A Review of Algae-Based Produced Water Treatment for Biomass and Biofuel Production




Abstract
:1. Introduction
2. Algae-Based PW Systems
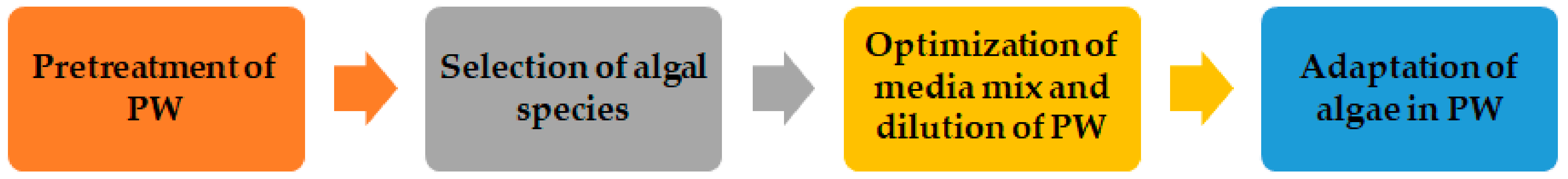
2.1. Cultivation of Algae in PW for Biomass Production
2.1.1. Pre-Treatment of PW
2.1.2. Selection of Algal Species Based on the Salinity of PW
2.1.3. Adaptation of Algae in PW
2.1.4. Addition of Media Mixes and Nutrients to PW
2.1.5. Dilution of PW for Optimal Growth of Algae
2.2. Algal-Based Biofuel Production Using PW
2.3. Bioremediation of PW Using Algae
2.4. Post-Processing of Algae and PW
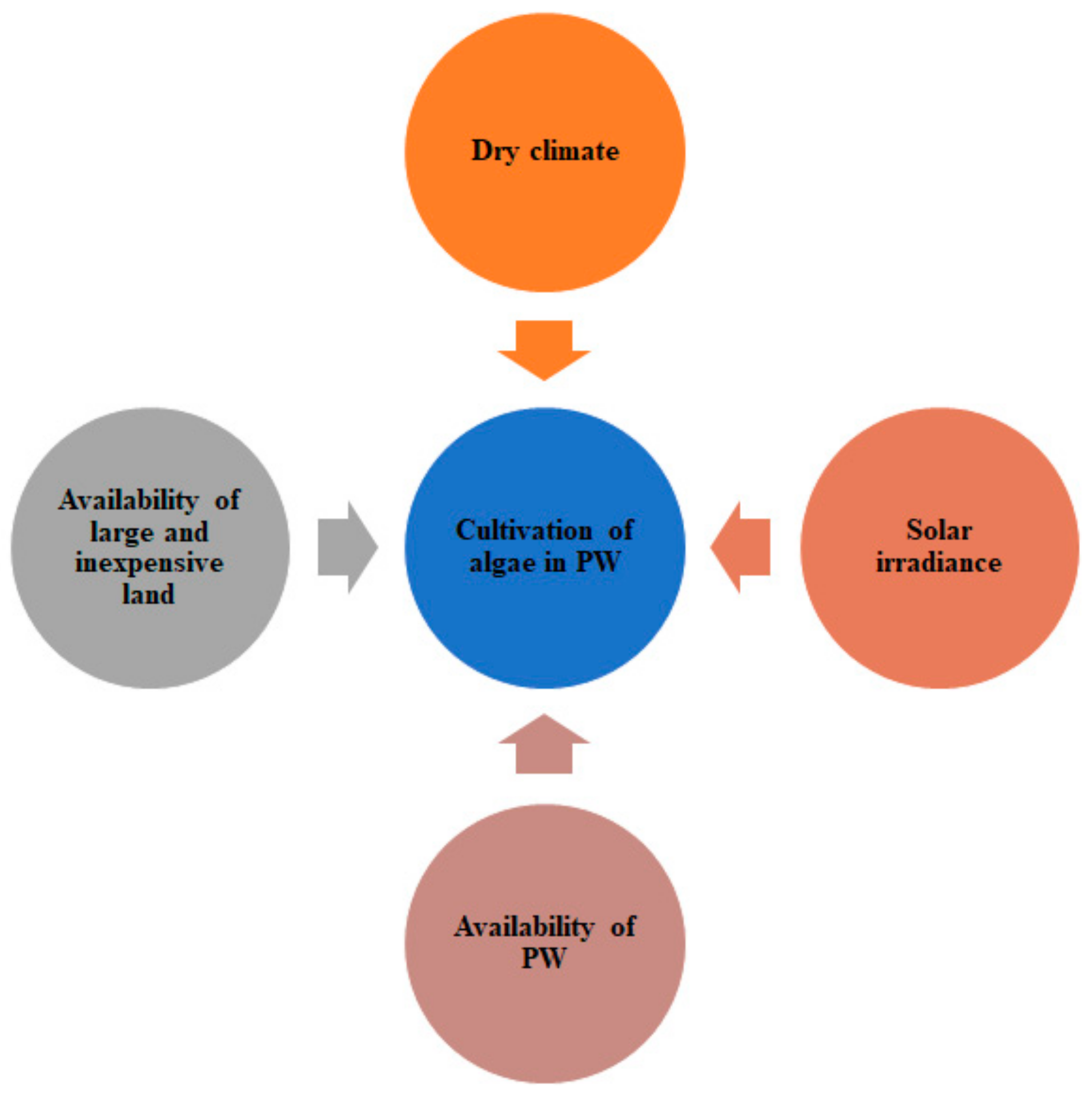
3. The Large-Scale Development of Algae-Based PW System
3.1. Factors Affecting the Large-Scale Development
3.2. Challenges and Future Prospectives
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bakke, T.; Klungsøyr, J.; Sanni, S. Environmental impacts of produced water and drilling waste discharges from the Norwegian offshore petroleum industry. Mar. Environ. Res. 2013, 92, 154–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igunnu, E.T.; Chen, G.Z. Produced water treatment technologies. Int. J. Low Carbon Technol. 2014, 9, 157–177. [Google Scholar] [CrossRef] [Green Version]
- Brasil, B.; Silva, F.C.P.; Siqueira, F.G. Microalgae biorefineries: The Brazilian scenario in perspective. New BioTechnol. 2017, 39, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Das, P.; AbdulQuadir, M.; Thaher, M.; Khan, S.; Chaudhary, A.K.; Alghasal, G.; Al-Jabri, H.M.S. Microalgal bioremediation of petroleum-derived low salinity and low pH produced water. J. Appl. Phycol. 2019, 31, 435–444. [Google Scholar] [CrossRef]
- Campos, J.C.; Borges, R.M.H.; Filho, A.O.; Nobrega, R.; Sant’Anna, G.L., Jr. Oilfield wastewater treatment by combined microfiltration and biological processes. Water Res. 2002, 36, 95–104. [Google Scholar] [CrossRef]
- Fakhru’l-Razi, A.; Pendashteh, A.; Abdullah, L.C.; Biak, D.R.A.; Madaeni, S.S.; Abidin, Z.Z. Review of technologies for oil and gas produced water treatment. J. Hazard Mater. 2009, 170, 530–551. [Google Scholar] [CrossRef]
- Chen, L.; Xu, Q.; Gossage, J.L.; Lou, H.H. Simulation and economic evaluation of a coupled thermal vapor compression desalination process for produced water management. J. Nat. Gas Sci. Eng. 2016, 36, 442–453. [Google Scholar] [CrossRef]
- Bayati, F.; Shayegan, J.; Noorjahan, A. Treatment of oilfield produced water by dissolved air precipitation/solvent sublation. J. Petrol Sci. Eng. 2011, 80, 26–31. [Google Scholar] [CrossRef]
- Fathy, M.; El-Sayed, M.; Ramzi, M.; Abdelraheem, O.H. Adsorption separation of condensate oil from produced water using ACTF prepared of oil palm leaves by batch and fixed bed techniques. Egypt J. Petrol 2018, 27, 319–326. [Google Scholar] [CrossRef]
- Jiang, Q.; Rentschler, J.; Perrone, R.; Liu, K. Application of ceramic membrane and ion-exchange for the treatment of the flowback water from Marcellus shale gas production. J. Membrane Sci. 2013, 431, 55–61. [Google Scholar] [CrossRef]
- Al-Ghouti, M.A.; Al-Kaabi, M.A.; Ashfaq, M.Y.; Da’na, D.A. Produced water characteristics, treatment and reuse: A review. J. Water Process. Eng. 2019, 28, 222–239. [Google Scholar] [CrossRef]
- Lu, M.; Zhang, Z.; Yu, W.; Zhu, W. Biological treatment of oilfield-produced water: A field pilot study. Int. Biodeter. Biodegr. 2009, 63, 316–321. [Google Scholar] [CrossRef]
- Chavan, A.; Mukherji, S. Treatment of hydrocarbon-rich wastewater using oil degrading bacteria and phototrophic microorganisms in rotating biological contactor: Effect of N:P ratio. J. Hazard Mater. 2008, 154, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Selvaratnam, T.; Pegallapati, A.K.; Montelya, F.; Rodriguez, G.; Nirmalakhandan, N.; Van Voorhies, W.; Lammers, P.J. Evaluation of a thermo-tolerant acidophilic alga, Galdieria sulphuraria, for nutrient removal from urban wastewaters. Bioresour. Technol. 2014, 156, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Luo, S.; Fan, X.; Yang, Z.; Guo, R. Biomass and lipid production of marine microalgae using municipal wastewater and high concentration of CO2. Appl. Energy 2011, 88, 3336–3341. [Google Scholar] [CrossRef]
- Henkanatte-Gedera, S.M.; Selvaratnam, T.; Caskan, N.; Nirmalakhandan, N.; Van Voorhies, W.; Lammers, P.J. Algal-based, single-step treatment of urban wastewaters. Bioresour. Technol. 2015, 189, 273–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawaz, T.; Rahman, A.; Pan, S.; Dixon, K.; Petri, B.; Selvaratnam, T. A review of landfill leachate treatment by microalgae: Current status and future directions. Processes 2020, 8, 384. [Google Scholar] [CrossRef] [Green Version]
- Cai, T.; Ge, X.; Park, S.Y.; Li, Y. Comparison of Synechocystis sp. PCC6803 and Nannochloropsis salina for lipid production using artificial seawater and nutrients from anaerobic digestion effluent. Bioresour. Technol. 2013, 144, 255–260. [Google Scholar] [CrossRef]
- Hidaka, T.; Inoue, K.; Suzuki, Y.; Tsumori, J. Growth and anaerobic digestion characteristics of microalgae cultivated using various types of sewage. Bioresour. Technol. 2014, 170, 83–89. [Google Scholar] [CrossRef]
- Rahman, A.; Kumar, S.; Bafana, A.; Lin, J.; Dahoumane, S.A.; Jeffryes, C. A mechanistic view of the light-induced synthesis of silver nanoparticles using extracellular polymeric substances of Chlamydomonas reinhardtii. Molecules 2019, 24, 3506. [Google Scholar] [CrossRef] [Green Version]
- Klimmek, S.; Stan, H.-J.; Wilke, A.; Bunke, G.; Buchholz, R. Comparative analysis of the biosorption of cadmium, lead, nickel, and zinc by algae. Environ. Sci. Technol. 2001, 35, 4283–4288. [Google Scholar] [CrossRef] [PubMed]
- Krejci, M.R.; Finney, L.; Vogt, S.; Joester, D. Selective sequestration of strontium in desmid green algae by biogenic co-precipitation with barite. Chemsuschem 2011, 4, 470–473. [Google Scholar] [CrossRef] [PubMed]
- Mehta, S.K.; Gaur, J.P. Use of algae for removing heavy metal ions from wastewater: Progress and prospects. Cr. Rev. Biotechnol. 2005, 25, 113–152. [Google Scholar] [CrossRef]
- Rahman, A.; Lin, J.; Jaramillo, F.E.; Bazylinski, D.A.; Jeffryes, C.; Dahoumane, S.A. In vivo biosynthesis of inorganic nanomaterials using eukaryotes—A Review. Molecules 2020, 25, 3246. [Google Scholar] [CrossRef]
- Mishra, A.; Jha, B. Isolation and characterization of extracellular polymeric substances from micro-algae Dunaliella salina under salt stress. Bioresour. Technol. 2009, 100, 3382–3386. [Google Scholar] [CrossRef]
- Selvaratnam, T.; Pegallapati, A.; Montelya, F.; Rodriguez, G.; Nirmalakhandan, N.; Lammers, P.J.; van Voorhies, W. Feasibility of algal systems for sustainable wastewater treatment. Renew. Energy 2015, 82, 71–76. [Google Scholar] [CrossRef] [Green Version]
- Lammers, P.J.; Huesemann, M.; Boeing, W.; Anderson, D.B.; Arnold, R.G.; Bai, X.; Bhole, M.; Brhanavan, Y.; Brown, L.; Brown, J.; et al. Review of the cultivation program within the National Alliance for Advanced Biofuels and Bioproducts. Algal. Res. 2017, 22, 166–186. [Google Scholar] [CrossRef]
- Harkness, J.S.; Dwyer, G.S.; Warner, N.R.; Parker, K.M.; Mitch, W.A.; Vengosh, A. Iodide, bromide, and ammonium in hydraulic fracturing and oil and gas wastewaters: Environmental implications. Environ. Sci. Technol. 2015, 49, 1955–1963. [Google Scholar] [CrossRef] [PubMed]
- Fakhru’l-Razi, A.; Pendashteh, A.; Abidin, Z.Z.; Abdullah, L.C.; Biak, D.R.A.; Madaeni, S.S. Application of membrane-coupled sequencing batch reactor for oilfield produced water recycle and beneficial re-use. Bioresour. Technol. 2010, 101, 6942–6949. [Google Scholar] [CrossRef]
- Riley, S.M.; Oliveira, J.M.S.; Regnery, J.; Cath, T.Y. Hybrid membrane bio-systems for sustainable treatment of oil and gas produced water and fracturing flowback water. Sep. Purif. Technol. 2016, 171, 297–311. [Google Scholar] [CrossRef] [Green Version]
- Pendashteh, A.R.; Abdullah, L.C.; Fakhru’l-Razi, A.; Madaeni, S.S.; Zainal Abidin, Z.; Awang Biak, D.R. Evaluation of membrane bioreactor for hypersaline oily wastewater treatment. Process. Saf. Environ. 2012, 90, 45–55. [Google Scholar] [CrossRef]
- Neal, J.N.; Sullivan, E.J.; Dean, C.A.; Steichen, S.A. Recycling Produced Water for Algal Cultivation for Biofuels; Los Alamos National Lab. (LANL): Los Alamos, NM, USA, 2012. [Google Scholar]
- Markou, G.; Georgakakis, D. Cultivation of filamentous cyanobacteria (blue-green algae) in agro-industrial wastes and wastewaters: A review. Appl. Energy 2011, 88, 3389–3401. [Google Scholar] [CrossRef]
- Eriksen, N.T. Production of phycocyanin—A pigment with applications in biology, biotechnology, foods and medicine. Appl. Microbiol. Biotechnol. 2008, 80, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sekar, S.; Chandramohan, M. Phycobiliproteins as a commodity: Trends in applied research, patents and commercialization. J. Appl. Phycol. 2008, 20, 113–136. [Google Scholar] [CrossRef]
- Chu, W.-L. Biotechnological applications of microalgae. Int. J. Sci. Med. Edu. 2012, 6, S24–S37. [Google Scholar]
- Reddy, H.K.; Muppaneni, T.; Ponnusamy, S.; Sudasinghe, N.; Pegallapati, A.; Selvaratnam, T.; Seger, M.; Dungan, B.; Nirmalakhandan, N.; Schaub, T.; et al. Temperature effect on hydrothermal liquefaction of Nannochloropsis gaditana and Chlorella sp. Appl. Energy 2016, 165, 943–951. [Google Scholar] [CrossRef] [Green Version]
- Selvaratnam, T.; Reddy, H.; Muppaneni, T.; Holguin, F.O.; Nirmalakhandan, N.; Lammers, P.J.; Deng, S. Optimizing energy yields from nutrient recycling using sequential hydrothermal liquefaction with Galdieria sulphuraria. Algal. Res. 2015, 12, 74–79. [Google Scholar] [CrossRef] [Green Version]
- Ranjbar, S.; Quaranta, J.D.; Tehrani, R.; Van Aken, B. Algae-Based Treatment of Hydraulic Fracturing Produced Water: Metal Removal and Biodiesel Production by the Halophilic Microalgae Dunaliella salina. In Bioremediation and Sustainable Environmental Technologies. In Proceedings of the Third International Symposium on Bioremediation and Sustainable Environmental Technologies, Miami, FL, USA, 18–21 May 2015. [Google Scholar]
- Sullivan Graham, E.J.; Dean, C.A.; Yoshida, T.M.; Twary, S.N.; Teshima, M.; Alvarez, M.A.; Zidenga, T.; Heikoop, J.M.; Perkins, G.B.; Rahn, T.A.; et al. Oil and gas produced water as a growth medium for microalgae cultivation: A review and feasibility analysis. Algal. Res. 2017, 24, 492–504. [Google Scholar] [CrossRef]
- Hopkins, T.C.; Graham, E.J.S.; Schwilling, J.; Ingram, S.; Gómez, S.M.; Schuler, A.J. Effects of salinity and nitrogen source on growth and lipid production for a wild algal polyculture in produced water media. Algal. Res. 2019, 38, 101406. [Google Scholar] [CrossRef]
- Arriada, A.A.; Abreu, P.C. Nannochloropsis oculata growth in produced water: An alternative for massive microalgae biomass production. Braz. J. Petrol Gas 2014, 8, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Almaraz, N.; Regnery, J.; Vanzin, G.F.; Riley, S.M.; Ahoor, D.C.; Cath, T.Y. Emergence and fate of volatile iodinated organic compounds during biological treatment of oil and gas produced water. Sci. Total Environ. 2020, 699, 134202. [Google Scholar] [CrossRef]
- Nie, H.; Nie, M.; Diwu, Z.; Wang, L.; Yan, H.; Lin, Y.; Zhang, B.; Wang, Y. Biological treatment of high salinity and low pH produced water in oilfield with immobilized cells of P. aeruginosa NY3 in a pilot-scale. J. Hazard Mater. 2020, 381, 121232. [Google Scholar] [CrossRef] [PubMed]
- Ammar, S.H.; Khadim, H.J.; Mohamed, A.I. Cultivation of Nannochloropsis oculata and Isochrysis galbana microalgae in produced water for bioremediation and biomass production. Environ. Technol. Innov. 2018, 10, 132–142. [Google Scholar] [CrossRef]
- Talebi, A.F.; Dastgheib, S.M.M.; Tirandaz, H.; Ghafari, A.; Alaie, E.; Tabatabaei, M. Enhanced algal-based treatment of petroleum produced water and biodiesel production. RSC Adv. 2016, 6, 47001–47009. [Google Scholar] [CrossRef]
- Godfrey, V. Production of Biodiesel from Oleaginous Organisms Using Underutilized Wastewaters. Master’s Thesis, Utah State University, Logan, UT, USA, 2012. [Google Scholar]
- “Production of Biodiesel from Oleaginous Organisms Using Underutilized” by Valerie Godfrey. Available online: https://digitalcommons.usu.edu/etd/1337/ (accessed on 23 July 2020).
- Winckelmann, D.; Bleeke, F.; Thomas, B.; Elle, C.; Klöck, G. Open pond cultures of indigenous algae grown on non-arable land in an arid desert using wastewater. Int. Aquat. Res. 2015, 7, 221–233. [Google Scholar] [CrossRef] [Green Version]
- Badrinarayanan, I. Using Produced Water to Grow Microalgae; Russel School of Chemical Engineering the University of Tulsa: Tulsa, OK, USA, 2017; p. 20. [Google Scholar]
- Johnson, R.J.; Jurawan, I.; Frenzel, M.; Price, A.C. The identification and mechanism of a Scenedesmus spp. causing bio-fouling of an oil field produced water treatment plant. Int. Biodeter. Biodegr. 2016, 108, 207–213. [Google Scholar] [CrossRef]
- Peng, J.; Yin, K.; Yuan, J.-P.; Cao, G.-X.; Xue, M.; Wu, C.-F.; Wang, J.-H. Characterization of a newly isolated green microalga Scenedesmus sp. as a potential source of biodiesel. Afr. J. Biotechnol. 2012, 11, 16083–16094. [Google Scholar]
- Büdel, B.; Darienko, T.; Deutschewitz, K.; Dojani, S.; Friedl, T.; Mohr, K.I.; Salisch, M.; Reisser, W.; Weber, B. Southern African biological soil crusts are ubiquitous and highly diverse in drylands, being restricted by rainfall frequency. Microbial. Ecol. 2009, 57, 229–247. [Google Scholar] [CrossRef]
- Eliaš, M.; Němcová, Y.; Škaloud, P.; Neustupa, J.; Kaufnerova, V.; Šejnohová, L. Hylodesmus singaporensis gen. et sp. nov., a new autosporic subaerial green alga (Scenedesmaceae, Chlorophyta) from Singapore. Int. J. Syst. Evol. Micr. 2010, 60, 1224–1235. [Google Scholar] [CrossRef] [Green Version]
- Lewis, L.A.; Flechtner, V.R. Green algae (Chlorophyta) of desert microbiotic crusts: Diversity of North American taxa. Taxon 2002, 51, 443–451. [Google Scholar] [CrossRef]
- Lewis, L.A.; Flechtner, V.R. Cryptic species of Scenedesmus (chlorophyta) from desert soil communities of Western North America. J. Phycol. 2004, 40, 1127–1137. [Google Scholar] [CrossRef]
- Bartley, M.L.; Boeing, W.J.; Corcoran, A.A.; Holguin, F.O.; Schaub, T. Effects of salinity on growth and lipid accumulation of biofuel microalga Nannochloropsis salina and invading organisms. Biomass Bioenergy 2013, 54, 83–88. [Google Scholar] [CrossRef]
- Racharaks, R.; Ge, X.; Li, Y. Cultivation of marine microalgae using shale gas flowback water and anaerobic digestion effluent as the cultivation medium. Bioresour. Technol. 2015, 191, 146–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aravinthan, V.; Harrington, D. Coal seam gas water as a medium to grow Dunalliella tertiolecta microalgae for lipid extraction. Desalin. Water Treat 2014, 52, 947–958. [Google Scholar] [CrossRef]
- Hopkins, T.C.; Graham, E.J.S.; Schuler, A.J. Biomass and lipid productivity of Dunaliella tertiolecta in a produced water-based medium over a range of salinities. J. Appl. Phycol. 2019, 31, 3349–3358. [Google Scholar] [CrossRef]
- Calderón-Delgado, I.C.; Mora-Solarte, D.A.; Velasco-Santamaría, Y.M. Physiological and enzymatic responses of Chlorella vulgaris exposed to produced water and its potential for bioremediation. Environ. Monit. Assess 2019, 191, 399. [Google Scholar] [CrossRef]
- Mendes, L.B.B.; Cunha, P.C.R.; D’oca, M.G.M.; Abreu, P.C.; Primel, E.G. Method for Removing Pollutants from Produced Water. U.S. Patent US 7,955,505, 7 June 2011. [Google Scholar]
- Mohd Abdul Hakim, M.A.Q. A Potential Application of Microalgae in Produced Water Treatment. Master’s Thesis, Qatar University, Doha, Qatar, 2016. [Google Scholar]
- Andrade, B.G.; Andrade, V.T.; Costa, B.R.; Campos, J.C.; Dezotti, M. Distillation of oil field produced water for reuse on irrigation water: Evaluation of pollutants removal and ecotoxicity. J. Water Reuse Desal. 2011, 1, 224–236. [Google Scholar] [CrossRef]
- Wood, J.L.; Miller, C.D.; Sims, R.C.; Takemoto, J.Y. Biomass and phycocyanin production from cyanobacteria dominated biofilm reactors cultured using oilfield and natural gas extraction produced water. Algal Res. 2015, 11, 165–168. [Google Scholar] [CrossRef]
- Vento, S.D.; Dachs, J. Prediction of uptake dynamics of persistent organic pollutants by bacteria and phytoplankton. Environ. Toxicol. Chem. 2002, 21, 2099–2107. [Google Scholar] [CrossRef]
- Fan, C.-W.; Reinfelder, J.R. Phenanthrene accumulation kinetics in marine diatoms. Environ. Sci. Technol. 2003, 37, 3405–3412. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Blain, R.B. Hormesis and plant biology. Environ. Pollut. 2009, 157, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Spoljaric, D.; Cipak, A.; Horvatic, J.; Andrisic, L.; Waeg, G.; Zarkovic, N.; Jaganjac, M. Endogenous 4-hydroxy-2-nonenal in microalga Chlorella kessleri acts as a bioactive indicator of pollution with common herbicides and growth regulating factor of hormesis. Aquat. Toxicol. 2011, 105, 552–558. [Google Scholar] [CrossRef]
- Franqueira, D.; Orosa, M.; Torres, E.; Herrero, C.; Cid, A. Potential use of flow cytometry in toxicity studies with microalgae. Sci. Total Environ. 2000, 247, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Witt, G. Occurrence and transport of polycyclic aromatic hydrocarbons in the water bodies of the Baltic Sea. Mar. Chem. 2002, 79, 49–66. [Google Scholar] [CrossRef]
- Ramanan, R.; Kim, B.-H.; Cho, D.-H.; Oh, H.-M.; Kim, H.-S. Algae–bacteria interactions: Evolution, ecology and emerging applications. BioTechnol. Adv. 2016, 34, 14–29. [Google Scholar] [CrossRef] [Green Version]
- Gorain, P.C.; Bagchi, S.K.; Mallick, N. Effects of calcium, magnesium and sodium chloride in enhancing lipid accumulation in two green microalgae. Environ. Technol. 2013, 34, 1887–1894. [Google Scholar] [CrossRef]
- Affenzeller, M.J.; Darehshouri, A.; Andosch, A.; Lutz, C.; Lutz-Meindl, U. Salt stress-induced cell death in the unicellular green alga Micrasterias denticulata. J. Exp. Bot. 2009, 60, 939–954. [Google Scholar] [CrossRef] [Green Version]
- Salama, E.-S.; Kim, H.-C.; Abou-Shanab, R.A.I.; Ji, M.-K.; Oh, Y.-K.; Kim, S.-H.; Jeon, B.-H. Biomass, lipid content, and fatty acid composition of freshwater Chlamydomonas mexicana and Scenedesmus obliquus grown under salt stress. Bioprocess Biosyst. Eng. 2013, 36, 827–833. [Google Scholar] [CrossRef]
- O’Connell, D.W.; Birkinshaw, C.; O’Dwyer, T.F. Heavy metal adsorbents prepared from the modification of cellulose: A review. Bioresour. Technol. 2008, 99, 6709–6724. [Google Scholar] [CrossRef]
- Rahman, A.; Kumar, S.; Bafana, A.; Dahoumane, S.; Jeffryes, C. Biosynthetic conversion of Ag+ to highly stable Ag0 nanoparticles by wild type and cell wall deficient strains of Chlamydomonas reinhardtii. Molecules 2018, 24, 98. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Chen, C. Biosorbents for heavy metals removal and their future. BioTechnol. Adv 2009, 27, 195–226. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Kumar, S.; Bafana, A.; Dahoumane, S.A.; Jeffryes, C. Individual and combined effects of extracellular polymeric substances and whole cell components of Chlamydomonas reinhardtii on silver nanoparticle synthesis and stability. Molecules 2019, 24, 956. [Google Scholar] [CrossRef] [Green Version]
- Finocchio, E.; Lodi, A.; Solisio, C.; Converti, A. Chromium (VI) removal by methylated biomass of Spirulina platensis: The effect of methylation process. Chem. Eng. J. 2010, 156, 264–269. [Google Scholar] [CrossRef]
- Rahman, A. Biosynthetic conversion of Ag+ to colloidally stable Ag0 nanoparticles by Chlamydomonas reinhardtii: A Mechanistic View of the Light-Induced Synthesis Process and the Impact of Extracellular Polymeric Substances on the Stability of the Nanoparticles. Ph.D. Thesis, Lamar University, Beaumont, TX, USA, 2019. [Google Scholar]
- Rahman, A.; Kumar, S.; Nawaz, T. Chapter 17—Biosynthesis of nanomaterials using algae. In Microalgae Cultivation for Biofuels Production; Yousuf, A., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 265–279. ISBN 978-0-12-817536-1. [Google Scholar]
- Volesky, B. Biosorption and me. Water Res. 2007, 41, 4017–4029. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.; Carmen Bartolomé, M.; Sánchez-Fortún, S. Influence of pH on the survival of Dictyosphaerium chlorelloides populations living in aquatic environments highly contaminated with chromium. Ecotox. Environ. Safe 2013, 98, 82–87. [Google Scholar] [CrossRef] [PubMed]
- López-Rodas, V.; Marvá, F.; Rouco, M.; Costas, E.; Flores-Moya, A. Adaptation of the chlorophycean Dictyosphaerium chlorelloides to stressful acidic, mine metal-rich waters as result of pre-selective mutations. Chemosphere 2008, 72, 703–707. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.S.; Rodrigues, M.S.; de Carvalho, J.C.M.; Lodi, A.; Finocchio, E.; Perego, P.; Converti, A. Adsorption of Ni2+, Zn2+ and Pb2+ onto dry biomass of Arthrospira (Spirulina) platensis and Chlorella vulgaris. I. single metal systems. Chem. Eng. J. 2011, 173, 326–333. [Google Scholar] [CrossRef]
- Ojeda-Barrios, D.; Abadía, J.; Lombardini, L.; Abadía, A.; Vázquez, S. Zinc deficiency in field-grown pecan trees: Changes in leaf nutrient concentrations and structure. J. Sci. Food Agric. 2012, 92, 1672–1678. [Google Scholar] [CrossRef]
- Öncel, I.; Keleş, Y.; Üstün, A.S. Interactive effects of temperature and heavy metal stress on the growth and some biochemical compounds in wheat seedlings. Environ. Pollut. 2000, 107, 315–320. [Google Scholar] [CrossRef]
- Lefebvre, O.; Moletta, R. Treatment of organic pollution in industrial saline wastewater: A literature review. Water Res. 2006, 40, 3671–3682. [Google Scholar] [CrossRef]
- Mahdavi, H.; Prasad, V.; Liu, Y.; Ulrich, A.C. In situ biodegradation of naphthenic acids in oil sands tailings pond water using indigenous algae–bacteria consortium. Bioresour. Technol. 2015, 187, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Guo, X.; Su, G.; Danquah, M.K.; Zhang, S.; Lu, Y.; Sun, Y.; Lin, L. Bioprocess considerations for microalgal-based wastewater treatment and biomass production. Renew. Sustain. Energy Rev. 2015, 42, 1385–1392. [Google Scholar] [CrossRef]
- Richardson, J.W.; Johnson, M.D.; Lacey, R.; Oyler, J.; Capareda, S. Harvesting and extraction technology contributions to algae biofuels economic viability. Algal. Res. 2014, 5, 70–78. [Google Scholar] [CrossRef]
- Brennan, L.; Owende, P. Biofuels from microalgae: Towards meeting advanced fuel standards. In Advanced Biofuels and Bioproducts; Springer: New York, NY, USA, 2013; pp. 553–599. [Google Scholar]
- Coons, J.E.; Kalb, D.M.; Dale, T.; Marrone, B.L. Getting to low-cost algal biofuels: A monograph on conventional and cutting-edge harvesting and extraction technologies. Algal. Res. 2014, 6, 250–270. [Google Scholar] [CrossRef] [Green Version]
- Roselet, F.; Vandamme, D.; Roselet, M.; Muylaert, K.; Abreu, P.C. Screening of commercial natural and synthetic cationic polymers for flocculation of freshwater and marine microalgae and effects of molecular weight and charge density. Algal. Res. 2015, 10, 183–188. [Google Scholar] [CrossRef]
- Schlesinger, A.; Eisenstadt, D.; Bar-Gil, A.; Carmely, H.; Einbinder, S.; Gressel, J. Inexpensive non-toxic flocculation of microalgae contradicts theories; overcoming a major hurdle to bulk algal production. BioTechnol. Adv. 2012, 30, 1023–1030. [Google Scholar] [CrossRef]
- Emamjomeh, M.M.; Sivakumar, M. Review of pollutants removed by electrocoagulation and electrocoagulation/flotation processes. J. Environ. Manag. 2009, 90, 1663–1679. [Google Scholar] [CrossRef]
- Lakaniemi, A.-M.; Tuovinen, O.H.; Puhakka, J.A. Anaerobic conversion of microalgal biomass to sustainable energy carriers—A review. Bioresour. Technol. 2013, 135, 222–231. [Google Scholar] [CrossRef]
- Lee, O.K.; Kim, A.L.; Seong, D.H.; Lee, C.G.; Jung, Y.T.; Lee, J.W.; Lee, E.Y. Chemo-enzymatic saccharification and bioethanol fermentation of lipid-extracted residual biomass of the microalga, Dunaliella tertiolecta. Bioresour. Technol. 2013, 132, 197–201. [Google Scholar] [CrossRef]
- Tang, H.; Abunasser, N.; Garcia, M.E.D.; Chen, M.; Simon Ng, K.Y.; Salley, S.O. Potential of microalgae oil from Dunaliella tertiolecta as a feedstock for biodiesel. Appl. Energy 2011, 88, 3324–3330. [Google Scholar] [CrossRef]
- Kim, S.-S.; Ly, H.V.; Kim, J.; Lee, E.Y.; Woo, H.C. Pyrolysis of microalgae residual biomass derived from Dunaliella tertiolecta after lipid extraction and carbohydrate saccharification. Chem. Eng. J. 2015, 263, 194–199. [Google Scholar] [CrossRef]
- Chen, Y.; Zhao, N.; Wu, Y.; Wu, K.; Wu, X.; Liu, J.; Yang, M. Distributions of organic compounds to the products from hydrothermal liquefaction of microalgae. Environ. Prog. Sustain. 2017, 36, 259–268. [Google Scholar] [CrossRef]
- Richardson, J.W.; Johnson, M.D. Financial feasibility analysis of NAABB developed technologies. Algal. Res. 2015, 10, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Frank, E.D.; Elgowainy, A.; Han, J.; Wang, Z. Life cycle comparison of hydrothermal liquefaction and lipid extraction pathways to renewable diesel from algae. Mitig. Adapt. Strat. Gl. 2013, 18, 137–158. [Google Scholar] [CrossRef] [Green Version]
- Uduman, N.; Qi, Y.; Danquah, M.K.; Forde, G.M.; Hoadley, A. Dewatering of microalgal cultures: A major bottleneck to algae-based fuels. J. Renew. Sustain. Energy 2010, 2, 012701. [Google Scholar] [CrossRef]
- Hu, Y.; Feng, S.; Yuan, Z.; Xu, C.C.; Bassi, A. Investigation of aqueous phase recycling for improving bio-crude oil yield in hydrothermal liquefaction of algae. Bioresour. Technol. 2017, 239, 151–159. [Google Scholar] [CrossRef]
- Chisti, Y. Constraints to commercialization of algal fuels. J. Biotechnol. 2013, 167, 201–214. [Google Scholar] [CrossRef]
- Chaudhary, B.K.; Willman, S.E.; Researcher, P.-M.; Carroll, K.C. Spatial Variability and Geochemistry of Produced Water in Southeastern New Mexico, USA; New Mexico Water Resources Research Institute: Las Cruces, NM, USA, 2016; p. 33. [Google Scholar]
- Ishika, T.; Moheimani, N.R.; Bahri, P.A. Sustainable saline microalgae co-cultivation for biofuel production: A critical review. Renew. Sust. Energy Rev. 2017, 78, 356–368. [Google Scholar] [CrossRef]
- Guieysse, B.; Béchet, Q.; Shilton, A. Variability and uncertainty in water demand and water footprint assessments of fresh algae cultivation based on case studies from five climatic regions. Bioresour. Technol. 2013, 128, 317–323. [Google Scholar] [CrossRef]
- Stephens, E.; Ross, I.L.; King, Z.; Mussgnug, J.H.; Kruse, O.; Posten, C.; Borowitzka, M.A.; Hankamer, B. An economic and technical evaluation of microalgal biofuels. New Biotechnol. 2010, 28, 126–128. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Guria, C.; Pathak, A.K. Potential CO2 fixation and optimal Dunaliella tertiolecta cultivation: Influence of fertilizer, wavelength of light-emitting diodes, salinity and carbon supply strategy. J. CO2 Util. 2017, 22, 164–177. [Google Scholar] [CrossRef]
- Andersen, R.A. Algal Culturing Techniques; Academic Press: Cambridge, MA, USA, 2005; ISBN 978-0-12-088426-1. [Google Scholar]
- Mendes, L.B.B.; Vermelho, A.B. Allelopathy as a potential strategy to improve microalgae cultivation. BioTechnol. Biofuels 2013, 6, 1–14. [Google Scholar]
- Graneli, E.; Weberg, M.; Salomon, P.S. Harmful algal blooms of allelopathic microalgal species: The role of eutrophication. Harmful Algae 2008, 8, 94–102. [Google Scholar] [CrossRef]
- El-Kassas, H.Y.; Khairy, H.M. A trial for biological control of a pathogenic fungus (Fusarium solani) by some marine microorganisms. Environ. Sci. 2009, 8, 434–440. [Google Scholar]
- Lutzu, G.A.; Dunford, N.T. Algal treatment of wastewater generated during oil and gas production using hydraulic fracturing technology. Environ. Technol. 2019, 40, 1027–1034. [Google Scholar] [CrossRef]
- Taikhao, S.; Incharoensakdi, A.; Phunpruch, S. Dark fermentative hydrogen production by the unicellular halotolerant cyanobacterium Aphanothece halophytica grown in seawater. J. Appl. Phycol. 2015, 27, 187–196. [Google Scholar] [CrossRef]
- Papry, R.I.; Ishii, K.; Mamun, M.A.A.; Miah, S.; Naito, K.; Mashio, A.S.; Maki, T.; Hasegawa, H. Arsenic biotransformation potential of six marine diatom species: Effect of temperature and salinity. Sci. Rep.-UK 2019, 9, 10226. [Google Scholar] [CrossRef] [Green Version]
- Shikata, T.; Matsubara, T.; Yoshida, M.; Sakamoto, S.; Yamaguchi, M. Effects of temperature, salinity, and photosynthetic photon flux density on the growth of the harmful diatom Asteroplanus karianus in the Ariake Sea, Japan. Fisheries Sci. 2015, 81, 1063–1069. [Google Scholar] [CrossRef]
- Paul, J.T.; Ramaiah, N.; Sardessai, S. Nutrient regimes and their effect on distribution of phytoplankton in the Bay of Bengal. Mar. Environ. Res. 2008, 66, 337–344. [Google Scholar] [CrossRef] [Green Version]
- Saros, J.E. Changes in the growth rates of saline-lake diatoms in response to variation in salinity, brine type and nitrogen form. J. Plankton Res. 2000, 22, 1071–1083. [Google Scholar] [CrossRef] [Green Version]
- Pahl, S.L.; Lewis, D.M.; Chen, F.; King, K.D. Heterotrophic growth and nutritional aspects of the diatom Cyclotella cryptica (Bacillariophyceae): Effect of some environmental factors. J. Biosci. Bioeng. 2010, 109, 235–239. [Google Scholar] [CrossRef]
- Hevia-Orube, J.; Orive, E.; David, H.; Díez, A.; Laza-Martínez, A.; Miguel, I.; Seoane, S. Molecular and morphological analyses of solitary forms of brackish Thalassiosiroid diatoms (Coscinodiscophyceae), with emphasis on their phenotypic plasticity. Eur J. Phycol. 2016, 51, 11–30. [Google Scholar] [CrossRef] [Green Version]
- Tanimura, Y.; Nagumo, T.; Kato, M. A New Variety of Cyclotella atomus from Tokyo Bay, Japan; C. atomus var. marina var. nov. Bull. Nat. Sci. Mus. 2004, 30, 5–11. [Google Scholar]
- Wen, Z.; Mian-ping, Z.; Xian-zhong, X.; Xi-Fang, L.; Gan-lin, G.; Zhi-hui, H. Biological and ecological features of saline lakes in northern Tibet, China. Hydrobiologia 2005, 541, 189–203. [Google Scholar] [CrossRef]
- Rynearson, T.A.; Newton, J.A.; Armbrust, E.V. Spring bloom development, genetic variation, and population succession in the planktonic diatom Ditylum brightwellii. Limnol. Oceanogr. 2006, 51, 1249–1261. [Google Scholar] [CrossRef]
- Papry, R.I.; Omori, Y.; Fujisawa, S.; Al Mamun, M.A.; Miah, S.; Mashio, A.S.; Maki, T.; Hasegawa, H. Arsenic biotransformation potential of marine phytoplankton under a salinity gradient. Algal. Res. 2020, 47, 101842. [Google Scholar] [CrossRef]
- Xu, N.; Pang, S.; Shan, T.; Liu, F.; Zhao, X.; Gao, S. Molecular identification and culture trials of Eutreptiella gymnastica (Eutreptiales, Euglenophyceae). Chin. J. Oceanol. Limn. 2012, 30, 446–455. [Google Scholar] [CrossRef]
- Jeong, H.J.; Kim, T.H.; Yoo, Y.D.; Yoon, E.Y.; Kim, J.S.; Seong, K.A.; Kim, K.Y.; Park, J.Y. Grazing impact of heterotrophic dinoflagellates and ciliates on common red-tide euglenophyte Eutreptiella gymnastica in Masan Bay, Korea. Harmful Algae 2011, 10, 576–588. [Google Scholar] [CrossRef]
- Carić, M.; Jasprica, N.; Čalić, M.; Batistić, M. Phytoplankton response to high salinity and nutrient limitation in the eastern Adriatic marine lakes. Sci. Mar. 2011, 75, 493–505. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Lambert, W.; Giang, S.; Goericke, R.; Palenik, B. Microalgal assemblages in a poikilohaline pond. J. Phycol. 2014, 50, 303–309. [Google Scholar] [CrossRef]
- Wang, S.; Shi, X.; Palenik, B. Characterization of Picochlorum sp. use of wastewater generated from hydrothermal liquefaction as a nitrogen source. Algal. Res. 2016, 13, 311–317. [Google Scholar] [CrossRef] [Green Version]
- Foflonker, F.; Ananyev, G.; Qiu, H.; Morrison, A.; Palenik, B.; Dismukes, G.C.; Bhattacharya, D. The unexpected extremophile: Tolerance to fluctuating salinity in the green alga Picochlorum. Algal. Res. 2016, 16, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Foflonker, F.; Mollegard, D.; Ong, M.; Yoon, H.S.; Bhattacharya, D. Genomic analysis of Picochlorum species reveals how microalgae may adapt to variable environments. Mol. Biol. Evol. 2018, 35, 2702–2711. [Google Scholar] [CrossRef] [Green Version]
- Pálmai, T.; Szabó, B.; Kotut, K.; Krienitz, L.; Padisák, J. Ecophysiology of a successful phytoplankton competitor in the African flamingo lakes: The green alga Picocystis salinarum (Picocystophyceae). J. Appl. Phycol. 2020, 32, 1813–1825. [Google Scholar] [CrossRef] [Green Version]
- Fanjing, K.; Qinxian, J.; Jia, E.; Mianping, Z. Characterization of a eukaryotic picoplankton alga, strain DGN-Z1, isolated from a soda lake in inner Mongolia, China. Nat. Resour. Env. Iss. 2009, 15, 38. [Google Scholar]
- Krienitz, L.; Bock, C.; Kotut, K.; Luo, W. Picocystis salinarum (Chlorophyta) in saline lakes and hot springs of East Africa. Phycologia 2012, 51, 22–32. [Google Scholar] [CrossRef]
- Brooks, B.W.; James, S.V.; Valenti, T.W., Jr.; Urena-Boeck, F.; Serrano, C.; Berninger, J.P.; Schwierzke, L.; Mydlarz, L.D.; Grover, J.P.; Roelke, D.L. Comparative toxicity of Prymnesium parvum in inland waters. J. Am. Water Resour As. 2010, 46, 45–62. [Google Scholar] [CrossRef]
- Granéli, E.; Salomon, P.S. Factors influencing allelopathy and toxicity in Prymnesium parvum. J. Am. Water Resour As. 2010, 46, 108–120. [Google Scholar] [CrossRef]
- Balzano, S.; Sarno, D.; Kooistra, W.H.C.F. Effects of salinity on the growth rate and morphology of ten Skeletonema strains. J. Plankton Res. 2011, 33, 937–945. [Google Scholar] [CrossRef] [Green Version]
- Demir, N. Changes in the phytoplankton community of a coastal, hyposaline lake in western Anatolia, Turkey. Limnology 2007, 8, 337–342. [Google Scholar] [CrossRef]
- Costa-Böddeker, S.; Thuyên, L.X.; Schwarz, A.; Huy, H.Đ.; Schwalb, A. Diatom assemblages in surface sediments along nutrient and salinity gradients of Thi Vai Estuary and Can Gio Mangrove Forest, Southern Vietnam. Estuar. Coast 2017, 40, 479–492. [Google Scholar] [CrossRef]
- Craggs, R.J.; Heubeck, S.; Lundquist, T.J.; Benemann, J.R. Algal biofuels from wastewater treatment high rate algal ponds. Water Sci. Technol. 2011, 63, 660–665. [Google Scholar] [CrossRef] [PubMed]
- Park, J.B.K.; Craggs, R.J.; Shilton, A.N. Wastewater treatment high rate algal ponds for biofuel production. Bioresour. Technol. 2011, 102, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Vendramel, S.; Bassin, J.P.; Dezotti, M.; Sant’Anna, G.L., Jr. Treatment of petroleum refinery wastewater containing heavily polluting substances in an aerobic submerged fixed-bed reactor. Environ. Technol. 2015, 36, 2052–2059. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Parameter Group | Parameter | Unit | Typical Range | Ref |
|---|---|---|---|---|
| General water quality | pH | S.U. | 6.0–8.5 | [39,40,43,44,45] |
| Density | Kg/m3 | ~1021 | [45] | |
| Salinity | mg/L NaCl | 52,700–633,000 | [39] | |
| Electrical conductivity (EC) | mS/cm | 30–600 | [39,40,43,45] | |
| Turbidity | NTU | ~206 | [45] | |
| Total suspended solids (TSS) | mg L−1 | 20–254 | [39,45,46] | |
| Total dissolved solids (TDS) | mg L−1 | 2650–138,500 | [39,40,44,45,46] | |
| Suspended solids (SS) | mg L−1 | ~550 | [44] | |
| Oil and grease | mg L−1 | 73–540 | [44] | |
| Dissolved organic carbon (DOC) | mg L−1 | 270–470 | [43] | |
| Total organic carbon (TOC) | mg L−1 | 55–63 | [39,44,46] | |
| Total nitrogen (TN) | mg L−1 | 30–34 | [43] | |
| Total phosphorus (TP) | mg L−1 | 0.08–0.2 | [44,46] | |
| Biological oxygen demand (BOD5) | mg L−1 | 5–86 | [44,45] | |
| Chemical oxygen demand (COD) | mg L−1 | 177–1300 | [39,44] | |
| Sodium Adsorption ratio (SAR) | mg L−1 | ~182 | [46] | |
| Polyatomic ions/radicals | Ammonium (NH4+) | mg L−1 | 8–20 | [40,44,46] |
| Bicarbonate (HCO3−) | mg L−1 | 71–263 | [44,45] | |
| Nitrate | mg L−1 | ~622 | [43,46] | |
| Nitrite | mg L−1 | 8.2–9.2 | [43] | |
| Phosphate (PO4) | mg L−1 | 2–300 | [39,46] | |
| Halides | Fluorine | mg L−1 | 0.3–204 | [40] |
| Bromide | mg L−1 | 126–134 | [43] | |
| Chloride | mg L−1 | 750–11,500 | [40,43,44,45,46] | |
| Iodide | mg L−1 | 46–48 | [43] | |
| Cations | Aluminium | mg L−1 | 0.01–1.5 | [39,40] |
| Barium | mg L−1 | 0.1–172 | [39,40] | |
| Boron | mg L−1 | 5–11 | [40] | |
| Calcium | mg L−1 | 4–7800 | [39,40,43,44,46] | |
| Iron | mg L−1 | 0.1–40 | [39,40,43,45,46] | |
| Lithium | mg L−1 | 0.1–507 | [39,40,46] | |
| Magnesium | mg L−1 | 2–700 | [39,40,43,45,46] | |
| Manganese | mg L−1 | 0.1–5.8 | [39,40,43,45] | |
| Potassium | mg L−1 | 170–390 | [39,44,46] | |
| Sodium | mg L−1 | 850–20,000 | [39,43,46] | |
| Heavy metals | Chromium | mg L−1 | 0.1–0.2 | [39,45] |
| Copper | mg L−1 | 0.06–4.2 | [39,40] | |
| Lead | mg L−1 | ~0.03 | [39] | |
| Nickel | mg L−1 | 0.06–0.2 | [39,45] | |
| Strontium | mg L−1 | 360–470 | [39] | |
| Zinc | mg L−1 | 0.003–1 | [39,40,45] |
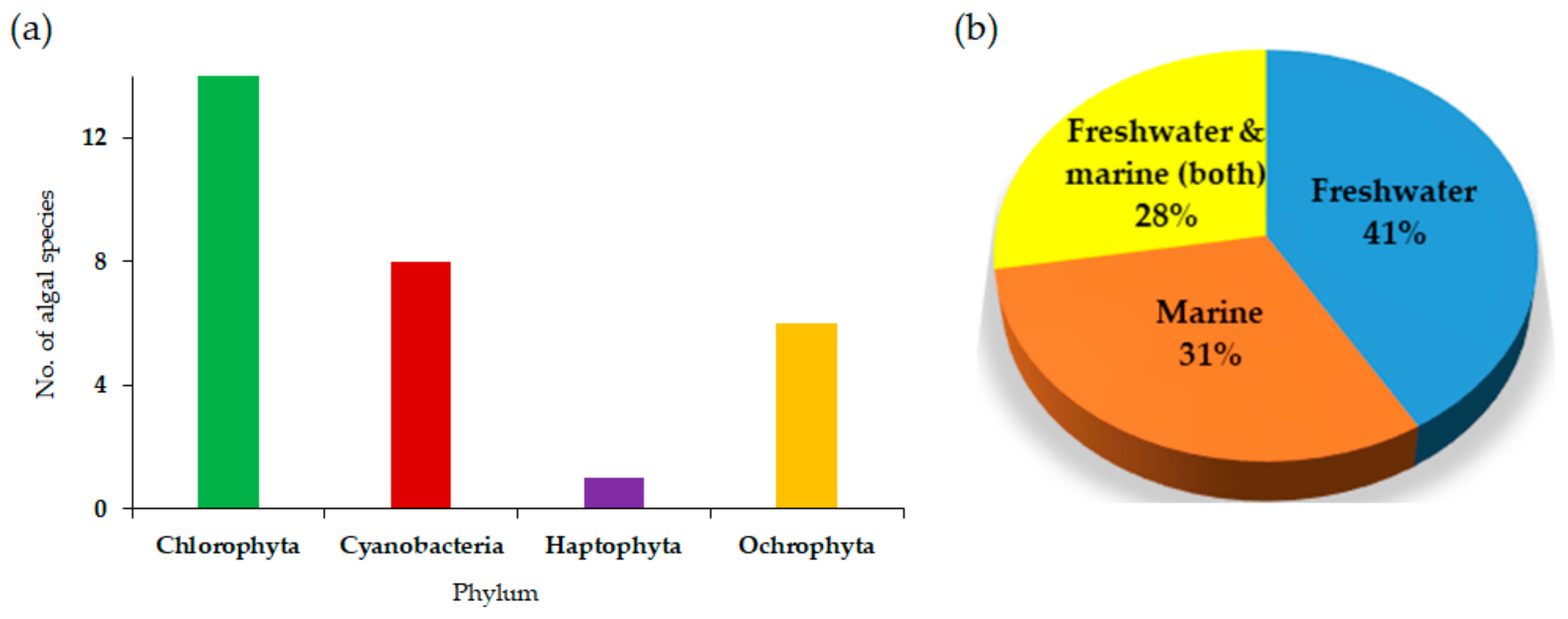
| Phylum | Name | Type of Algae | Assessment | PW Loading | Medium | Cultivation Type | Potential for Bioremediation | Ref | |
|---|---|---|---|---|---|---|---|---|---|
| Biomass Productivity | Bioremediation Potential | ||||||||
| Chlorophyta | Chlamydomonas reinhardtii | freshwater | ✓ | 0%, 20%, 40%, 60%, 80%, 100% | TAP | [50] | |||
| Chlorella sp. | freshwater | ✓ | ✓ | 0%, 25%, 50%, 100% PW loading [61] | H2 medium [62], BG-11 [4], f/2 [61], Broad seawater medium [47] | 1 L Photobioreactor (PBR), 450 mL Tube reactors [48] | TOC, TN, Al, Zn, Fe | [4,11,47,61,62,63] | |
| Dictyosphaerium sp. | freshwater/marine | ✓ | ✓ | 50%, 60%, 75%, 100% | Al, Zn, Fe | [11,63] | |||
| Dunaliella salina | marine | ✓ | ✓ | artificial seawater [39], Seawater [46] | 125 mL conical flask | Zn, Ni | [39,46] | ||
| Dunaliella tertiolecta | marine | ✓ | f/2 | [60] | |||||
| Monoraphidium sp. | freshwater | ✓ | ✓ | 50%, 60%, 75%,100% [63] | H2 medium [62], BG-11 [4] | Al, Zn, Fe | [4,11,62,63] | ||
| Neochloris oleoabundans | freshwater/marine | ✓ | Broad seawater medium [47] | 450 mL Tube reactors | Al, Zn, Fe | [11,47] | |||
| Parachlorella kessleri | freshwater | ✓ | f/2 | [41] | |||||
| Picochlorum sp. | marine | ✓ | f/2 | [4] | |||||
| Pseudokirchneriella subcapitata | freshwater | ✓ | [64] | ||||||
| Scenedesmus sp. | freshwater | ✓ | ✓ | 50%, 60%, 75%, 100% [63] | Wuxal (W), BG-11 [4] | Open pond-600 L [49] | Al, Zn, Fe | [4,11,49,63] | |
| Scenedesmus rotundus | freshwater | ✓ | [51] | ||||||
| Tetracystis sp. | freshwater | ✓ | [32] | ||||||
| Tetraselmis sp. | marine | ✓ | f/2 | [4] | |||||
| Ochrophyta | Amphora coffeaformis | freshwater/marine | ✓ | ✓ | H2 medium [62], Broad seawater medium [47] | 450 mL Tube reactors [48] | Ba, Ca, Fe, Si, Mn | [47,62] | |
| Chaetoceros gracilis | marine | ✓ | Broad seawater medium [47] | 450 mL Tube reactors | Ba, Ca, Fe, Si, Mn | [47] | |||
| Chaetoceros muelleri | marine | ✓ | Broad seawater medium [47] | 450 mL Tube reactors | [47] | ||||
| Nannochloropsis oculata | marine | ✓ | ✓ | 10–50% [45], 0%, 50%, 100% [42] | BG11 [45], f/2 [42] | Chemical oxygen demand (COD) and Oil content | [42,45] | ||
| Nannochloropsis salina | freshwater/marine | ✓ | f/2-Si (minus silica) | [32] | |||||
| Phaeodactylum tricornutum | freshwater/marine | ✓ | Broad seawater medium [47] | 450 mL Tube reactors | Ba, Ca, Fe, Si, Mn | [47] | |||
| Cyanobacteria | Cyanobacterium aponinum | freshwater/marine | ✓ | f/2 | [41] | ||||
| Cyanobium sp. | freshwater | ✓ | ✓ | H2 medium | phosphate | [62] | |||
| Logan Lagoons Cyanobacteria 2 (LLC2) | freshwater/marine | ✓ | [65] | ||||||
| Lyptolyngbya sp. | marine | ✓ | f/2 | [4] | |||||
| Phormidium sp. | freshwater | ✓ | ✓ | H2 medium | ammonia, phosphate | [62] | |||
| Pseudoanabaena sp. | freshwater | ✓ | H2 medium | [62] | |||||
| Synechocystis sp. PCC 6803 | freshwater | ✓ | Broad seawater medium [47] | 450 mL Tube reactors | [47] | ||||
| Synechococcus elongatus | freshwater | ✓ | 0%, 20%, 40%, 60%, 80%, 100% | Broad seawater medium [47] | 450 mL Tube reactors | [47] | |||
| Haptophyta | Isochrysis galbana | marine | ✓ | ✓ | 0%, 25%, 50%, 100% PW loading [61] | BG-11 | COD and Oil content | [45] | |
| Species (In Alphabetical Order) | Phylum | Salinity Range (‰) | Ref |
|---|---|---|---|
| Aphanothece sp. | Cyanobacteria | 0–175 | [117,118] |
| Asteroplanus karianus | Bacillariophyta | 10–35 | [119,120] |
| Chaetoceros sp. (e.g., Chaetoceros didymus, Chaetoceros elmorei) | Bacillariophyta | 10–35 | [119,121,122] |
| Cyanobacterium aponinum | Cyanobacteria | 15–60 | [41] |
| Cyclotella sp. (e.g., Cyclotella cryptica, Cyclotella marina, Cyclotella quillensis) | Bacillariophyta | 3–90 | [123,124,125] |
| Cymbella pusilla | Bacillariophyta | 3–200 | [122,126] |
| Ditylum brightwellii | Heterokonta | 10–35 | [119,127] |
| Dunaliella sp. (e.g., Dunaliella tertiolecta, Dunaliella salina) | Chlorophyta | 5–360 | [39,60] |
| Eutreptiella gymnestica | Euglenozoa | 5–30 | [128,129,130] |
| Nannochloropsis sp. (e.g., Nannochloropsis oculata) | Ochrophyta | 40 | [42,57,60] |
| Nitzschia longissima | Bacillariophyta | 10–35 | [119,131] |
| Oltmannsiellopsis viridis | Chlorophyta | 5–30 | [128] |
| Parachlorella kessleri | Chlorophyta | 15–60 | [41] |
| Picochlorum sp. (e.g., Picochlorum oklahomensis) | Chlorophyta | 35–150 | [132,133,134,135] |
| Picocystis salinarum | Chlorophyta | 15–300 | [136,137,138] |
| Prymnesium parvum | Haptophyta | 3–30 | [128,139,140] |
| Skeletonema sp. (e.g., Skeletonema costatum, Skeletonema subsalsum) | Bacillariophyta | 0–35 | [119,141] |
| Thalassionema nitzschioides | Bacillariophyta | 10–35 | [119,142,143] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, A.; Agrawal, S.; Nawaz, T.; Pan, S.; Selvaratnam, T. A Review of Algae-Based Produced Water Treatment for Biomass and Biofuel Production. Water 2020, 12, 2351. https://doi.org/10.3390/w12092351
Rahman A, Agrawal S, Nawaz T, Pan S, Selvaratnam T. A Review of Algae-Based Produced Water Treatment for Biomass and Biofuel Production. Water. 2020; 12(9):2351. https://doi.org/10.3390/w12092351
Chicago/Turabian StyleRahman, Ashiqur, Saumya Agrawal, Tabish Nawaz, Shanglei Pan, and Thinesh Selvaratnam. 2020. "A Review of Algae-Based Produced Water Treatment for Biomass and Biofuel Production" Water 12, no. 9: 2351. https://doi.org/10.3390/w12092351
APA StyleRahman, A., Agrawal, S., Nawaz, T., Pan, S., & Selvaratnam, T. (2020). A Review of Algae-Based Produced Water Treatment for Biomass and Biofuel Production. Water, 12(9), 2351. https://doi.org/10.3390/w12092351