 43, 561–575. [Google Scholar] [CrossRef]
- Pinseel, E.; Vanormelingen, P.; Hamilton, P.B.; Vyverman, W.; Van De Vijver, B.; Kopalova, K. Molecular and morphological characterization of the Achnanthidium minutissimum complex (Bacillariophyta) in Petuniabukta (Spitsbergen, High Arctic) including the description of A. digitatum sp. nov. Eur. J. Phycol. 2017, 52, 264–280. [Google Scholar] [CrossRef]
- Romero, O.E.; Jahn, R. Typification of Cocconeis lineata and Cocconeis euglypta (Bacillariophyta). Diatom Res. 2013, 28, 175–184. [Google Scholar] [CrossRef]
- Berg, S.K.; Stenbro, R. Densification or Dilution? On Cultural and Economic Value Creation along the Aker River in Oslo, Norway. Hist. Environ. Policy Pract. 2015, 6, 197–213. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mutinova, P.T.; Kahlert, M.; Kupilas, B.; McKie, B.G.; Friberg, N.; Burdon, F.J. Benthic Diatom Communities in Urban Streams and the Role of Riparian Buffers. Water 2020, 12, 2799. https://doi.org/10.3390/w12102799
Mutinova PT, Kahlert M, Kupilas B, McKie BG, Friberg N, Burdon FJ. Benthic Diatom Communities in Urban Streams and the Role of Riparian Buffers. Water. 2020; 12(10):2799. https://doi.org/10.3390/w12102799
Chicago/Turabian StyleMutinova, Petra Thea, Maria Kahlert, Benjamin Kupilas, Brendan G. McKie, Nikolai Friberg, and Francis J. Burdon. 2020. "Benthic Diatom Communities in Urban Streams and the Role of Riparian Buffers" Water 12, no. 10: 2799. https://doi.org/10.3390/w12102799
APA StyleMutinova, P. T., Kahlert, M., Kupilas, B., McKie, B. G., Friberg, N., & Burdon, F. J. (2020). Benthic Diatom Communities in Urban Streams and the Role of Riparian Buffers. Water, 12(10), 2799. https://doi.org/10.3390/w12102799