
Cryptic Constituents: The Paradox of High Flux–Low Concentration Components of Aquatic Ecosystems

 ,
,
Abstract
:1. Introduction
2. Carbon Cycling in Aquatic Environments
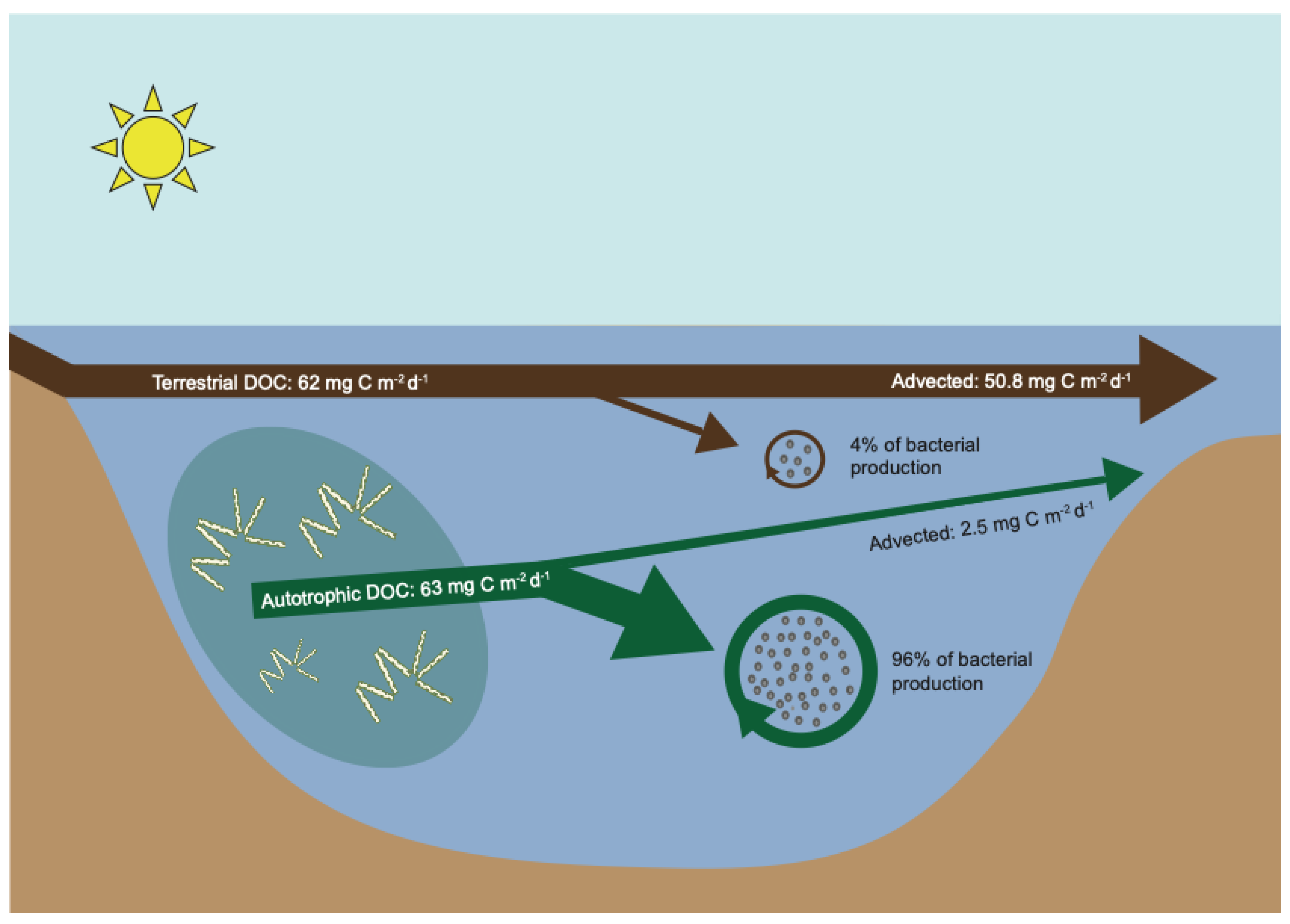
2.1. Dissolved Organic Carbon Metabolism
2.2. Cryptic Grazing—Pelagic Ecosystems
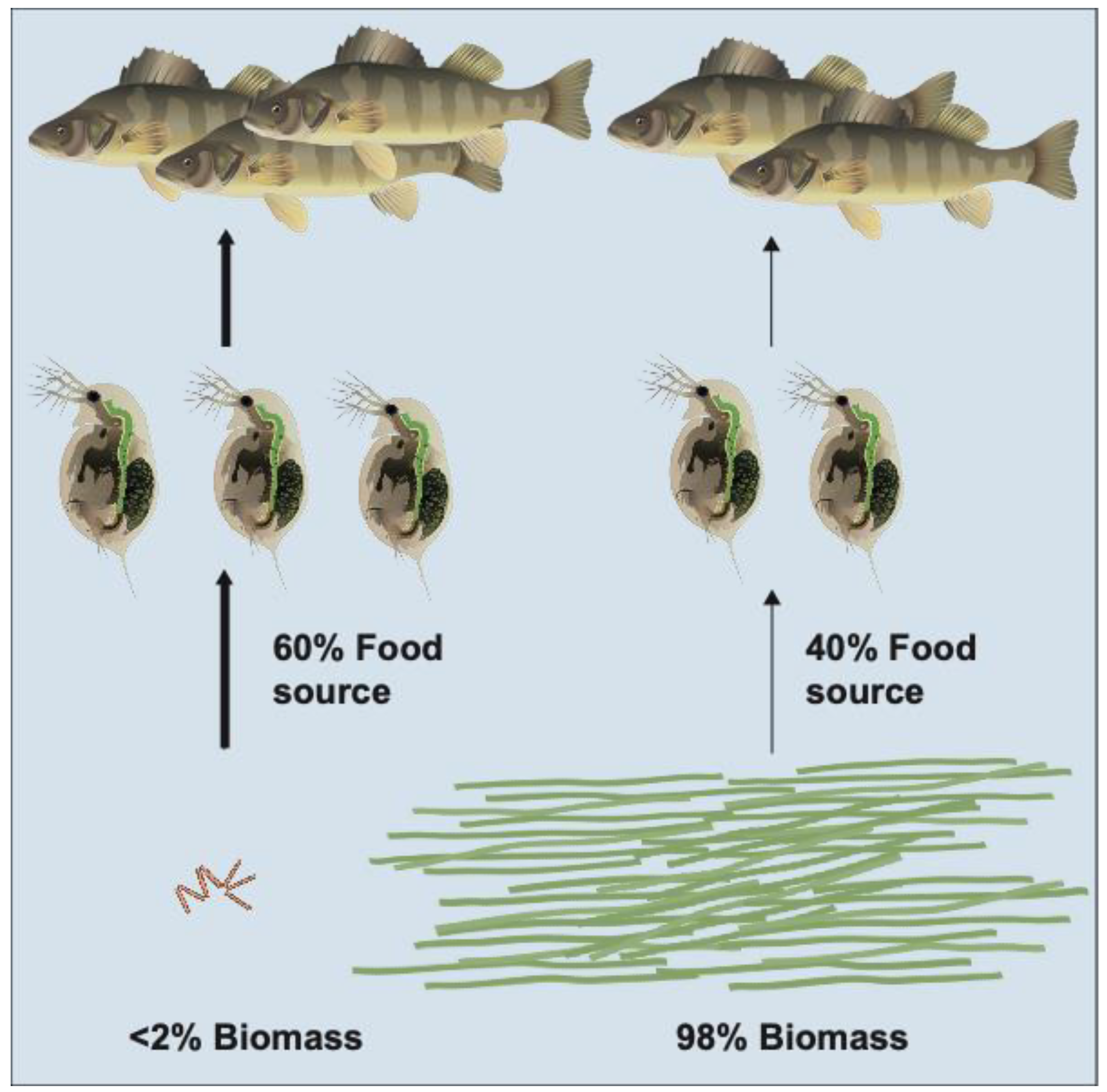
2.3. Cryptic Grazing in Benthic Food Webs
3. Nitrogen Limitation
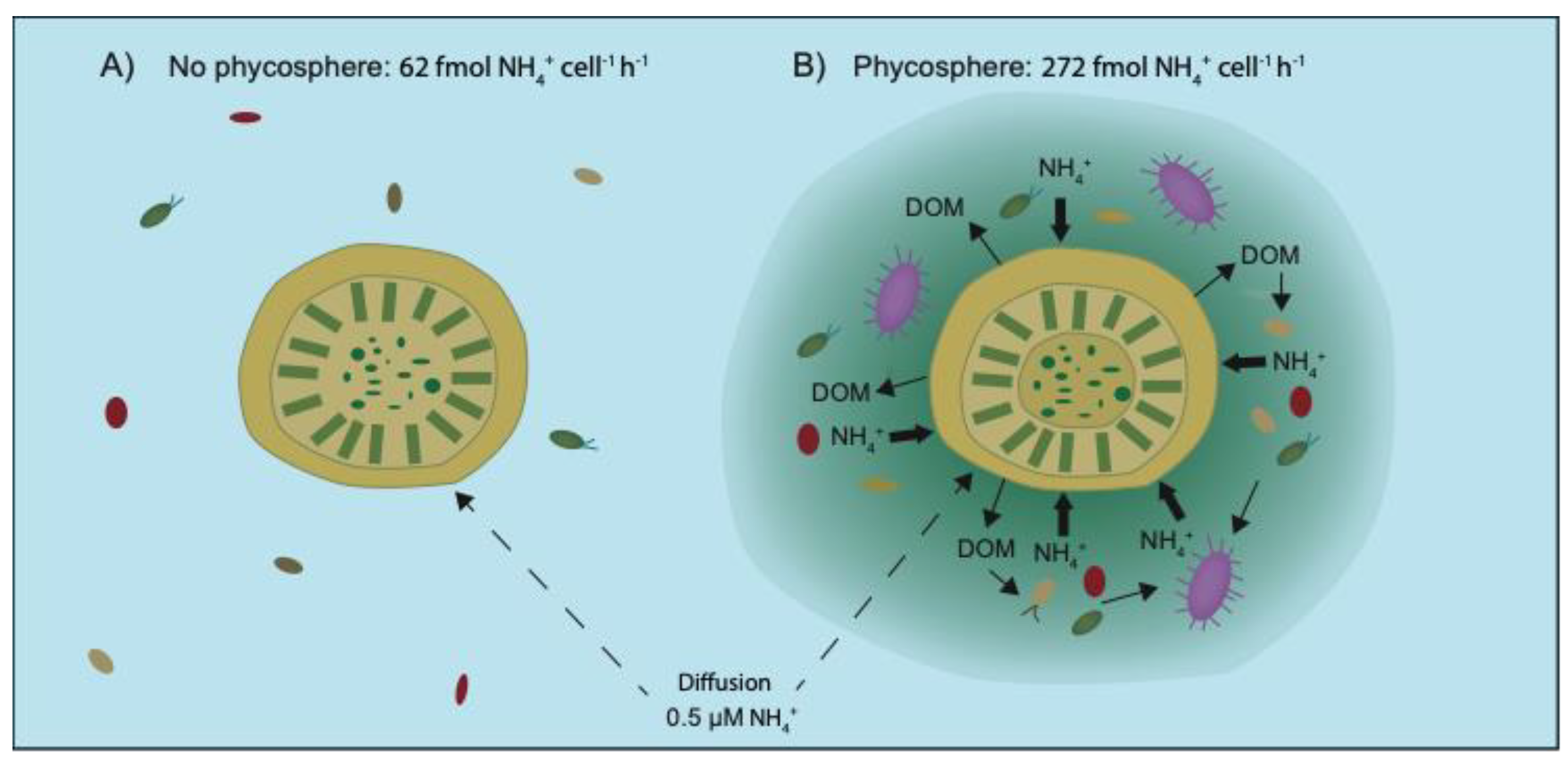
3.1. Ammonium Utilization
3.2. Nitrite Transformation
4. Conclusions and Future Implications
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [Green Version]
- Williams, P.J.I. Heterotrophic bacteria and the dynamics of dissolved organic material. In Microbial Ecology of the Oceans; Kirchman, D.L., Ed.; Wiley-Liss: New York, NY, USA, 2000; pp. 153–200. [Google Scholar]
- Karlson, A.; Duberg, J.; Motwani, N.H.; Hogfors, H.; Klawonn, I.; Ploug, H.; Barthel Svedén, J.; Garbaras, A.; Sundelin, B.; Hajdu, S.; et al. Nitrogen fixation by cyanobacteria stimulates production in Baltic food webs. AMBIO 2015, 44, S413–S426. [Google Scholar] [CrossRef] [Green Version]
- Hanson, P.C.; Hamilton, D.P.; Stanley, E.H.; Preston, N.; Langman, O.C.; Kara, E.L. Fate of allochthonous dissolved organic carbon in lakes: A quantitative approach. PLoS ONE 2011, 6, e21884. [Google Scholar] [CrossRef] [Green Version]
- Brett, M.T.; Bunn, S.E.; Chandra, S.; Galloway, A.W.E.; Guo, F.; Kainz, M.J.; Kankaala, P.; Lau, D.C.P.; Moulton, T.P.; Power, M.E.; et al. How important are terrestrial organic carbon inputs for secondary production in freshwater ecosystems? Freshw. Biol. 2017, 62, 833–853. [Google Scholar] [CrossRef]
- Creed, I.F.; Bergström, A.-K.; Trick, C.G.; Grimm, N.B.; Hessen, D.O.; Karlsson, J.; Kidd, K.A.; Kritzberg, E.; McKnight, D.M.; Freeman, E.C.; et al. Global change-driven effects on dissolved organic matter composition: Implications for food webs of northern lakes. Glob. Chang. Biol. 2018, 24, 3692–3714. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, R.G. Death, detritus and energy flow in aquatic ecosystems. Freshwat. Biol. 1995, 33, 83–89. [Google Scholar] [CrossRef]
- Prairie, Y.T. Carbocentric limnology: Looking back, looking forward. Can. J. Fish. Aquat. Sci. 2008, 65, 543–548. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, C.S. A changing paradigm of pelagic food webs. Int. Rev. Hydrobio. 2008, 93, 517–531. [Google Scholar] [CrossRef]
- Brett, M.T.; Arhonditsis, G.B.; Chandra, S.; Kainz, M.J. Mass flux calculations show strong allochthonous support of freshwater zooplankton production is unlikely. PLoS ONE 2012, 7, e39508. [Google Scholar] [CrossRef]
- Brett, M.T.; Benjamin, M.M. A reassessment of lake phosphorus retention and the nutrient loading concept in limnology. Freshw. Biol. 2008, 53, 194–211. [Google Scholar]
- Wetzel, R.G. Limnology, Lake and River Ecosystems, 3rd ed.; Academic Press: San Diego, CA, USA, 2001. [Google Scholar]
- Lewis, W.M., Jr. Global primary production of lakes: 19th Baldi Memorial Lecture. Inland Waters 2011, 1, 1–28. [Google Scholar] [CrossRef] [Green Version]
- von Wachenfeldt, E.; Tranvik, L.J. Sedimentation in Boreal Lakes—The Role of Flocculation of Allochthonous Dissolved Organic Matter in the Water Column. Ecosystems 2008, 11, 803–814. [Google Scholar] [CrossRef]
- del Giorgio, P.A.; Cole, J.J. Bacterial growth efficiency in natural aquatic systems. Annu. Rev. Ecol. Syst. 1998, 29, 503–541. [Google Scholar] [CrossRef] [Green Version]
- Olofsson, M.; Robertson, E.K.; Edler, L.; Arneborg, L.; Whitehouse, M.J.; Ploug, H. Nitrate and ammonium fluxes to diatoms and dinoflagellates at a single cell level in mixed field communities in the sea. Sci. Rep. 2019, 9, 1424. [Google Scholar] [CrossRef] [PubMed]
- Tiselius, P.; Belgrano, A.; Andersson, L.; Lindahl, O. Primary productivity in a coastal ecosystem: A trophic perspective on a long-term time series. J. Plankton Res. 2015, 38, 1092–1102. [Google Scholar] [CrossRef]
- Galloway, A.W.E.; Brett, M.T.; Holtgrieve, G.W.; Ward, E.J.; Ballantyne, A.P.; Burns, C.W.; Kainz, M.J.; Müller-Navarra, D.C.; Persson, J.; Ravet, J.L.; et al. A fatty acid based Bayesian approach for inferring diet in aquatic consumers. PLoS ONE 2015, 10, e0129723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brett, M.T.; Müeller-Navarra, D.C. The role of highly unsaturated fatty acids in aquatic food web processes. Freshw. Biol. 1997, 38, 483–499. [Google Scholar] [CrossRef]
- Klawonn, I.; Nahar, N.; Walve, J.; Andersson, B.; Olofsson, M.; Svedén, J.B.; Littman, S.; Whitehouse, M.J.; Kuypers, M.M.M.; Ploug, H. Cell-specific nitrogen- and carbon-fixation of cyanobacteria in a temperate marine system (Baltic Sea). Environ. Microbiol. 2016, 18, 4596–4609. [Google Scholar] [CrossRef]
- Stoecker, D.K.; Hansen, P.J.; Caron, D.A.; Mitra, A. Mixotrophy in the marine plankton. Annu. Rev. Mar. Sci. 2017, 9, 311–335. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, J.M.; Clare, E.; Hayden, B.; Brett, M.T.; Kratina, P. Diet tracing in ecology: Method comparison and selection. Meth. Ecol. Evol. 2017, 9, 278–291. [Google Scholar] [CrossRef]
- Glibert, P.M.; Middelburg, J.J.; McClelland, J.W.; Vander Zanden, M.J. Stable isotope tracers: Enriching our perspectives and questions on sources, fates, rates, and pathways of major elements in aquatic systems. Limnol. Oceanogr. 2018, 9999, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Hungate, B.A.; Mau, R.L.; Schwartz, E.; Caporaso, J.G.; Dijkstra, P.; van Gestel, N.; Koch, B.J.; Liu, C.M.; McHugh, T.A.; Marks, J.C.; et al. Quantitative Microbial Ecology through Stable Isotope Probing. Appl. Envir. Microbiol. 2015, 81, 7570–7581. [Google Scholar] [CrossRef] [Green Version]
- Vadeboncoeur, Y.; Power, M.E. Attached algae: The cryptic base of inverted trophic pyramids in freshwaters. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 255–279. [Google Scholar] [CrossRef]
- Power, M.E. Resource Enhancement by Indirect Effects of Grazers: Armored Catfish, Algae, and Sediment. Ecology 1990, 71, 897–904. [Google Scholar] [CrossRef]
- André, E.R.; Hecky, R.E.; Duthie, H.C. Nitrogen and phosphorus regeneration by cichlids in the littoral zone of Lake Malawi, Africa. J. Great Lakes Res. 2003, 29, 190–201. [Google Scholar] [CrossRef]
- Munubi, R.N.; McIntyre, P.B.; Vadeboncoeur, Y. Do grazers respond to or control food quality? Cross-scale analysis of algivorous fish in littoral Lake Tanganyika. Oecologia 2018, 188, 889–900. [Google Scholar] [CrossRef] [PubMed]
- Power, M.E. Habitat Quality and the Distribution of Algae-Grazing Catfish in a Panamanian Stream. J. Anim. Ecol. 1984, 53, 357–374. [Google Scholar] [CrossRef]
- Mittelbach, G.G.; Osenberg, C.W.; Leibold, M.A. Trophic relations and Ontogenetic niche shifts in Aquatic Ecosystems. In Size Structured Populations; Ebenman, B., Persson, L., Eds.; Springer: Berlin, Germany, 1988; pp. 1–17. [Google Scholar]
- Vander Zanden, M.J.; Chandra, S.; Park, S.-K.; Vadeboncoeur, Y.; Goldman, C.R. The relative efficiencies of benthic and pelagic trophic pathways in a subalpine lake. Can. J. Fish. Aquat. Sci. 2006, 63, 2608–2620. [Google Scholar] [CrossRef]
- Vadeboncoeur, Y.; McCann, K.; Zanden, M.; Rasmussen, J. Effects of multi-chain omnivory on the strength of trophic control in lakes. Ecosystems 2005, 8, 682–693. [Google Scholar] [CrossRef] [Green Version]
- James, M.R.; Hawes, I.; Weatherhead, M.; Carmen Stanger, C.; Gibbs, M. Carbon flow in the littoral food web of an oligotrophic lake. Hydrobiologia 2000, 441, 93–106. [Google Scholar] [CrossRef]
- Moore, J.W. Some factors effecting algal consumption in subarctic Ephemeroptera, Plecoptera and Simuliidae. Oecologia 1977, 27, 261–273. [Google Scholar] [CrossRef]
- Scharnvaeber, K.; Syväranta, J.; Hilt, S.; Brauns, M.; Vanni, M.J.; Brothers, S.; Köhler, J.; Knežević-Jarić, J.; Mehner, T. Whole-lake experiments reveal the fate of terrestrial particulate organic carbon in benthic food webs of shallow lakes. Ecology 2014, 95, 1496–1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, F.; Kainz, M.J.; Valdez, D.; Sheldon, F.; Bunn, S.E. High-quality algae attached to leaf litter boost invertebrate shredder growth. Freshw. Sci. 2016, 35, 1213–1221. [Google Scholar] [CrossRef] [Green Version]
- Cashman, M.J.; Harvey, G.L.; Wharton, G.; Bruno, M.C. Wood mitigates the effect of hydropeaking scour on periphyton biomass and nutritional quality in semi-natural flume simulations. Aquat. Sci. 2017, 79, 459–471. [Google Scholar] [CrossRef]
- Kupferberg, S.J.; Marks, J.C.; Power, M.E. Effects of variation in natural algal and detrital diets on larval anuran (Hyla regilla) life-history traits. Copeia 1994, 1994, 446–457. [Google Scholar] [CrossRef]
- Power, M.E.; Lowe, R.; Furey, P.; Welter, J.; Limm, M.; Finlay, J.; Bode, C.; Chang, S.; Goodrich, M.; Sculley, J. Algal mats and insect emergence in rivers under Mediterranean climates: Towards photogrammetric surveillance. Freshw. Biol. 2008, 54, 2101–2115. [Google Scholar] [CrossRef]
- Torres-Ruiz, M.; Wehr, J.D.; Perrone, A.A. Trophic relations in a stream food web: Importance of fatty acids for macroinvertebrate consumers. J. North. Am. Benthol. Soc. 2007, 26, 509–522. [Google Scholar] [CrossRef]
- Guo, F.; Kainz, M.J.; Sheldon, F.; Bunn, S.E. The importance of high-quality algal food sources in stream food webs-current status and future perspectives. Freshw. Biol. 2016, 61, 815–831. [Google Scholar] [CrossRef]
- Glibert, P.M.; Goldman, J. Rapid ammonium uptake by marine phytoplankton. Mar. Biol. Lett. 1981, 2, 25–31. [Google Scholar]
- Canfield, D.E.; Glazer, A.N.; Falkowski, P.G. The evolution and future of the nitrogen cycle. Science 2010, 330, 192–196. [Google Scholar] [CrossRef] [Green Version]
- Olofsson, M.; Karlberg, M.; Lage, S.; Ploug, H. Phytoplankton community composition and primary production in the tropical tidal ecosystem, Maputo Bay (the Indian Ocean). J. Sea Res. 2017, 125, 18–25. [Google Scholar] [CrossRef]
- Klawonn, I.; Bonaglia, S.; Whitehouse, M.J.; Littmann, S.; Tienken, D.; Kuypers, M.M.M.; Brüchert, V.; Ploug, H. Untangling hidden nutrient dynamics: Rapid ammonium cycling and single-cell ammonium assimilation in marine plankton communities. ISME J. 2019, 13, 1960–1974. [Google Scholar] [CrossRef]
- Musat, N.; Foster, R.; Vagner, T.; Adam, B.; Kuypers, M.M.M. Detecting metabolic activities in single cells, with emphasis on nanoSIMS. FEMS Microb. Rev. 2012, 36, 486–511. [Google Scholar] [CrossRef] [Green Version]
- Bergkvist, J.; Klawonn, I.; Whitehouse, M.J.; Lavik, G.; Brüchert, V.; Ploug, H. Turbulence simultaneously stimulates small- and large-scale CO2 sequestration by chain-forming diatoms in the sea. Nat. Comms. 2018, 9, 3046. [Google Scholar] [CrossRef] [PubMed]
- Clift, R.; Grace, J.R.; Weber, M.E. Bubbles, Drops, and Particles; Acad. Press: New York, NY, USA, 1978. [Google Scholar]
- Seymour, J.R.; Amin, S.A.; Raina, J.-B.; Stocker, R. Zooming in on the phycosphere: The ecological interface for phytoplankton-bacteria relationship. Nat. Microb. 2017, 2, 17065. [Google Scholar] [CrossRef] [PubMed]
- Glibert, P.M.; Wilkerson, F.P.; Dugdale, R.-C.; Raven, J.A.; Dupont, C.L.; Leavitt, P.R.; Parker, A.E.; Burkholder, J.M.; Kana, R.M. Pluses and minuses of ammonium and nitrate uptake and assimilation by phytoplankton and implications for productivity and community composition, with emphasis on nitrogen-enriched conditions. Limnol. Oceanogr. 2016, 61, 165–197. [Google Scholar] [CrossRef]
- Ward, B.B. Nitrification: An introduction and overview of the state of the field. In Nitrification; Ward, B.B., Arp, D.J., Klotz, M.G., Eds.; ASM Press: Washington, DC, USA, 2011; pp. 3–8. [Google Scholar]
- Jetten, M.S.M.; Strous, M.; van de Pas-Schoonen, K.T.; Schalk, J.; van Dongen, U.G.J.M.; can de Graaf, A.A.; Logemann, S.; Muyzer, G.; van Loosdrecht, M.C.M.; Kuenen, J.G. The anaerobic oxidation of ammonium. FEMS Microbiol. Rev. 1998, 22, 421–437. [Google Scholar] [CrossRef] [PubMed]
- Zakem, E.; Al-Haj, A.; Church, M.J.; van Dijken, G.L.; Dutkiewicz, S.; Foster, S.Q.; Fulweiler, R.W.; Mills, M.M.; Follows, M.J. Ecological control of nitrite in the upper ocean. Nat. Comms. 2018, 9, 1206. [Google Scholar] [CrossRef] [Green Version]
- Steinberg, P.D.; Brett, M.T.; Bechtold, J.S.; Richey, J.E.; Porensky, L.; Smith, S.N. The influence of watershed characteristics on nitrogen export to and marine fate in Hood Canal, Washington, USA. Biogeochemistry 2011, 106, 415–433. [Google Scholar] [CrossRef]
- Marchant, H.K.; Mohr, W.; Kuypers, M.M.M. Recent advances in marine N-cycle studies using 15N labeling methods. Curr. Opin. Biotechnol. 2016, 41, 53–59. [Google Scholar] [CrossRef]
- Robertson, E.K.; Bartoli, M.; Brüchert, V.; Dalsgaard, T.; Hall, P.O.J.; Hellemann, D.; Hietanen, S.; Zilius, M.; Conley, D.J. Application of the isotope pairing technique in sediments: Use, challengers, and new directions. Limnol. Oceanogr. Methods 2019, 17, 112–136. [Google Scholar] [CrossRef]
- Massé, S.; Botrel, M.; Walsh, D.A.; Maranger, R. Annual nitrification dynamics in a seasonally ice-covered lake. PLoS ONE 2019, 14, e0213748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Könneke, M.; Bernhard, A.E.; de la Torre, J.R.; Walker, C.B.; Waterbury, J.B.; Stahl, D.A. Isolation of an autotrophic ammonia-oxidizing marine archaeon. Nature 2005, 437, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Martens-Habbena, W.; Berube, P.M.; Urakawa, H.; de la Torre, J.R.; Stahl, D.A. Ammonia oxidation kinetics determine niche separation of nitrifying Archaea and Bacteria. Nature 2009, 461, 976–979. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Jacob, C.; Michelutti, N.; Paterson, A.M.; Cumming, B.F.; Keller, W.B.; Smol, J.P. The browning and re-browning of lakes: Divergent lake-water organic carbon trends linked to acid deposition and climate change. Sci. Rep. 2019, 9, 16676. [Google Scholar] [CrossRef] [PubMed]
- Monteith, D.T.; Stoddard, J.L.; Evans, C.D.; de Wit, H.A.; Forsius, M.; Hogåsen, T.; Wilander, A.; Sjelkvåle, B.L.; Jeffries, D.S.; Vuorenmaa, J.; et al. Dissolved organic carbon trends resulting from changes in atmospheric deposition chemistry. Nature 2007, 450, 537–540. [Google Scholar] [CrossRef]
- Garmo, Ø.A.; Skjelkvåle, B.L.; de Wit, H.A.; Colombo, L.; Curtis, C.; Fölster, J.; Hoffmann, A.; Hruska, J.; Hogåsen, T.; Jeffries, D.S.; et al. Trends in surface water chemistry in acidified areas in Europe and North America from 1990 to 2008. Water Air Soil Poll. 2014, 225, 1880. [Google Scholar] [CrossRef] [Green Version]
- Cavicchioli, R.; Ripple, W.J.; Timmis, K.N.; Azam, F.; Bakken, L.R.; Baylis, M.; Behrenfeld, M.J.; Boetius, A.; Boyd, P.W.; Classen, A.T.; et al. Scientists’ warning to humanity: Microorganisms and climate change. Nat. Rev. Microb. 2019, 17, 569–586. [Google Scholar] [CrossRef] [Green Version]
- Hansell, D.A. Recalcitrant dissolved organic carbon fractions. Annu. Rev. Mar. Sci. 2013, 5, 421–445. [Google Scholar] [CrossRef] [PubMed]
- Azam, F.; Fenchel, T.; Field, J.G.; Gray, I.S.; Meyerreil, L.A.; Thingstad, F. The ecological role of water-column microbes in the sea. Mar. Ecol Prog Ser. 1983, 10, 257–263. [Google Scholar] [CrossRef]
- Arandia-Gorostidi, N.; Weber, P.K.; Alonso-Sáez, L.; Morán, X.A.G.; Mayali, X. Elevated temperature increases carbon and nitrogen fluxes between phytoplankton and heterotrophic bacteria through physical attachment. ISME J. 2016, 11, 641–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durham, B.P.; Sharma, S.; Luo, H.; Smith, C.B.; Amin, S.A.; Bender, S.J.; Dearth, S.P.; Van Mooy, B.A.S.; Campagna, S.R.; Kujawinski, E.B.; et al. Cryptic carbon and sulfur cycling between surface ocean plankton. Proc. Natl. Acad. Sci. USA 2015, 12, 453–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amin, S.A.; Hmelo, L.R.; van Tol, H.M.; Durham, B.P.; Carlson, L.T.; Heal, K.R.; Morales, R.L.; Berthiaume, C.T.; Parker, M.S.; Djunaedi, B.; et al. Interaction and signaling between a cosmopolitan phytoplankton and associated bacteria. Nature 2015, 522, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Hipsey, M.R.; Gal, G.; Arhonditsis, G.B.; Carey, C.C.; Elliott, J.A.; Frassl, M.A.; Janse, J.H.; Mora, L.; Robson, B.J. A system of metrics for the assessment and improvement of aquatic ecosystem models. Environ. Model. Softw. 2020, 128, 104697. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameter | Unit | PPr | Terrestrial | Reference/Derived from |
|---|---|---|---|---|
| Benthic and pelagic PPr | mg C (m−2 d−1) | 253 | - | Brett et al. [10] |
| Bacterial growth efficiency (BGE) | biomass produced/mass ingested | 0.5 | 0.1 | del Giorgio and Cole [15] |
| DOC loading | mg C (m−2 d−1) | 63 | 62 | Brett et al. [10] |
| DOC first-order loss rate (σ) | d−1 | 0.1 | 0.001 | Hanson et al. [4]; Brett et al. [10]; Wetzel [12] |
| HRT | yrs | 0.6 | 0.6 | Brett and Benjamin [11] |
| Flushing rate (ρ) | d−1 | 0.005 | 0.005 | Brett and Benjamin [11] |
| Areal hydrologic loading | L C (m−2 d−1) | - | 5 | Brett and Benjamin [11] |
| Removal | unitless | 0.96 | 0.18 | σ/(σ + ρ) |
| Removed | mg C (m−2 d−1) | 60.5 | 11.2 | DOC load Removal |
| Percent metabolized | % | 84 | 16 | Removed/∑ Removed 100 |
| Remaining DOC flux | mg C (m−2 d−1) | 2.5 | 50.8 | DOC load—Removed |
| DOC concentration remaining | mg C L−1 | 0.5 | 10.1 | Remaining/areal load |
| Percent remaining | % | 5 | 95 | DOC conc./∑ DOC conc. 100 |
| Bacteria production from DOC | mg C (m−2 d−1) | 30.2 | 1.1 | DOC load Removal BGE |
| Bacteria production from DOC% | % | 96 | 4 | Bact. prod./∑ Bact. prod. 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olofsson, M.; Power, M.E.; Stahl, D.A.; Vadeboncoeur, Y.; Brett, M.T. Cryptic Constituents: The Paradox of High Flux–Low Concentration Components of Aquatic Ecosystems. Water 2021, 13, 2301. https://doi.org/10.3390/w13162301
Olofsson M, Power ME, Stahl DA, Vadeboncoeur Y, Brett MT. Cryptic Constituents: The Paradox of High Flux–Low Concentration Components of Aquatic Ecosystems. Water. 2021; 13(16):2301. https://doi.org/10.3390/w13162301
Chicago/Turabian StyleOlofsson, Malin, Mary E. Power, David A. Stahl, Yvonne Vadeboncoeur, and Michael T. Brett. 2021. "Cryptic Constituents: The Paradox of High Flux–Low Concentration Components of Aquatic Ecosystems" Water 13, no. 16: 2301. https://doi.org/10.3390/w13162301
APA StyleOlofsson, M., Power, M. E., Stahl, D. A., Vadeboncoeur, Y., & Brett, M. T. (2021). Cryptic Constituents: The Paradox of High Flux–Low Concentration Components of Aquatic Ecosystems. Water, 13(16), 2301. https://doi.org/10.3390/w13162301