The Effect of Monsoon Rainfall Patterns on Epilithic Diatom Communities in the Hantangang River, Korea



Abstract
:1. Introduction
2. Materials and Methods
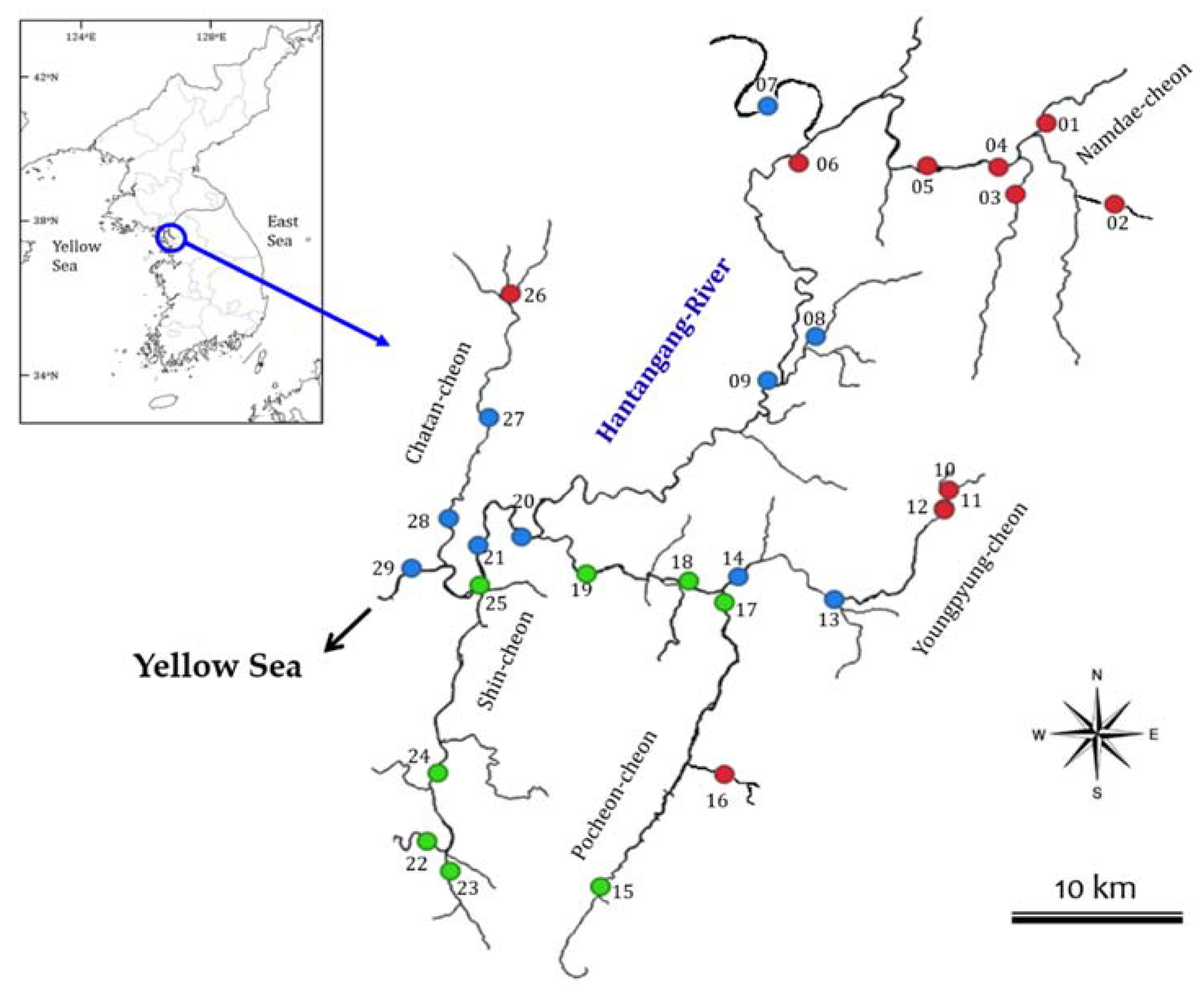
2.1. Study Area
2.2. Sampling of River Diatom and Waters
2.3. Analysis of Water Quality Parameters
2.4. Analysis of Epilithic Diatoms
2.5. Measurement of Diatom Community Changes
- d is the distance between two communities with different times at the specific site in PCA dimension.
- x1 and y1 is the first sampling site in PCA dimension before monsoon
- x2 and y2 is the second sampling site in PCA dimension after monsoon rainfall
- di is the distance between different times of ith community before and after monsoon rainfall
- n is the number of sampling sites or communities
2.6. Data Analysis
3. Results
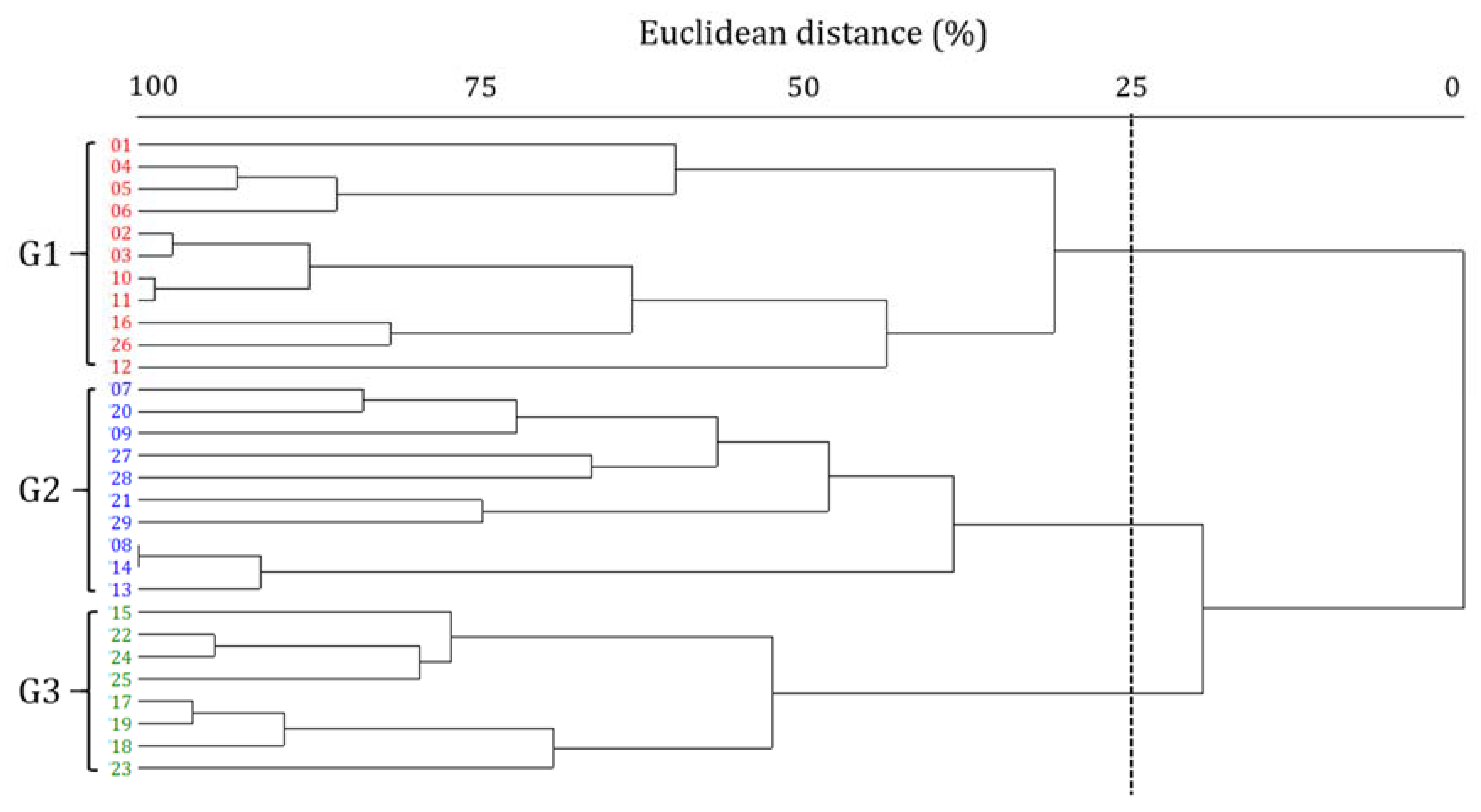
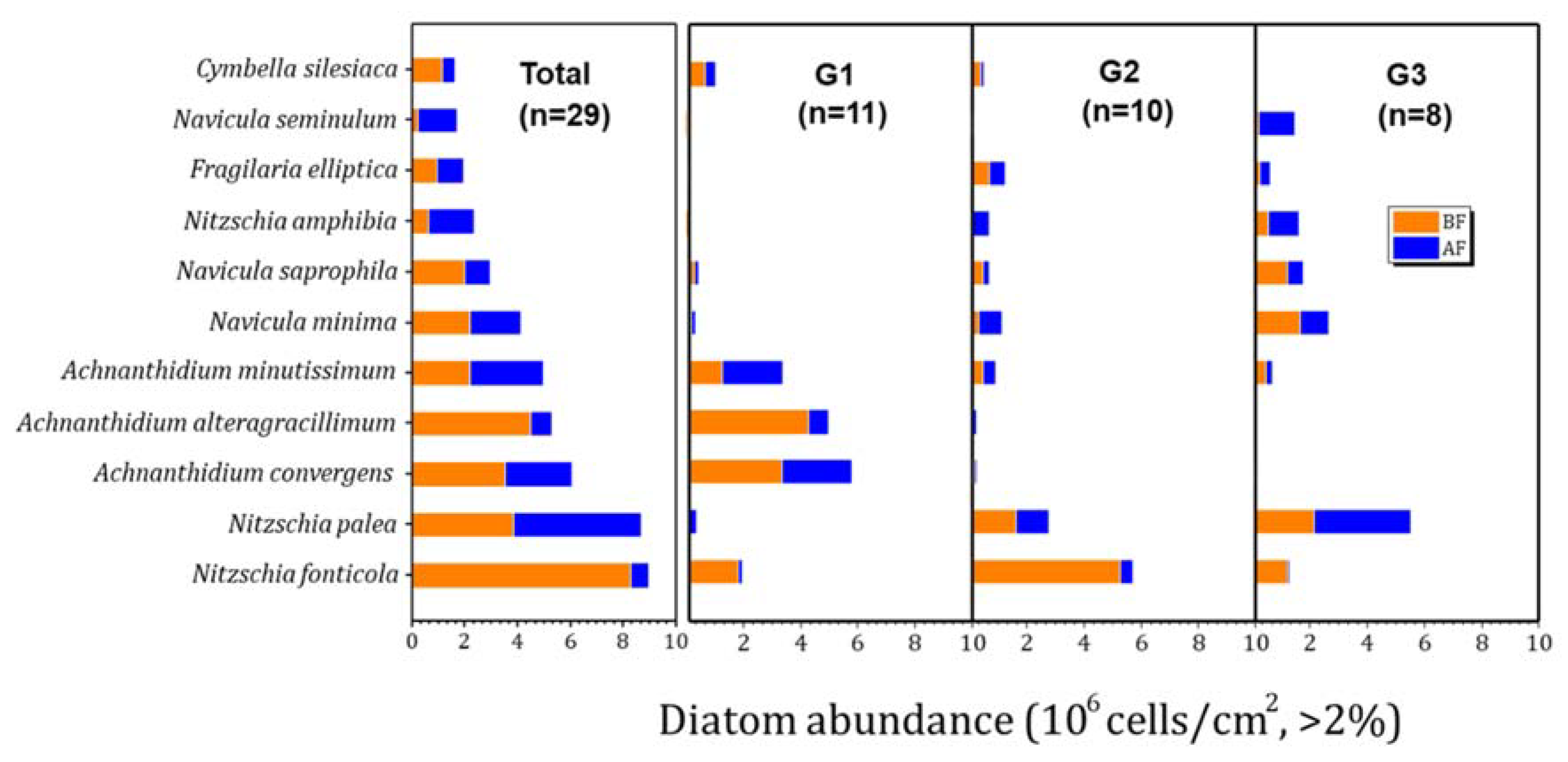
3.1. Epilithic Diatoms of the Hantangang River
3.2. Water Quality Parameters of the Hantangang River
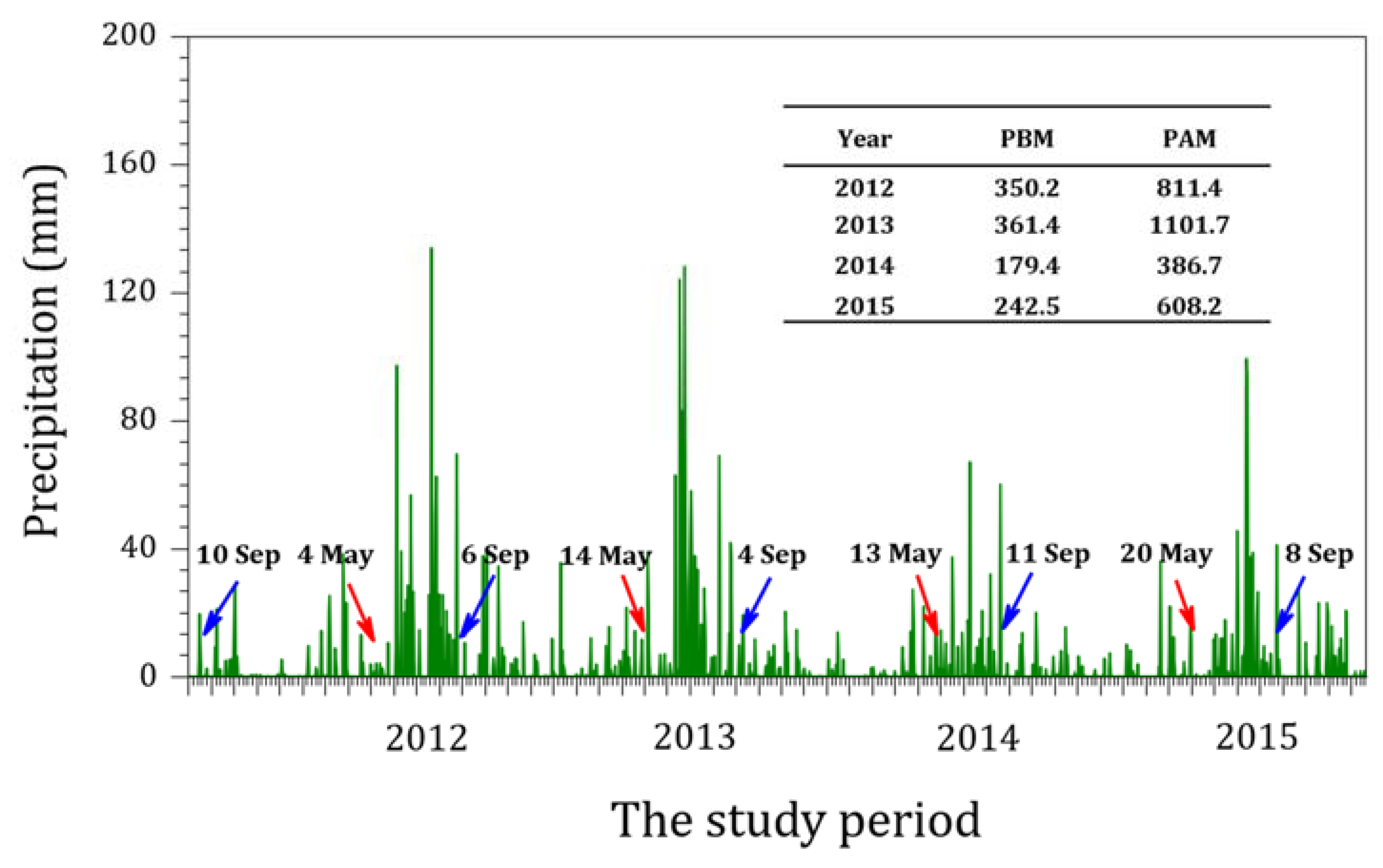
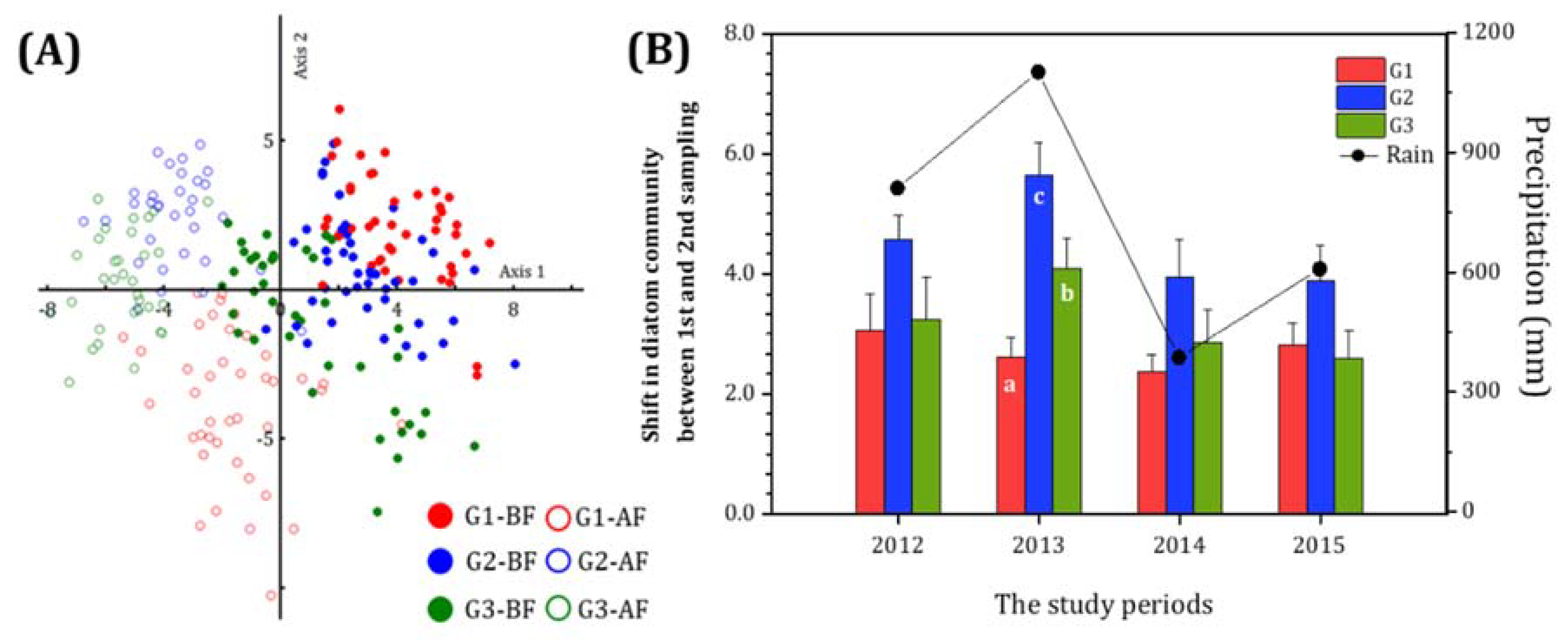
3.3. Shift in Diatom Communities before and after monsoon
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kim, C.H.; Jhun, J.G. Numerical simulations of the three-dimensional land and sea breezes under synoptic flows over South Korea. APJAS 1992, 28, 165–181. [Google Scholar]
- Yun, W.T.; Park, C.K.; Lee, J.W.; Lee, H.S.; Min, S.K. Analysis of the Korean Heavy Rainfall Features in Summer 1998. J. Korean Meteorol. Soc. 2001, 37, 181–194. [Google Scholar]
- Vincent, C.E.; Davies, T.D.; Beresford, A.K.C. Recent changes in the level of Lake Naivasha, Kenya, as an indicator of equatorial westerlies over East Africa. Clim. Chang. 1979, 2, 175–189. [Google Scholar] [CrossRef]
- Johnson, G.; Harper, D.M.; Mavuti, K. Information for the Sustainable Management of Shallow Lakes: Lake Naivasha, Kenya; The Sustainable Management of Tropical Catchments; Wiley: Chichester, UK, 1998. [Google Scholar]
- Harper, D.M. Primary production in Lake Naivasha, Kenya. Int. Ver. Theor. Angew. Limnol. 1991, 24, 1112–1116. [Google Scholar] [CrossRef]
- Mallin, M.A.; Johnson, V.L.; Ensign, S.H. Comparative impacts of stormwater runoff on water quality of an urban, a suburban, and a rural stream. Environ. Monit. Assess. 2009, 159, 475–491. [Google Scholar] [CrossRef] [PubMed]
- Jeong, K.S.; Kim, D.K.; Joo, G.J. Delayed influence of dam storage and discharge on the determination of seasonal proliferations of Microcystis aeruginosa and Stephanodiscus hantzschii in a regulated river system of the lower Nakdong River (South Korea). Water Res. 2007, 41, 1269–1279. [Google Scholar] [CrossRef]
- Liu, D.Y.; Sun, J.; Zou, J.Z.; Zhang, J. Phytoplankton succession during a red tide of Skeletonema costatum in Jiaozhou Bay of China. Mar. Pollut. Bull. 2005, 50, 91–94. [Google Scholar] [CrossRef]
- Jose, E.C.; Furio, E.F.; Borja, V.M.; Gatdula, N.C.; Santos, M.D. Zooplankton Composition and Abundance and its Relationship with Physico-Chemical Parameters in Manila Bay. Oceanography 2015, 3, 136. [Google Scholar] [CrossRef]
- Zhou, G.; Zhao, X.; Bi, Y.; Hu, Z. Effects of rainfall on spring phytoplankton community structure in Xiangxi Bay of the Three-Gorges Reservoir, China. Fresen. Environ. Bull. 2012, 21, 3533–3541. [Google Scholar]
- Lu, M.; Xie, P. Impacts of filter-feeding fishes on the long-term changes of crustacean zooplankton in a eutrophic subtropical Chinese lake. J. Freshw. Ecol. 2001, 16, 219–228. [Google Scholar] [CrossRef] [Green Version]
- Pinto-Coelho, R.; Pinel-Alloul, B.; Méthot, G.; Havens, K.E. Crustacean zooplankton in lakes and reservoirs of temperate and tropical regions: Variation with trophic status. Can. J. Fish. Aquat. Sci. 2005, 62, 348–361. [Google Scholar] [CrossRef] [Green Version]
- Da Silva Abes, S.; Agostinho, A.A. Spatial patterns in fish distributions and structure of the ichthyocenosis in the Água Nanci stream, upper Paraná River basin, Brazil. Hydrobiologia 2001, 445, 217–227. [Google Scholar] [CrossRef]
- Lacoul, P.; Freedman, B. Environmental influences on aquatic plants in freshwater ecosystems. Environ. Rev. 2006, 14, 89–136. [Google Scholar] [CrossRef]
- Staley, J.T.; Hodgson, C.J.; Mortimer, S.R.; Morecroft, M.D.; Masters, G.J.; Brown, V.K.; Taylor, M.E. Effects of summer rainfall manipulations on the abundance and vertical distribution of herbivorous soil macro-invertebrates. Eur. J. Soil Biol. 2007, 43, 189–198. [Google Scholar] [CrossRef]
- Beche, L.A.; Mcelravy, E.P.; Resh, V.H. Long-term seasonal variation in the biological traits of benthic-macroinvertebrates in two Mediterranean-climate streams in California, USA. Freshw. Biol. 2006, 51, 56–75. [Google Scholar] [CrossRef]
- Leland, H.V.; Porter, S.D. Distribution of benthic algae in the upper Illinois River basin in relation to geology and land use. Freshw. Biol. 2000, 44, 279–301. [Google Scholar] [CrossRef]
- Cushing, C.E.; Allan, J.D. Streams: Their Ecology and Life; Academic Press: San Diego, CA, USA, 2001. [Google Scholar]
- Khatoon, H.; Yusoff, F.; Banerjee, S.; Shariff, M.; Bujang, J.S. Formation of periphyton biofilm and subsequent biofouling on different substrates in nutrient enriched brackish water shrimp ponds. Aquaculture 2007, 273, 470–477. [Google Scholar] [CrossRef]
- Hill, W.R. Factors Affecting Benthic Algae—Effects of Light. Algal Ecology: Freshwater Benthic Ecosystem; Stevenson, R.J., Bothwell, M.L., Lowe, R.L., Eds.; Academic Press: San Diego, CA, USA, 1996. [Google Scholar]
- Kim, H.K.; Kim, Y.J.; Won, D.H.; Hwang, S.J.; Hwang, S.O.; Kim, B.H. Spatial and Temporal Distribution of Epilithic Diatom Communities in Major Harbors of Korean peninsula. J. Korean Soc. Wat. Environ. 2013, 29, 598–609. [Google Scholar]
- Cho, I.H.; Kim, H.K.; Choi, M.Y.; Kwon, Y.S.; Hwang, S.J.; Kim, S.H.; Kim, B.H. Distribution and Species Prediction of Epilithic Diatom in the Geum River Basin. Korean J. Limnol. 2015, 48, 153–167. [Google Scholar]
- Kim, Y.J. Temporal and Spatial Dynamics of an Epilithic Algal Community in the Hantan River. Algae 2004, 19, 15–22. [Google Scholar]
- Lau, K.M.; Li, M.T. The monsoon of East Asia and its global associations—A survey. Am. Meteorol. Soc. 1984, 65, 114–125. [Google Scholar] [CrossRef]
- Chung, H.S.; Chung, Y.A.; Kim, C.M.; Ryu, C.S. The Characteristics of Heavy Rainfall over the Korean Peninsula Case Studies of Heavy Rainfall Events during the On-and Off-Changma Season. J. Environ. Sci. 2012, 21, 1511–1521. [Google Scholar]
- Ha, K.J.; Yun, K.S.; Jhun, J.G.; Park, C.K. Definition of onset/retreat and intensity of changma during the boreal Summer monsoon season. J. Korean Meteorol. Soc. 2005, 41, 927–942. [Google Scholar]
- Byun, J.H.; Cho, I.H.; Hwang, S.J.; Park, M.H.; Byeon, M.S.; Kim, B.H. Relationship between a Dense Bloom of Cyanobacterium Anabaena spp. and Rainfalls in the North Han River System of South Korea. Korean J. Environ. Ecol. 2014, 47, 116–126. [Google Scholar] [CrossRef]
- APHA. Standard Methods for the Examination of Water and Waste Water; American Public Health Association: New York, NY, USA, 2001. [Google Scholar]
- MOE (Ministry of Environment, South Korea). The Survey and Evaluation of Aquatic Ecosystem Health in Korea; National Institute of Environmental Research (NIER): Incheon, Korea, 2015.
- Hendey, N.I. The permanganate method for cleaning freshly gathered diatoms. Microscopy 1974, 32, 423–426. [Google Scholar]
- McNaughton, S.J. Relationships among functional properties of Californian grassland. Nature 1967, 216, 168–169. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 318, 379–423. [Google Scholar]
- Margalef, R. Information theory in ecology, General Systems. Int. J. Gen. Syst. 1958, 32, 36–71. [Google Scholar]
- Pielou, E.C. Ecological Diversity; Wiley: New York, NY, USA, 1975. [Google Scholar]
- Hallett, L.M.; Hsu, J.S.; Cleland, E.E.; Collins, S.L. Biotic mechanisms of community stability shift along a precipitation gradient. Ecology 2014, 95, 1693–1700. [Google Scholar] [CrossRef] [Green Version]
- Pearson, K. On lines and planes of closest fit to systems of points in space. Lond. Edinb. Dubl. Philos. Mag. 1901, 2, 559–572. [Google Scholar] [CrossRef] [Green Version]
- Kruskal, J.B. Nonmetric multidimensional scaling: A numerical method. Psychometrika 1964, 29, 115–129. [Google Scholar] [CrossRef]
- Gower, J.C. Some distance properties of latent root and vector methods used in multivariate analysis. Biometrika 1966, 53, 322–338. [Google Scholar] [CrossRef]
- Hill, M.O.; Gauch, H.G., Jr. Detrended correspondence analysis: An improved ordination technique. Vegetation 1980, 42, 42–58. [Google Scholar] [CrossRef]
- Hill, M.O. Correspondence analysis: A neglected multivariate method. J. R. Stat. Soc. C Appl. 1974, 23, 340–354. [Google Scholar] [CrossRef]
- Van Den Wollenberg, A.L. Redundancy analysis an alternative for canonical correlation analysis. Psychometrika 1977, 42, 207–219. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F. Canonical Correspondence Analysis: A New Eigenvector Technique for Multivariate Direct Gradient Analysis. Ecology 1986, 67, 1167–1179. [Google Scholar] [CrossRef] [Green Version]
- Ko, H.S.; Jhun, M.S.; Jeong, H.C. A Comparion Study for Ordination Methods in Ecology. Korean J. Appl. Stat. 2015, 28, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Potapova, M.G.; Charles, D.F. Benthic diatoms in USA rivers: Distributions along spatial and environmental gradients. J. Biogeogr. 2002, 29, 167–187. [Google Scholar] [CrossRef]
- Lange-Bertalot, H. 85 Neue Taxa und über 100 weitre neudefinierte taxa ergänzend zur Süβwasserflora von Mitteleuropa vol. 2/1-4. Biblioth. Diatomol. 1993, 27, 1–454. [Google Scholar]
- Watanabe, T. Picture Book and Ecology of the Freshwater Diatoms; Uchida Rokakuho publishing Co: Tokyo, Japan, 2005. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Süsswasserflora von Mitteleuropa, Band 2/1: Bacillariophyceae. 2. Teil. Bacillariaceae, Epithemiaceae, Surirellaceae; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Verlag, Stuttgart: New York, NY, USA, 1988. [Google Scholar]
- Taylor, J.C.; Harding, W.R.; Archibald, C.G.M. An Illustrated Guide to Some Common Diatom Species from South Africa. Water Research Commission; WRC Report TT 282/07; RSA: Pretoria, South Africa, 2007. [Google Scholar]
- Kützing, F.T. Synopsis Diatomacearum oder Versuch einer systematischen Zusammentellung der Diatomeen. Linnaea 1833, 8, 529–620. [Google Scholar]
- Zhang, J.; Shu, X.; Zhang, Y.; Tan, X.; Zhang, Q. The responses of epilithic algal community structure and function to light and nutrients and their linkages in subtropical rivers. Hydrobiologia 2020, 847, 841–855. [Google Scholar] [CrossRef]
- Tan, X.; Zhang, Q.; Burford, M.A.; Sheldon, F.; Bunn, S.E. Benthic Diatom Based Indices for Water Quality Assessment in Two Subtropical Streams. Front. Microbiol. 2017, 8, 601. [Google Scholar] [CrossRef] [Green Version]
- Meybeck, M.; Friedrich, G.; Thomas, R.; Chapman, D. Rivers; Water Quality Assessments. A Guide to the use of Biota, Sediments and Water in Environmental Monitoring; Chapman, D., Ed.; Chapman and Hall: Cambridge, UK, 1992. [Google Scholar]
- Prathumratana, L.; Sthiannopkao, S.; Kim, K.W. The relationship of climatic and hydrological parameters to surface water quality in the lower Mekong River. Environ. Int. 2008, 34, 860–866. [Google Scholar] [CrossRef] [PubMed]
- Coulliette, A.D.; Noble, R.T. Impacts of rainfall on the water quality of the Newport River Estuary (Eastern North Carolina, USA). J. Water. Health 2008, 6, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.R.; Drever, J.I. Water chemistry of a stream following a storm, Absaroka Mountains, Wyoming. Geol. Soc. Am. Bull. 1977, 88, 286–290. [Google Scholar] [CrossRef]
- Cornish, P.M. The Variations of Dissolved Ion Concentration with Discharge in Some New South Wales Streams; The First National Symposium on Forest Hydrology: Melbourne, Australia, 1982.
- Muraoka, K.; Hirata, T. Streamwater chemistry during rainfall events in a forested basin. J. Hydrol. 1988, 102, 235–253. [Google Scholar] [CrossRef]
- Jénkins, A. Storm period hydrochemical response in an unforested Scottish catchment. Hydrol. Sci. J. 1989, 34, 393–404. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.J.; Kim, N.Y.; Yoon, S.A.; Kim, B.H.; Park, M.H.; You, K.A.; Lee, H.Y.; Kim, H.S.; Kim, Y.J.; Lee, J.; et al. Distribution of benthic diatoms in Korean rivers and streams in relation to environmental variables. Ann. Limnol. Int. J. Lim. 2011, 47, S15–S33. [Google Scholar] [CrossRef] [Green Version]
- Feio, M.J.; Dolédec, S.; Graça, M.A.S. Human disturbance affects the long-term spatial synchrony of freshwater invertebrate communities. Environ. Pollut. 2015, 196, 300–308. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | G1 | G2 | G3 | F | p |
|---|---|---|---|---|---|
| WT | 18.8 ± 3.0a | 22.7 ± 3.0b | 24.9 ± 2.8c | 83.457 | <0.001 |
| DO | 8.7 ± 1.0 | 9.3 ± 1.5 | 9.2 ± 2.6 | 2.829 | 0.061 |
| pH | 7.3 ± 0.5a | 7.9 ± 0.6c | 7.7 ± 0.5b | 26.001 | <0.001 |
| EC | 97 ± 59a | 171 ± 119a | 728 ± 564b | 89.128 | <0.001 |
| Turb | 8.5 ± 6.4a | 18.6 ± 9b | 26.7 ± 12.4c | 73.017 | <0.001 |
| BOD | 0.8 ± 0.4a | 1.7 ± 0.9a | 7.3 ± 5.4b | 103.037 | <0.001 |
| NH3-N | 0.21 ± 0.24a | 0.59 ± 0.67a | 2.84 ± 2.78b | 61.559 | <0.001 |
| NO3-N | 1.24 ± 0.54a | 1.75 ± 0.64b | 1.84 ± 1.02b | 15.715 | <0.001 |
| TN | 2.37 ± 0.88a | 3.30 ± 1.23b | 6.28 ± 3.22c | 80.512 | <0.001 |
| PO4-P | 0.02 ± 0.02a | 0.07 ± 0.07b | 0.16 ± 0.15c | 41.414 | <0.001 |
| TP | 0.04 ± 0.04a | 0.11 ± 0.11b | 0.25 ± 0.25c | 41.944 | <0.001 |
| Biomass | 29.3 ± 32.7 | 27.2 ± 27.0 | 31.8 ± 29.4 | 0.412 | 0.663 |
| No. species | 28.5 ± 7.9b | 37.7 ± 9.2c | 23.2 ± 6.5a | 61.606 | <0.001 |
| Dominance (DI) | 0.55 ± 0.16a | 0.50 ± 0.16a | 0.63 ± 0.16b | 12.076 | <0.001 |
| Diversity (H’) | 3.06 ± 0.73b | 3.42 ± 0.79c | 2.64 ± 0.74a | 19.424 | <0.001 |
| Richness (j) | 2.29 ± 0.70b | 3.06 ± 0.87c | 1.80 ± 0.54a | 55.323 | <0.001 |
| Evenness (e) | 0.64 ± 0.11b | 0.66 ± 0.12b | 0.59 ± 0.13a | 6.233 | 0.002 |
| Years | G1 | G2 | G3 | |
|---|---|---|---|---|
| 2012 | BF | Achnanthidium alteragracillimum (36.1) | Nitzschia fonticola (43.2) | Nitzschia palea (19.1) |
| AF | Achnanthidium convergens (22.5) | Nitzschia palea (14.0) | Nitzschia palea (27.6) | |
| 2013 | BF | Achnanthidium convergens (38.5) | Nitzschia fonticola (26.0) | Nitzschia fonticola (27.6) |
| AF | Achnanthidium convergens (43.3) | Achnanthidium minutissimum (16.1) | Nitzschia palea (45.2) | |
| 2014 | BF | Achnanthidium convergens (30.8) | Nitzschia palea (54.4) | Nitzschia palea (22.4) |
| AF | Achnanthidium minutissimum (29.8) | Nitzschia palea (20.2) | Nitzschia palea (26.8) | |
| 2015 | BF | Achnanthidium alteragracillimum (26.7) | Nitzschia fonticola (31.2) | Nitzschia fonticola (31.5) |
| AF | Achnanthidium minutissimum (27.3) | Fragilaria elliptica (20.0) | Nitzschia palea (26.3) |
| DS | Rain | WT | DO | pH | EC | Turb. | BOD | NH3-N | NO3-N | TN | PO4-P | TP | Group |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ACCN | - | −0.55** | - | −0.25** | −0.59** | −0.37** | −0.60** | −0.58** | −0.36** | −0.63** | −0.48** | −0.48** | G1 |
| ACAL | - | −0.36** | - | −0.13* | −0.41** | −0.32** | −0.51** | −0.55** | −0.30** | −0.55** | −0.44** | −0.45** | G1 |
| ACMI | - | −0.34** | - | −0.24** | −0.53** | −0.33** | −0.49** | −0.54** | −0.25** | −0.55** | −0.36** | −0.38** | G1 |
| NIFO | −0.29** | −0.22** | 0.15* | - | −0.16* | - | −0.21** | −0.32** | 0.19** | −0.18** | −0.19** | −0.17** | G2 |
| NIPA | 0.23** | 0.49** | - | 0.23** | 0.49** | 0.39** | 0.45** | 0.41** | 0.31** | 0.42** | 0.29** | 0.29** | G3 |
| NAMI | - | 0.23** | - | - | 0.32** | 0.21** | 0.29** | 0.25** | 0.25** | 0.26** | 0.21** | 0.23** | G3 |
| Biological Indices | CDI |
|---|---|
| Abundance | −0.03 |
| No. of species | 0.44 ** |
| Dominant index | −0.31 ** |
| Diversity index | 0.35 ** |
| Richness index | 0.41 ** |
| Evenness index | 0.28 ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, I.-H.; Kim, H.-K.; Lee, M.-H.; Kim, Y.-J.; Lee, H.; Kim, B.-H. The Effect of Monsoon Rainfall Patterns on Epilithic Diatom Communities in the Hantangang River, Korea. Water 2020, 12, 1471. https://doi.org/10.3390/w12051471
Cho I-H, Kim H-K, Lee M-H, Kim Y-J, Lee H, Kim B-H. The Effect of Monsoon Rainfall Patterns on Epilithic Diatom Communities in the Hantangang River, Korea. Water. 2020; 12(5):1471. https://doi.org/10.3390/w12051471
Chicago/Turabian StyleCho, In-Hwan, Ha-Kyung Kim, Min-Hyuk Lee, Yong-Jae Kim, Hyuk Lee, and Baik-Ho Kim. 2020. "The Effect of Monsoon Rainfall Patterns on Epilithic Diatom Communities in the Hantangang River, Korea" Water 12, no. 5: 1471. https://doi.org/10.3390/w12051471
APA StyleCho, I. -H., Kim, H. -K., Lee, M. -H., Kim, Y. -J., Lee, H., & Kim, B. -H. (2020). The Effect of Monsoon Rainfall Patterns on Epilithic Diatom Communities in the Hantangang River, Korea. Water, 12(5), 1471. https://doi.org/10.3390/w12051471