
Epiphytic Diatom-Based Biomonitoring in Mediterranean Ponds: Traditional Microscopy versus Metabarcoding Approaches

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
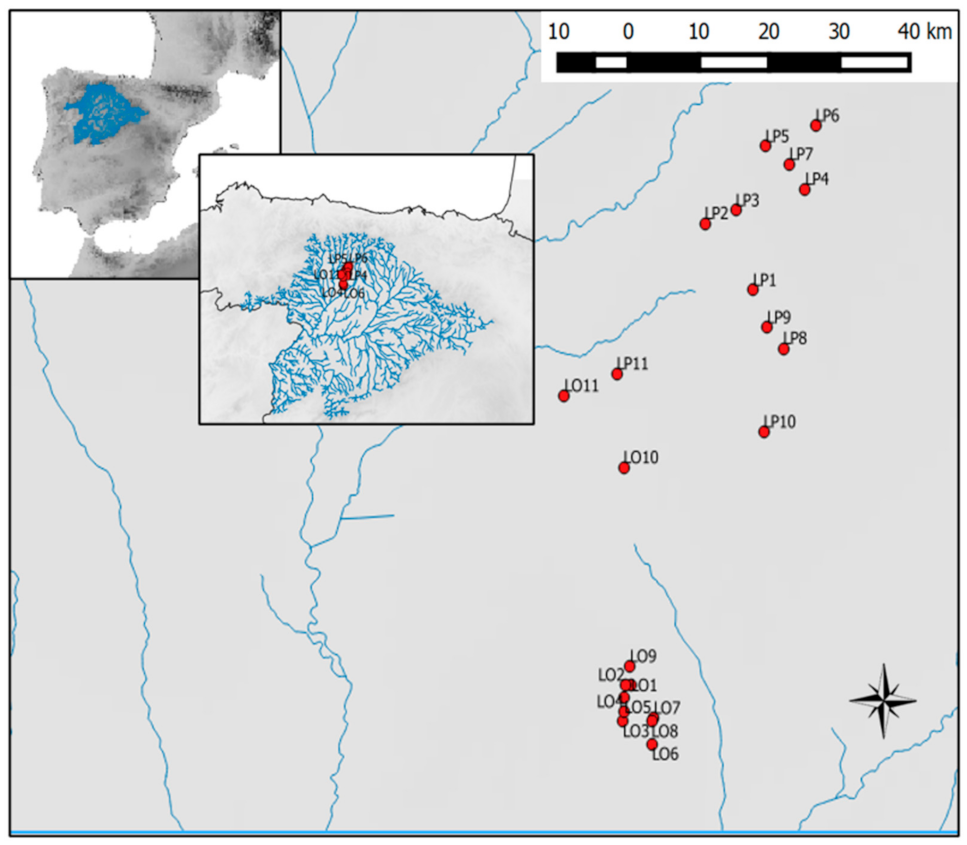
2.1. Study Area
2.2. Diatom Sampling
2.3. Morphological Analysis
2.4. DNA Extraction, PCR Amplification and Sequencing Data Processing
2.5. Bioinformatic Analysis
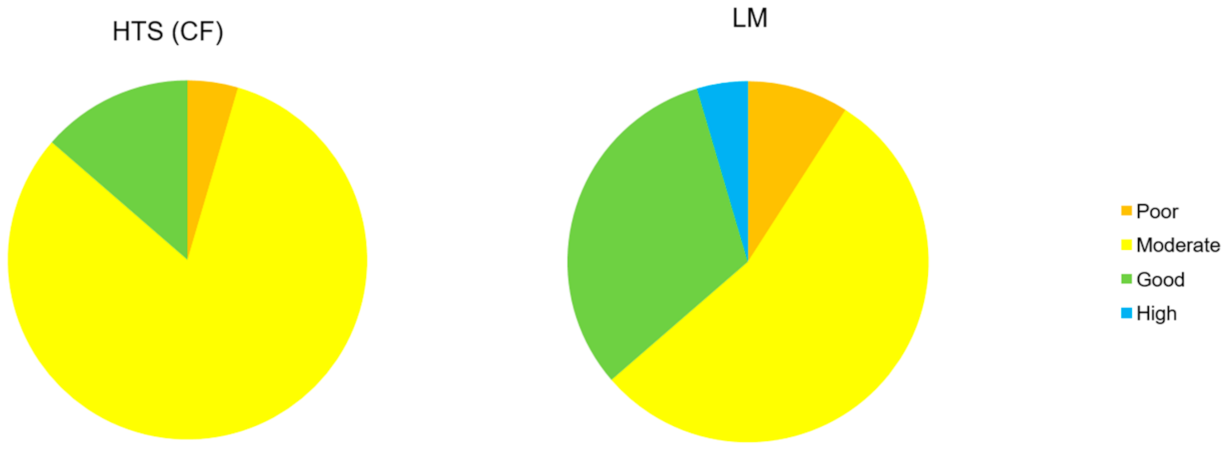
2.6. Ecological Status Class Assignment
3. Results
3.1. Morphological Analysis
3.2. HTS Analysis
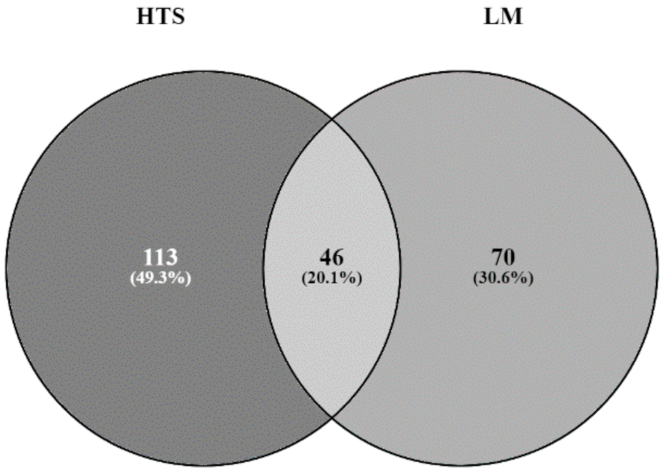
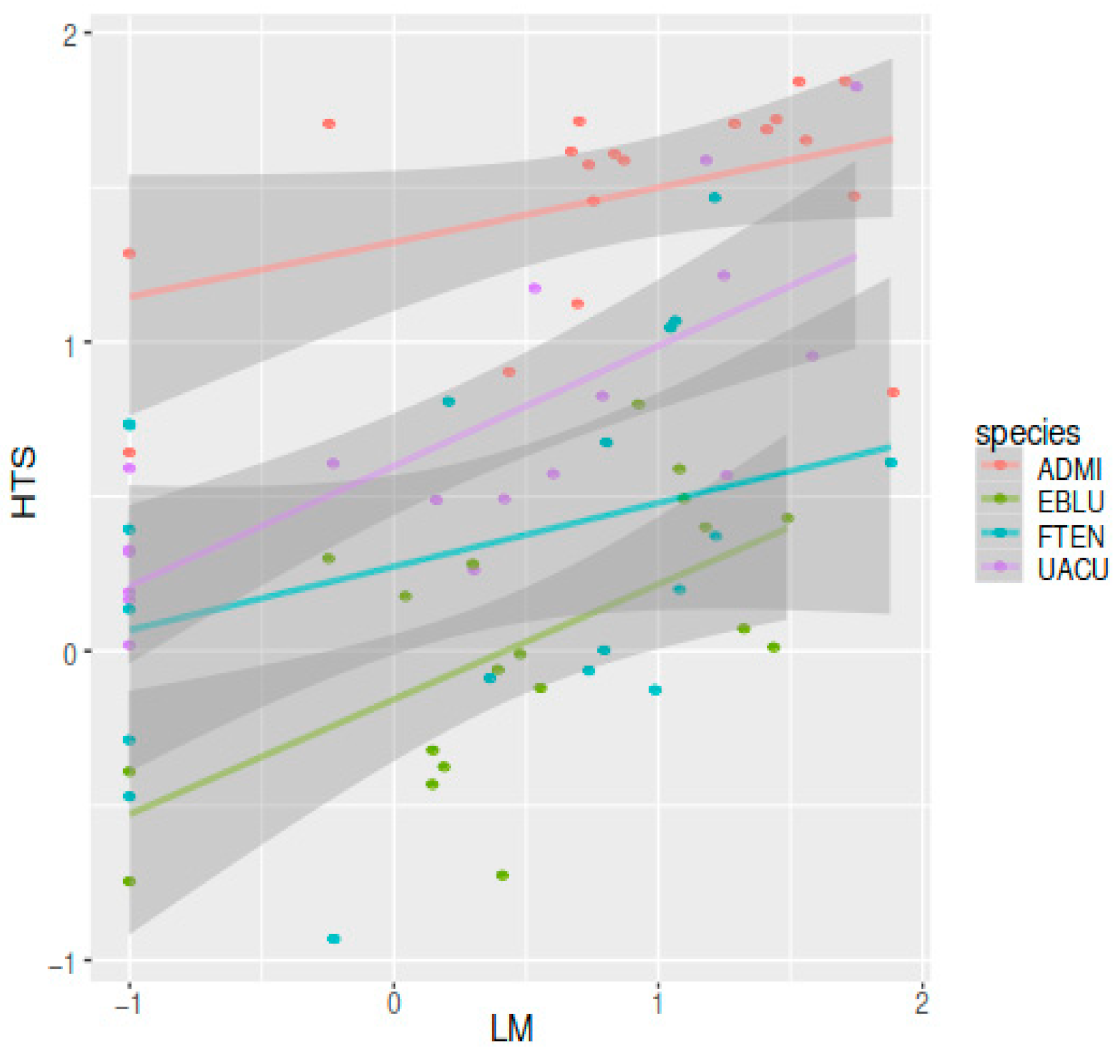
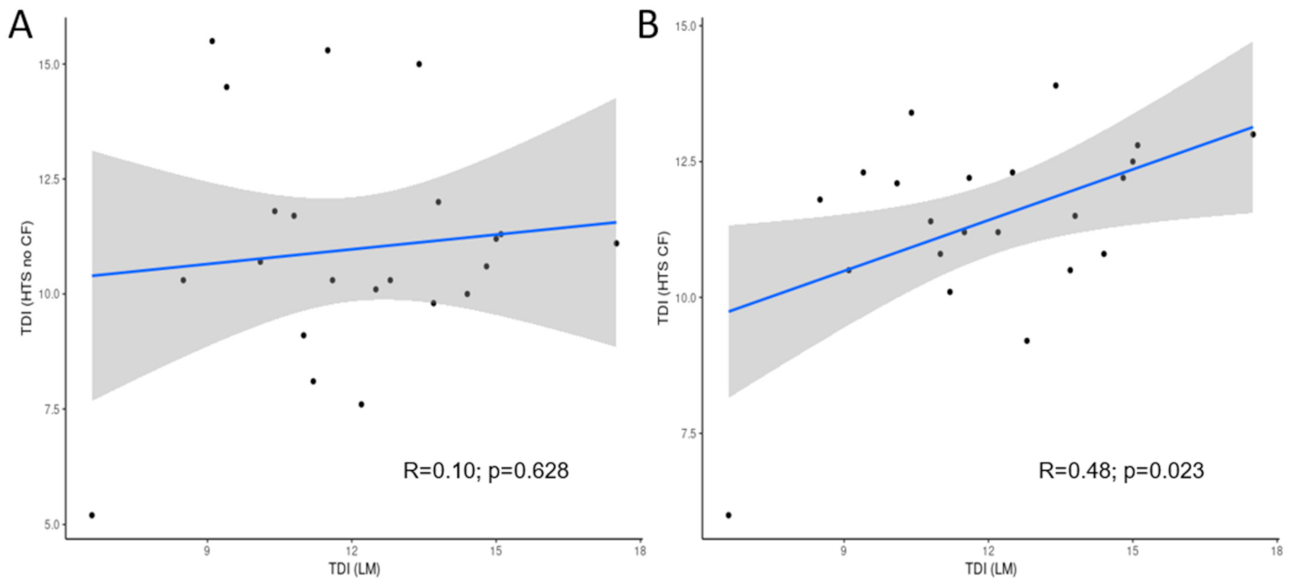
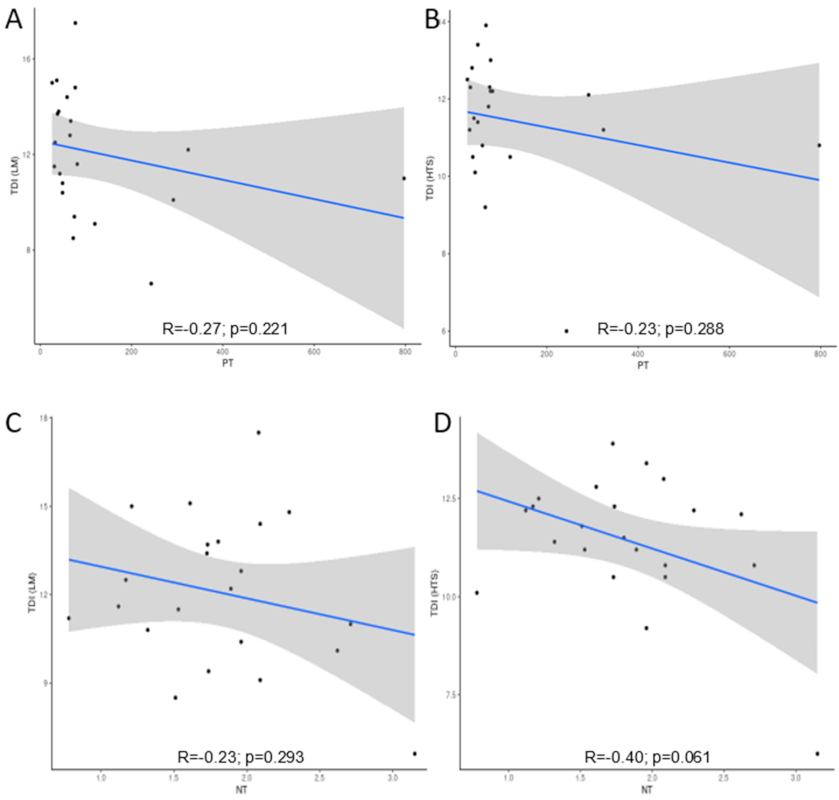
3.3. Comparison of LM and HTS Inventories
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- BARBE J; DES CLERCS S; PHILIPPE M., Etude Des Méthodes Biologiques d’appréciation Quantitative de La Qualité Des Eaux. Available online: https://www.oieau.org/eaudoc/notice/ETUDE-DES-METHODES-BIOLOGIQUES-DAPPRECIATION-QUANTITATIVE-DE-LA-QUALITE-DES-EAUX (accessed on 11 May 2021).
- Kelly, M.G.; Whitton, B.A. The Trophic Diatom Index: A New Index for Monitoring Eutrophication in Rivers. J. Appl. Phycol. 1995, 7, 433–444. [Google Scholar] [CrossRef]
- Kermarrec, L.; Franc, A.; Rimet, F.; Chaumeil, P.; Humbert, J.-F.; Bouchez, A. Next-Generation Sequencing to Inventory Taxonomic Diversity in Eukaryotic Communities: A Test for Freshwater Diatoms. Mol. Ecol. Resour. 2013, 13, 607–619. [Google Scholar] [CrossRef]
- Zimmermann, J.; Glöckner, G.; Jahn, R.; Enke, N.; Gemeinholzer, B. Metabarcoding vs. Morphological Identification to Assess Diatom Diversity in Environmental Studies. Mol. Ecol. Resour. 2015, 15, 526–542. [Google Scholar] [CrossRef]
- Rimet, F.; Abarca, N.; Bouchez, A.; Kusber, W.-H.; Jahn, R.; Kahlert, M.; Keck, F.; Kelly, M.G.; Mann, D.G.; Piuz, A.; et al. The Potential of High-Throughput Sequencing (HTS) of Natural Samples as a Source of Primary Taxonomic Information for Reference Libraries of Diatom Barcodes. Fottea 2018, 18, 37–54. [Google Scholar] [CrossRef] [Green Version]
- An, S.M.; Choi, D.H.; Lee, J.H.; Lee, H.; Noh, J.H. Identification of Benthic Diatoms Isolated from the Eastern Tidal Flats of the Yellow Sea: Comparison between Morphological and Molecular Approaches. PLoS ONE 2017, 12, e0179422. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Zhao, Y.; Sun, Y.; Li, Y.; Wu, P.; Zhou, S.; Ren, L. Comparative Study on Diatom Morphology and Molecular Identification in Drowning Cases. Forensic Sci. Int. 2020, 317, 110552. [Google Scholar] [CrossRef] [PubMed]
- Vasselon, V.; Rimet, F.; Domaizon, I.; Monnier, O.; Reyjol, R.; Bouchez, A. Assessing Pollution of Aquatic Environments with Diatoms’ DNA Metabarcoding: Experience and Developments from France Water Framework Directive Networks. Metabarcod. Metagenom. 2019, 3, e39646. [Google Scholar]
- Rivera, S.F.; Vasselon, V.; Bouchez, A.; Rimet, F. Diatom Metabarcoding Applied to Large Scale Monitoring Networks: Optimization of Bioinformatics Strategies Using Mothur Software. Ecol. Indic. 2020, 109, 105775. [Google Scholar] [CrossRef]
- Kelly, M.G.; Juggins, S.; Mann, D.G.; Sato, S.; Glover, R.; Boonham, N.; Sapp, M.; Lewis, E.; Hany, U.; Kille, P.; et al. Development of a Novel Metric for Evaluating Diatom Assemblages in Rivers Using DNA Metabarcoding. Ecol. Indic. 2020, 118, 106725. [Google Scholar] [CrossRef]
- Pérez-Burillo, J.; Trobajo, R.; Vasselon, V.; Rimet, F.; Bouchez, A.; Mann, D.G. Evaluation and Sensitivity Analysis of Diatom DNA Metabarcoding for WFD Bioassessment of Mediterranean Rivers. Sci. Total Environ. 2020, 727, 138445. [Google Scholar] [CrossRef]
- Masouras, A.; Karaouzas, I.; Dimitriou, E.; Tsirtsis, G.; Smeti, E. Benthic Diatoms in River Biomonitoring—Present and Future Perspectives within the Water Framework Directive. Water 2021, 13, 478. [Google Scholar] [CrossRef]
- Feio, M.J.; Serra, S.R.Q.; Mortágua, A.; Bouchez, A.; Rimet, F.; Vasselon, V.; Almeida, S.F.P. A Taxonomy-Free Approach Based on Machine Learning to Assess the Quality of Rivers with Diatoms. Sci. Total Environ. 2020, 722, 137900. [Google Scholar] [CrossRef]
- Visco, J.A.; Apothéloz-Perret-Gentil, L.; Cordonier, A.; Esling, P.; Pillet, L.; Pawlowski, J. Environmental Monitoring: Inferring the Diatom Index from next-Generation Sequencing Data. Environ. Sci. Technol. 2015, 49, 7597–7605. [Google Scholar] [CrossRef]
- Mortágua, A.; Vasselon, V.; Oliveira, R.; Elias, C.; Chardon, C.; Bouchez, A.; Rimet, F.; Feio, M.J.; Almeida, S.F. Applicability of DNA Metabarcoding Approach in the Bioassessment of Portuguese Rivers Using Diatoms. Ecol. Indic. 2019, 106, 105470. [Google Scholar] [CrossRef]
- Rivera, S.F.; Vasselon, V.; Jacquet, S.; Bouchez, A.; Ariztegui, D.; Rimet, F. Metabarcoding of Lake Benthic Diatoms: From Structure Assemblages to Ecological Assessment. Hydrobiologia 2018, 807, 37–51. [Google Scholar] [CrossRef]
- Blanco, S.; Romo, S.; Villena, M.-J.; Martínez, S. Fish Communities and Food Web Interactions in Some Shallow Mediterranean Lakes. Hydrobiologia 2003, 506, 473–480. [Google Scholar] [CrossRef]
- Blanco, S.; Ector, L.; Bécares, E. Epiphytic Diatoms as Water Quality Indicators in Spanish Shallow Lakes. Vie Milieu 2004, 54, 71–80. [Google Scholar]
- Blanco, S.; Cejudo-Figueiras, C.; Álvarez-Blanco, I.; van Donk, E.; Gross, E.M.; Hansson, L.-A.; Irvine, K.; Jeppesen, E.; Kairesalo, T.; Moss, B.; et al. Epiphytic Diatoms along Environmental Gradients in Western European Shallow Lakes. CLEAN Soil Air Water 2014, 42, 229–235. [Google Scholar] [CrossRef]
- Cejudo-Figueiras, C.; Álvarez-Blanco, I.; Bécares, E.; Blanco, S.; Cejudo-Figueiras, C.; Álvarez-Blanco, I.; Bécares, E.; Blanco, S. Epiphytic Diatoms and Water Quality in Shallow Lakes: The Neutral Substrate Hypothesis Revisited. Mar. Freshw. Res. 2011, 61, 1457–1467. [Google Scholar] [CrossRef]
- Nistal, A.; García, P.; García-Girón, J.; Borrego-Ramos, M.; Blanco, S.; Bécares, E. DNA Metabarcoding and Morphological Methods Show Complementary Patterns in the Metacommunity Organization of Lentic Epiphytic Diatoms. In ARPHA Conference Abstracts; Pensoft Publishers: Sofia, Bulgaria, 2021. [Google Scholar]
- Poulíčková, A.; Duchoslav, M.; Dokulil, M. Littoral Diatom Assemblages as Bioindicators of Lake Trophic Status: A Case Study from Perialpine Lakes in Austria. Eur. J. Phycol. 2004, 39, 143–152. [Google Scholar] [CrossRef]
- Bennion, H.; Kelly, M.G.; Juggins, S.; Yallop, M.L.; Burgess, A.; Jamieson, J.; Krokowski, J. Assessment of Ecological Status in UK Lakes Using Benthic Diatoms. Freshw. Sci. 2014, 33, 639–654. [Google Scholar] [CrossRef] [Green Version]
- Cantonati, M.; Spitale, D. The Role of Environmental Variables in Structuring Epiphytic and Epilithic Diatom Assemblages in Springs and Streams of the Dolomiti Bellunesi National Park (South-Eastern Alps). Fundam. Appl. Limnol. 2009, 174, 117–133. [Google Scholar] [CrossRef]
- Wojtal, A.Z.; Sobczyk, Ł. The Influence of Substrates and Physicochemical Factors on the Composition of Diatom Assemblages in Karst Springs and Their Applicability in Water-Quality Assessment. Hydrobiologia 2012, 695, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Winter, J.G.; Duthie, H.C. Stream Epilithic, Epipelic and Epiphytic Diatoms: Habitat Fidelity and Use in Biomonitoring. Aquat. Ecol. 2000, 34, 345–353. [Google Scholar] [CrossRef]
- Bécares, E.; Blanco, S. Método de muestreo de diatomeas epífitas en laguna para la aplicación de la Directiva Marco del Agua. Tecnología del Agua. 2006, 26, 42–47. [Google Scholar]
- Borrego-Ramos, M.; Olenici, A.; Blanco, S. Are Dead Stems Suitable Substrata for Diatom-Based Monitoring in Mediterranean Shallow Ponds? Fundam. Appl. Limnol. 2019, 192, 215–224. [Google Scholar] [CrossRef]
- Riato, L.; Leira, M.; Della Bella, V.; Oberholster, P.J. Development of a Diatom-Based Multimetric Index for Acid Mine Drainage Impacted Depressional Wetlands. Sci. Total Environ. 2018, 612, 214–222. [Google Scholar] [CrossRef] [Green Version]
- Zimba, P.V.; Hopson, M.S. Quantification of Epiphyte Removal Efficiency from Submersed Aquatic Plants. Aquat. Bot. 1997, 58, 173–179. [Google Scholar] [CrossRef]
- American Public Health Association. Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 1989. [Google Scholar]
- European Committee for Standardization (CEN). EN 13946:2014. Water Quality—Guidance for the Routine Sampling and Preparation of Benthic Diatoms from Rivers and Lakes; European Committee for Standardization: Brussels, Belgium, 2014. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Süsswasserflora von Mitteleuropa. Band 2/3. Bacillariophyceae. 3. Teil: Centrales, Fragilariaceae, Eunoticeae; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; G. Fischer Verlag: Stuttgart, Germany, 1991. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Süsswasserflora von Mitteleuropa. Band 2/4. Bacillariophyceae. 4. Teil: Achnanthaceae Kritische Ergänzungen Zu Navicula (Lineolatae) Und Gomphonema; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; G. Fischer Verlag: Stuttgart, Germany, 1991. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Süsswasserflora von Mitteleuropa, Band 2/1: Bacillariophyceae 1. Teil: Naviculaceae; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; G. Fischer Verlag: Stuttgart, Germany, 1986. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Sübwasserflora von Mitteleuropa. Band 2/2. Bacillariophyceae. 2. Teil: Bacillariaceae, Epithemiaceae, Surirellaceae; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; G. Fischer Verlag: Stuttgart, Germany, 1986. [Google Scholar]
- Lange-Bertalot, H.; Hofmann, G.; Werum, M.; Cantonati, M. Freshwater Benthic Diatoms of Central Europe: Over 800 Common Species Used in Ecological Assessment; Koeltz Botanical Books: Oberreifenberg, Germany, 2017. [Google Scholar]
- Vasselon, V.; Rimet, F.; Tapolczai, K.; Bouchez, A. Assessing Ecological Status with Diatoms DNA Metabarcoding: Scaling-up on a WFD Monitoring Network (Mayotte Island, France). Ecol. Indic. 2017, 82, 1–12. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing Mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME Improves Sensitivity and Speed of Chimera Detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Rimet, F.; Gusev, E.; Kahlert, M.; Kelly, M.G.; Kulikovskiy, M.; Maltsev, Y.; Mann, D.G.; Pfannkuchen, M.; Trobajo, R.; Vasselon, V.; et al. Diat.Barcode, an Open-Access Curated Barcode Library for Diatoms. Sci. Rep. 2019, 9, 15116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasselon, V.; Bouchez, A.; Rimet, F.; Jacquet, S.; Trobajo, R.; Corniquel, M.; Tapolczai, K.; Domaizon, I. Avoiding Quantification Bias in Metabarcoding: Application of a Cell Biovolume Correction Factor in Diatom Molecular Biomonitoring. Methods Ecol. Evol. 2018, 9, 1060–1069. [Google Scholar] [CrossRef] [Green Version]
- Lecointe, C.; Coste, M.; Prygiel, J. “Omnidia”: Software for Taxonomy, Calculation of Diatom Indices and Inventories Management. Hydrobiologia 1993, 269, 509–513. [Google Scholar] [CrossRef]
- Pinseel, E.; Vanormelingen, P.; Hamilton, P.B.; Vyverman, W.; Van de Vijver, B.; Kopalova, K. Molecular and Morphological Characterization of the Achnanthidium Minutissimum Complex (Bacillariophyta) in Petuniabukta (Spitsbergen, High Arctic) Including the Description of A. Digitatum sp. nov. Eur. J. Phycol. 2017, 52, 264–280. [Google Scholar] [CrossRef]
- Antoniades, D.; Hamilton, P.B.; Douglas, M.S.V.; Smol, J.P. Volume 17: Diatoms of North America: The Freshwater Floras of Prince Patrick, Ellef Ringnes and Northern Ellesmere Islands from the Canadian Arctic Archipelago; Iconographia Diatomologica. A.R.G.Gantner Verlag KG: Ruggell, Liechtenstein, 2008. [Google Scholar]
- Pla-Rabés, S.; Hamilton, P.B.; Ballesteros, E.; Gavrilo, M.; Friedlander, A.M.; Sala, E. The Structure and Diversity of Freshwater Diatom Assemblages from Franz Josef Land Archipelago: A Northern Outpost for Freshwater Diatoms. PeerJ 2016, 4, e1705. [Google Scholar] [CrossRef] [Green Version]
- Kermarrec, L.; Bouchez, A.; Rimet, F.; Humbert, J.-F. First Evidence of the Existence of Semi-Cryptic Species and of a Phylogeographic Structure in the Gomphonema Parvulum (Kützing) Kützing Complex (Bacillariophyta). Protist 2013, 164, 686–705. [Google Scholar] [CrossRef]
- Abarca, N.; Jahn, R.; Zimmermann, J.; Enke, N. Does the Cosmopolitan Diatom Gomphonema Parvulum (Kützing) Kützing Have a Biogeography? PLoS ONE 2014, 9, e86885. [Google Scholar] [CrossRef]
- Keck, F.; Vasselon, V.; Rimet, F.; Bouchez, A.; Kahlert, M. Boosting DNA Metabarcoding for Biomonitoring with Phylogenetic Estimation of Operational Taxonomic Units’ Ecological Profiles. Mol. Ecol. Resour. 2018, 18, 1299–1309. [Google Scholar] [CrossRef]
- Mann, D.G.; Evans, K.M. 13 Molecular Genetics and the Neglected Art of Diatomics. Unravelling Algae Past Present Future Algal Syst. 2007, 231. [Google Scholar] [CrossRef]
- Trobajo, R.; Mann, D.G.; Clavero, E.; Evans, K.M.; Vanormelingen, P.; McGregor, R.C. The Use of Partial Cox1, RbcL and LSU RDNA Sequences for Phylogenetics and Species Identification within the Nitzschia Palea Species Complex (Bacillariophyceae). Eur. J. Phycol. 2010, 45, 413–425. [Google Scholar] [CrossRef] [Green Version]
- Kahlert, M.; Kelly, M.G.; Mann, D.G.; Rimet, F.; Sato, S.; Bouchez, A.; Keck, F. Connecting the Morphological and Molecular Species Concepts to Facilitate Species Identification within the Genus Fragilaria (Bacillariophyta). J. Phycol. 2019, 55, 948–970. [Google Scholar] [CrossRef] [PubMed]
- Poulíčková, A.; Veselá, J.; Neustupa, J.; Škaloud, P. Pseudocryptic Diversity versus Cosmopolitanism in Diatoms: A Case Study on Navicula Cryptocephala Kütz. (Bacillariophyceae) and Morphologically Similar Taxa. Protist 2010, 161, 353–369. [Google Scholar] [CrossRef] [PubMed]
- Bailet, B.; Apothéloz-Perret-Gentil, L.; Baričević, A.; Chonova, T.; Franc, A.; Frigerio, J.-M.; Kelly, M.; Mora, D.; Pfannkuchen, M.; Proft, S.; et al. Diatom DNA Metabarcoding for Ecological Assessment: Comparison among Bioinformatics Pipelines Used in Six European Countries Reveals the Need for Standardization. Sci. Total Environ. 2020, 745, 140948. [Google Scholar] [CrossRef]
- Zgrundo, A.; Lemke, P.; Pniewski, F.; Cox, E.J.; Latała, A. Morphological and Molecular Phylogenetic Studies on Fistulifera Saprophila. Diatom Res. 2013, 28, 431–443. [Google Scholar] [CrossRef]
- Moss, B. Water Pollution by Agriculture. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 659–666. [Google Scholar] [CrossRef] [Green Version]
- Moss, B.; Jeppesen, E.; Søndergaard, M.; Lauridsen, T.L.; Liu, Z. Nitrogen, Macrophytes, Shallow Lakes and Nutrient Limitation: Resolution of a Current Controversy? Hydrobiologia 2013, 710, 3–21. [Google Scholar] [CrossRef]
- Alba Tercedor, J.; Jáimez-Cuéllar, P.; Álvarez, M.; Avilés, J.; Bonada i Caparrós, N.; Casas, J.; Mellado, A.; Ortega, M.; Pardo, I.; Prat i Fornells, N.; et al. Caracterización Del Estado Ecológico de Ríos Mediterráneos Ibéricos Mediante El Índice IBMWP (Antes BMWP’). Limnetica 2002, 21, 175–185. [Google Scholar]
- Tapolczai, K.; Keck, F.; Bouchez, A.; Rimet, F.; Kahlert, M.; Vasselon, V. Diatom DNA Metabarcoding for Biomonitoring: Strategies to Avoid Major Taxonomical and Bioinformatical Biases Limiting Molecular Indices Capacities. Front. Ecol. Evol. 2019, 7, 409. [Google Scholar] [CrossRef] [Green Version]
- Apothéloz-Perret-Gentil, L.; Bouchez, A.; Cordier, T.; Cordonier, A.; Guéguen, J.; Rimet, F.; Vasselon, V.; Pawlowski, J. Monitoring the Ecological Status of Rivers with Diatom EDNA Metabarcoding: A Comparison of Taxonomic Markers and Analytical Approaches for the Inference of a Molecular Diatom Index. Mol. Ecol. 2020. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pond | Code | UTM Zone | UTM N | UTM E | Area (Ha) | Elevation (m ASL) |
|---|---|---|---|---|---|---|
| Linos | LO1 | 30T | 4,685,289.29 | 309,108.76 | 0.69 | 843 |
| Grande | LO2 | 30T | 4,685,301.76 | 308,728.38 | 3.79 | 840 |
| Cifuentes 2 | LO3 | 30T | 4,683,329.74 | 308,606.01 | 0.44 | 841 |
| Cifuentes 1 | LO4 | 30T | 4,683,839.42 | 308,643.42 | 0.69 | 841 |
| La Raya | LO5 | 30T | 4,684,646.4 | 308,709.17 | 0.36 | 842 |
| Amor | LO6 | 30T | 4,682,078.48 | 310,393.97 | 1.21 | 829 |
| Valdecastillo 1 | LO7 | 30T | 4,683,488.1 | 310,502.12 | 0.78 | 829 |
| Valdecastillo 2 | LO8 | 30T | 4,683,321.89 | 310,363.43 | 0.44 | 829 |
| Adobes | LO9 | 30T | 4,686,327.06 | 309,039.06 | 2.14 | 845 |
| Seca | LO10 | 30T | 4,697,167.00 | 308,657.00 | 3.78 | 831 |
| SantasMartas | LO11 | 30T | 4,701,107.00 | 305,033.00 | 0.03 | 834 |
| Mayor | LP1 | 30T | 4,706,991.32 | 316,586.61 | 3.78 | 898 |
| Diel | LP2 | 30T | 4,710,657.68 | 313,622.79 | 1.75 | 889 |
| Cantos | LP3 | 30T | 4,711,368.47 | 315,523.73 | 1.56 | 907 |
| Trigo | LP4 | 30T | 4,712,540.36 | 319,786.92 | 1.10 | 930 |
| Gente | LP5 | 30T | 4,714,890.29 | 317,344.56 | 0.96 | 922 |
| Villaverde | LP6 | 30T | 4,716,025.62 | 320,422.88 | 0.76 | 946 |
| Sentiz | LP7 | 30T | 4,713,904.81 | 318,812.00 | 4.79 | 922 |
| Estorrubio | LP8 | 30T | 4,703,798.94 | 318,434.81 | 0.95 | 895 |
| Redos | LP9 | 30T | 4,704,938.79 | 317,450.21 | 0.95 | 895 |
| Manzana | LP10 | 30T | 4,699,206.44 | 317,237.03 | 0.95 | 879 |
| Carrillón | LP11 | 30T | 4,702,315.00 | 308,237.00 | 0.99 | 844 |
| SPECIES | LM | HTS | SPECIES | HTS | LM |
|---|---|---|---|---|---|
| ADMI | 19.66 | 41.98 | ADMI | 41.98 | 19.66 |
| ACHD | 10.96 | 0.26 | UACU | 10.73 | 8.68 |
| FTEN | 9.05 | 0.00 | FRAG | 7.54 | 0.13 |
| UACU | 8.68 | 10.73 | GSPP | 7.41 | 0.00 |
| EBLU | 7.59 | 1.46 | NPAL | 5.90 | 0.46 |
| GEXL | 7.24 | 0.00 | GAFF | 4.07 | 0.00 |
| NIGR | 6.82 | 0.01 | FNNO | 3.97 | 0.00 |
| NPAE | 6.16 | 0.49 | NIPU | 2.46 | 0.00 |
| NPAD | 5.04 | 0.00 | NACD | 2.24 | 0.02 |
| GPAR | 1.78 | 0.00 | FGRA | 1.50 | 0.00 |
| HTS | LM | ||||
| Poor | Moderate | Good | High | ||
| Poor | 1 | 0 | 0 | 0 | |
| Moderate | 1 | 11 | 6 | 0 | |
| Good | 0 | 1 | 1 | 1 | |
| High | 0 | 0 | 0 | 0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borrego-Ramos, M.; Bécares, E.; García, P.; Nistal, A.; Blanco, S. Epiphytic Diatom-Based Biomonitoring in Mediterranean Ponds: Traditional Microscopy versus Metabarcoding Approaches. Water 2021, 13, 1351. https://doi.org/10.3390/w13101351
Borrego-Ramos M, Bécares E, García P, Nistal A, Blanco S. Epiphytic Diatom-Based Biomonitoring in Mediterranean Ponds: Traditional Microscopy versus Metabarcoding Approaches. Water. 2021; 13(10):1351. https://doi.org/10.3390/w13101351
Chicago/Turabian StyleBorrego-Ramos, María, Eloy Bécares, Pedro García, Alejandro Nistal, and Saúl Blanco. 2021. "Epiphytic Diatom-Based Biomonitoring in Mediterranean Ponds: Traditional Microscopy versus Metabarcoding Approaches" Water 13, no. 10: 1351. https://doi.org/10.3390/w13101351
APA StyleBorrego-Ramos, M., Bécares, E., García, P., Nistal, A., & Blanco, S. (2021). Epiphytic Diatom-Based Biomonitoring in Mediterranean Ponds: Traditional Microscopy versus Metabarcoding Approaches. Water, 13(10), 1351. https://doi.org/10.3390/w13101351