
Unexpected Diversity of Feeding Modes among Chisel-Mouthed Ethiopian Labeobarbus (Cyprinidae)

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
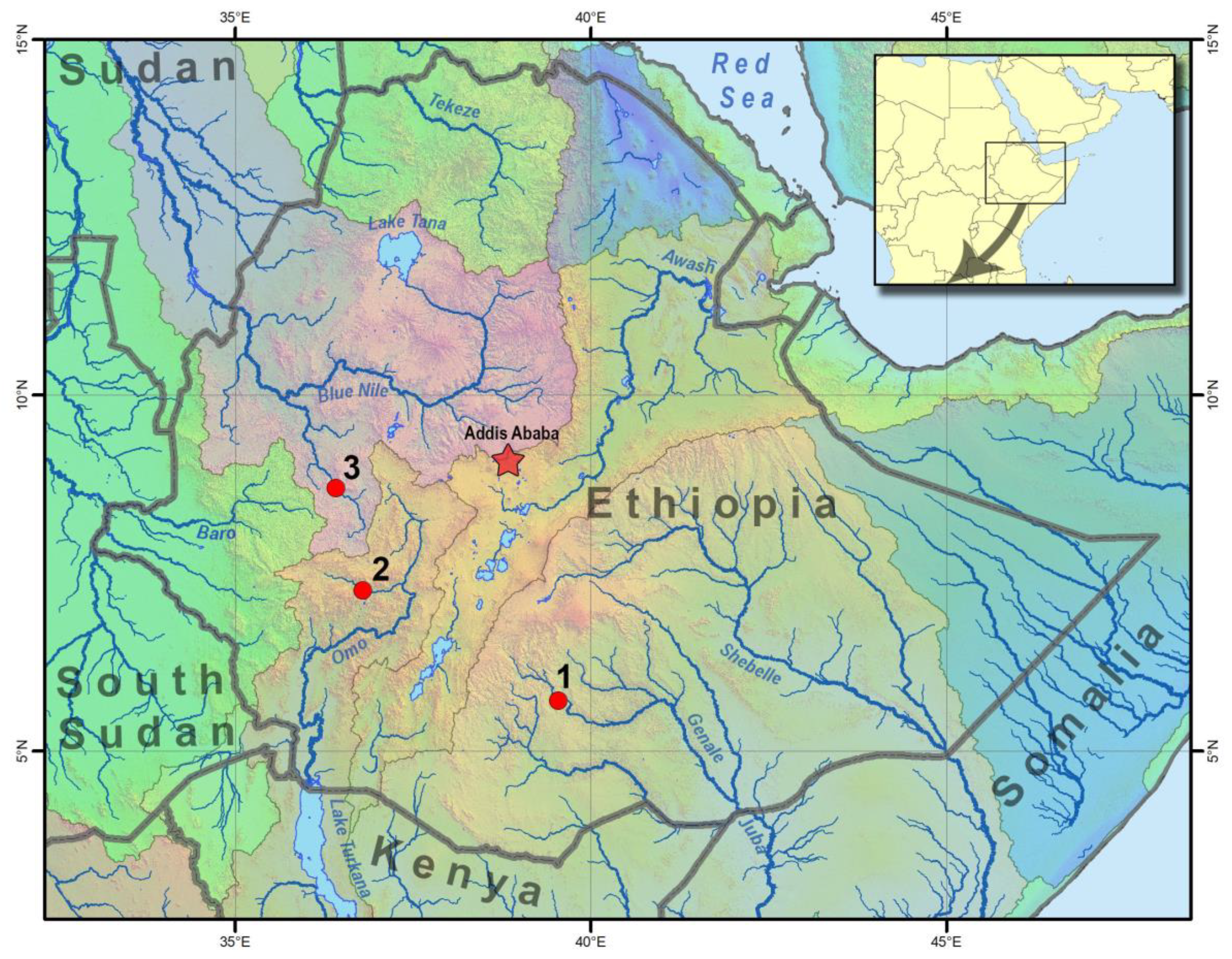
2.1. Study Area and Sampling
2.2. Examination of Morphological Characters
2.3. Diet and Stable Isotopes
3. Statistical Analysis
4. Results
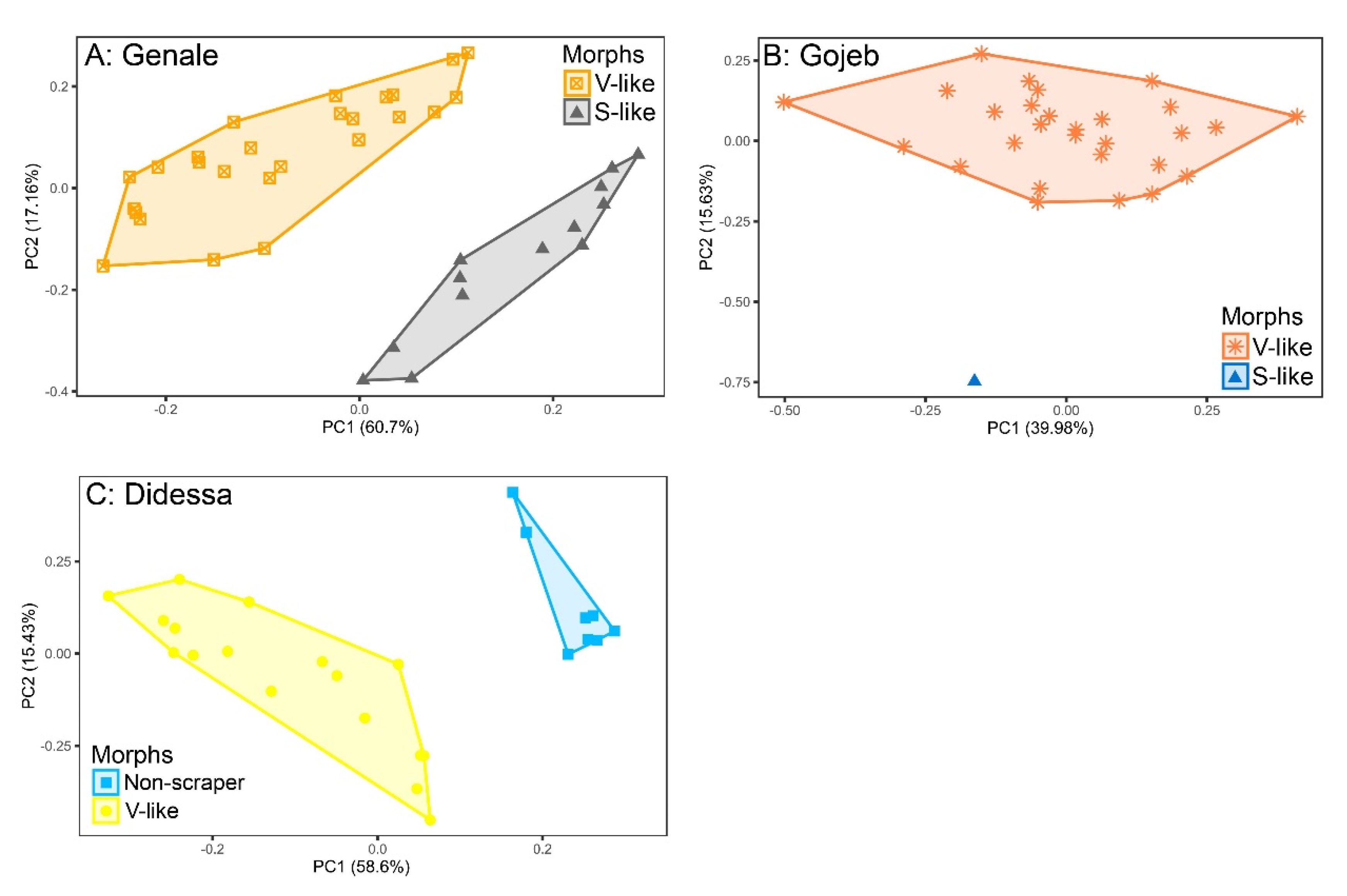
4.1. Morphology
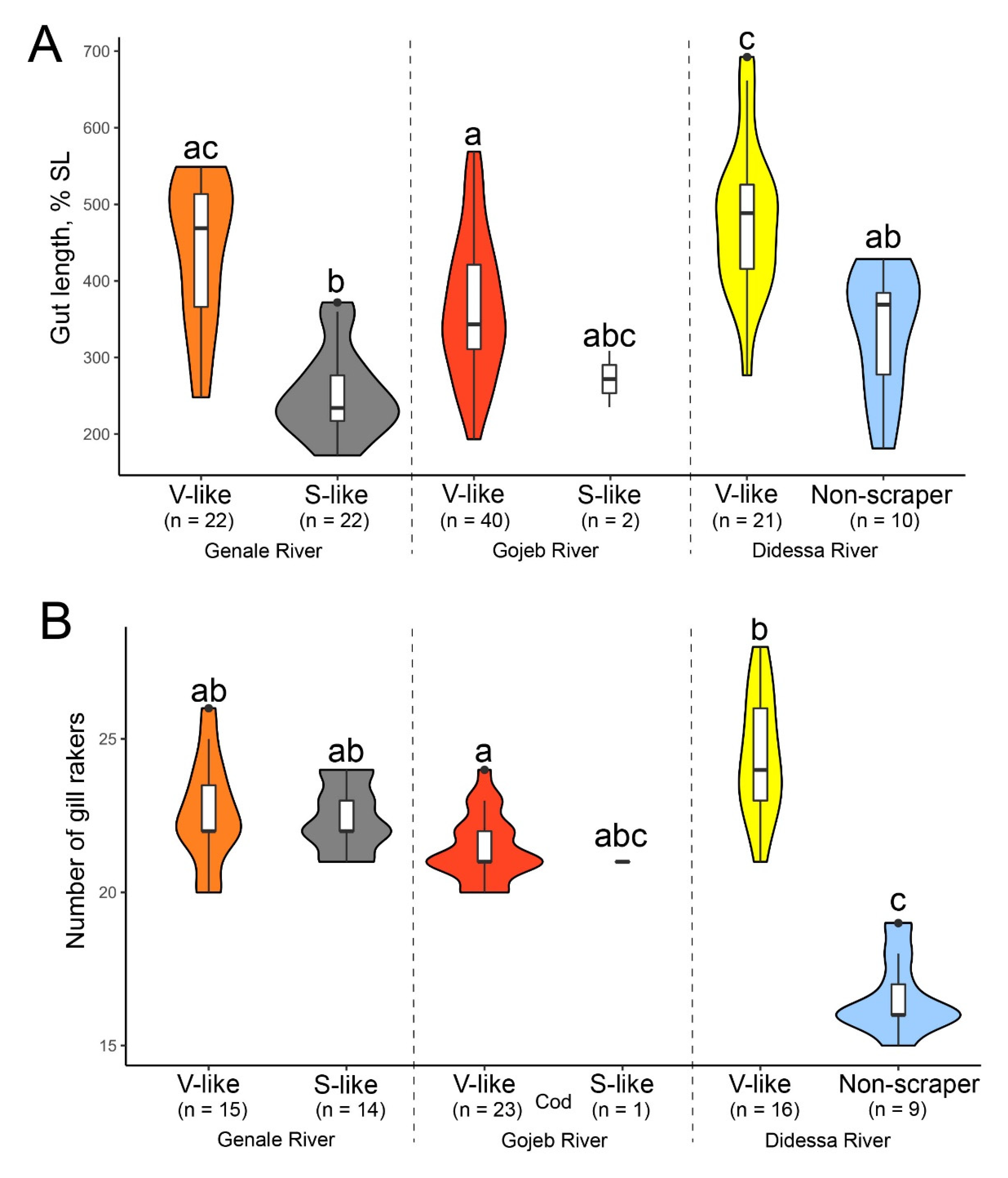
4.2. Gut Length
4.3. Gill Rakers
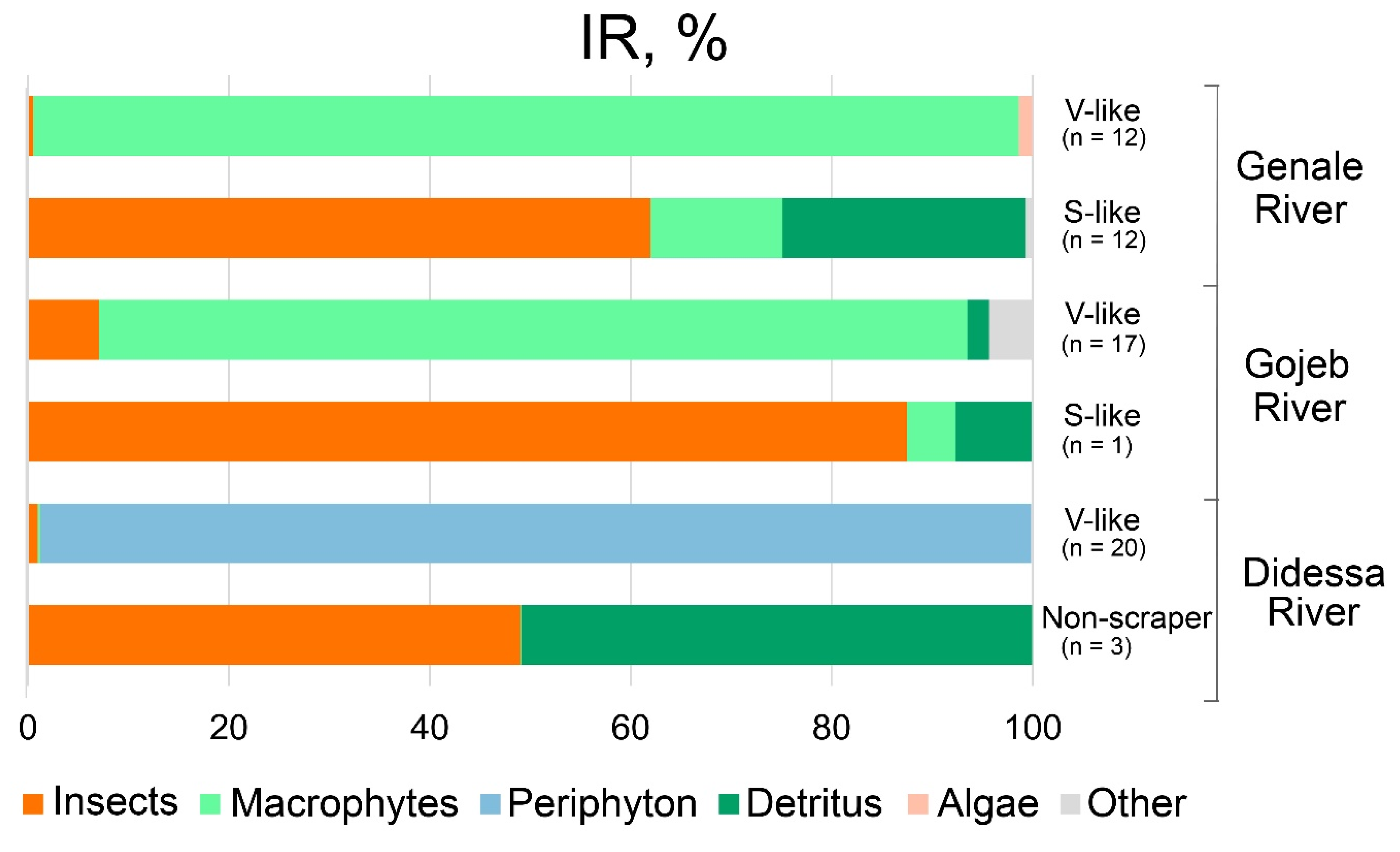
4.4. Diet
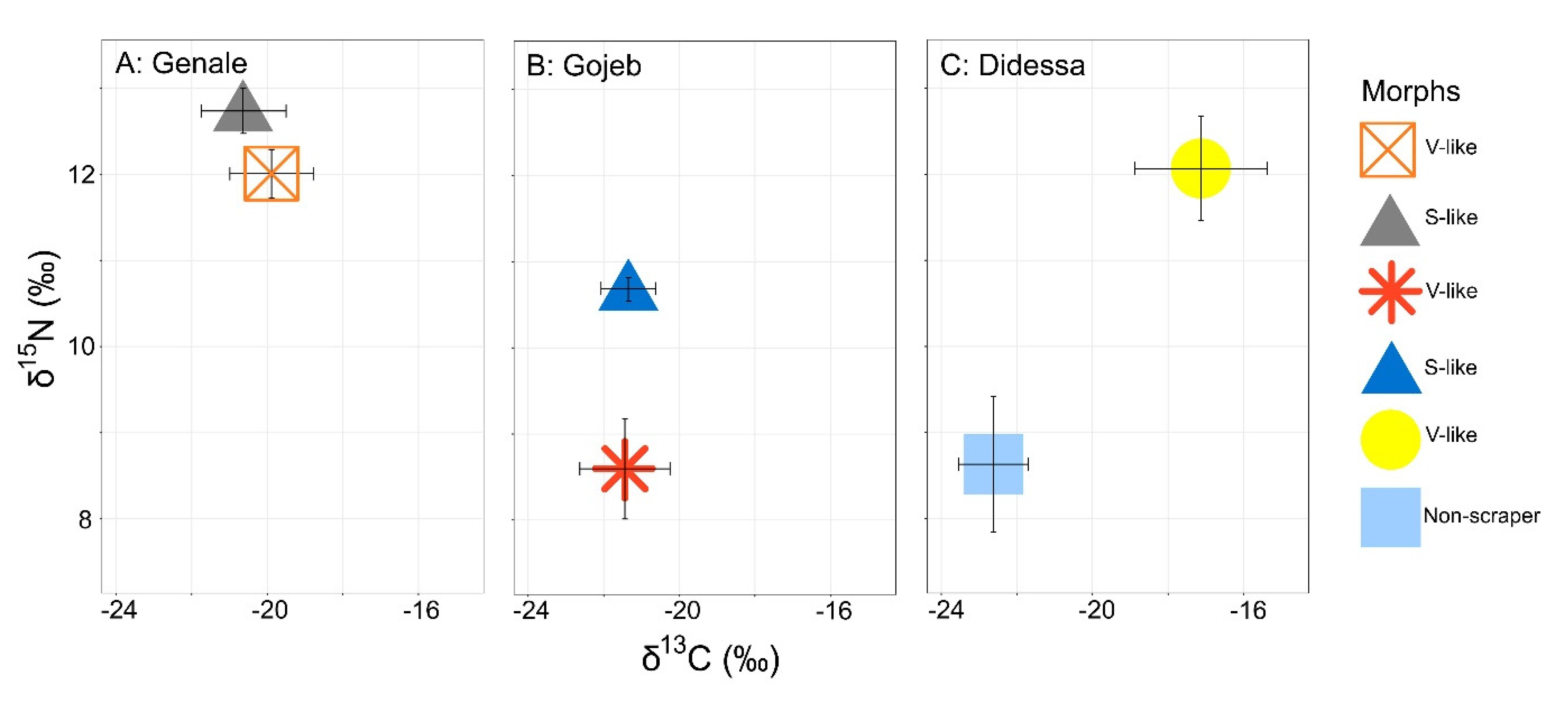
4.5. Stable Isotopes
5. Discussion
5.1. Trophic Resource Partitioning
5.2. Parallel Ecological Speciation among Ethiopian Labeobarbus Scrapers
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schluter, D. The Ecology of Adaptive Radiation; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Seehausen, O. African cichlid fish: A model system in adaptive radiation research. Proc. R. Soc. B Biol. Sci. 2006, 273, 1987–1998. [Google Scholar] [CrossRef] [Green Version]
- Stroud, J.T.; Losos, J.B. Ecological opportunity and adaptive radiation. Ann. Rev. Ecol. Evol. Syst. 2016, 47, 507–532. [Google Scholar] [CrossRef]
- Gillespie, R.G.; Bennett, G.M.; De Meester, L.; Feder, J.L.; Fleischer, R.C.; Harmon, L.J.; Wogan, G.O. Comparing adaptive radiations across space, time, and taxa. J. Hered. 2020, 111, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagelkerke, L.A.J.; Sibbing, F.A.; van den Boogaart, J.G.M.; Lammens, E.H.R.R.; Osse, J.W.M. The barbs (Barbus spp.) of Lake Tana: A forgotten species flock? Env. Biol. Fish. 1994, 39, 1–22. [Google Scholar] [CrossRef]
- Mina, M.V.; Mironovsky, A.N.; Dgebuadze, Y. Lake Tana large barbs: Phenetics, growth and diversification. J. Fish Biol. 1996, 48, 383–404. [Google Scholar] [CrossRef]
- Nagelkerke, L.A.J.; Sibbing, F.A. Reproductive segregation among the large barbs (Barbus intermedius complex) of Lake Tana, Ethiopia. An example of intralacustrine speciation? J. Fish. Biol. 1996, 49, 1244–1266. [Google Scholar] [CrossRef]
- de Graaf, M.; Megens, H.-J.; Samallo, J.; Sibbing, F.A. Evolutionary origin of Lake Tana’s (Ethiopia) small Barbus species: Indications of rapid ecological divergence and speciation. Anim. Biol. 2007, 57, 39–48. [Google Scholar] [CrossRef]
- de Graaf, M.; Dejen, E.; Osse, J.W.M.; Sibbing, F.A. Adaptive radiation of Lake Tana’s (Ethiopia) Labeobarbus species flock (Pisces, Cyprinidae). Mar. Freshw. Res. 2008, 59, 391–407. [Google Scholar] [CrossRef]
- Dimmick, W.W.; Berendzen, P.B.; Golubtsov, A.S. Genetic comparison of three Barbus (Cyprinidae) morphotypes from the Genale River, Ethiopia. Copeia 2001, 4, 1123–1129. [Google Scholar] [CrossRef]
- Sibbing, F.A.; Nagelkerke, L.A.J. Resource partitioning by Lake Tana barbs predicted from fish morphometrics and prey characteristics. Rev. Fish. Biol. Fisher. 2001, 10, 393–437. [Google Scholar] [CrossRef]
- Stiassny, M.L.J.; Getahun, A. An overview of labeonin relationships and the phylogenetic placement of the Afro-Asian genus Garra Hamilton, 1822 (Teleostei: Cyprinidae), with the description of five new species of Garra from Ethiopia, and a key to all African species. Zool. J. Linn. Soc. 2007, 150, 41–83. [Google Scholar] [CrossRef]
- Golubtsov, A.S.; Cherenkov, S.E.; Tefera, F. High morphological diversity of the genus Garra in the Sore River (the White Nile Basin, Ethiopia): One more cyprinid species flock? J. Ichthyol. 2012, 52, 817–820. [Google Scholar] [CrossRef]
- Golubtsov, A.S.; Korostelev, N.B.; Levin, B.A. Monsters with a shortened vertebral column: A population phenomenon in radiating fish Labeobarbus (Cyprinidae). PLoS ONE 2021, 16, e0239639. [Google Scholar] [CrossRef]
- Levin, B.A.; Casal-López, M.; Simonov, E.; Dgebuadze, Y.Y.; Mugue, N.S.; Tiunov, A.V.; Doadrio, I.; Golubtsov, A.S. Adaptive radiation of barbs of the genus Labeobarbus (Cyprinidae) in an East African river. Freshw. Biol. 2019, 64, 1721–1736. [Google Scholar] [CrossRef]
- Levin, B.A.; Simonov, E.; Dgebuadze, Y.Y.; Levina, M.; Golubtsov, A.S. In the rivers: Multiple adaptive radiations of cyprinid fishes (Labeobarbus) in Ethiopian Highlands. Sci. Rep. 2020, 10, 7192. [Google Scholar] [CrossRef]
- Levin, B.; Simonov, E.; Franchini, P.; Mugue, N.; Golubtsov, A.; Meyer, A. Adaptive radiation and burst speciation of hillstream cyprinid fish Garra in African river. Mol. Ecol. 2021, in press. [Google Scholar] [CrossRef]
- Nikolskii, G.V. The Ecology of Fishes; Academic Press: London, UK, 1963. [Google Scholar]
- Wootton, R.J. Ecology of Teleost Fishes; Chapman & Hall: London, UK, 1990. [Google Scholar]
- Bănărescu, P.; Coad, B.W. Cyprinids of Eurasia. In Cyprinid Fishes; Winfield, I.J., Nelson, J.S., Eds.; Chapman & Hall: London, UK, 1991; pp. 127–155. [Google Scholar] [CrossRef]
- Skelton, P.H.; Tweddle, D.; Jackson, P.B.N. Cyprinids of Africa. In Cyprinid Fishes; Winfield, I.J., Nelson, J.S., Eds.; Chapman & Hall: London, UK, 1991; pp. 211–239. [Google Scholar] [CrossRef]
- Mayden, R.L. Cyprinids of the New World. In Cyprinid Fishes; Winfield, I.J., Nelson, J.S., Eds.; Chapman & Hall: London, UK, 1991; pp. 240–263. [Google Scholar] [CrossRef]
- Howes, G. Anatomy and evolution of the jaws in the semiplotine carps with a review of the genus Cyprinion Hechel, 1843 (Teleostei: Cyprinidae). Bull. Nat. Hist. Mus. Zool. 1982, 42, 299–335. [Google Scholar]
- Chen, Y.-Y. Anatomy and evolution of the jaws in the semiplotine carps with a review of the genus Onychostoma Günther, 1896. Bull. Nat. Hist. Mus. Zool. 1989, 42, 299–335. [Google Scholar]
- Nikolskii, G.V. Special Ichthyology: Israel Program for Scientific Translation; Today & Tomorrow Printers & Publishers: New Delhi, India, 1989. [Google Scholar]
- Komarova, A.S.; Rozanova, O.L.; Levin, B.A. Trophic resource partitioning by sympatric ecomorphs of Schizopygopsis (Cyprinidae) in a young Pamir Mountain lake: Preliminary results. Ichthyol. Res. 2021, 68, 191–197. [Google Scholar] [CrossRef]
- Matthes, H. A comparative study of the feeding mechanisms of some African Cyprinidae (Pisces, Cypriniformes). Bijdr. Dierk. 1963, 33, 3–35. [Google Scholar] [CrossRef] [Green Version]
- Levin, B.A.; Freyhof, J.; Lajbner, Z.; Perea, S.; Abdoli, A.; Gaffaroğlu, M.; Özuluğ, M.; Rubenyan, H.R.; Salnikov, V.B.; Doadrio, I. Phylogenetic relationships of the algae scraping cyprinid genus Capoeta (Teleostei: Cyprinidae). Mol. Phylogenet. Evol. 2012, 62, 542–549. [Google Scholar] [CrossRef]
- Vreven, E.J.; Musschoot, T.; Snoeks, J.; Schliewen, U.K. The African hexaploid Torini (Cypriniformes: Cyprinidae): Review of a tumultuous history. Zool. J. Linn. Soc. 2016, 177, 231–305. [Google Scholar] [CrossRef] [Green Version]
- Vreven, E.J.; Musschoot, T.; Decru, E.; Wamuini Lunkayilakio, S.; Obiero, K.; Cerwenka, A.F.; Schliewen, U.K. The complex origins of mouth polymorphism in the Labeobarbus (Cypriniformes: Cyprinidae) of the Inkisi River basin (Lower Congo, DRC, Africa): Insights from an integrative approach. Zool. J. Linn. Soc. 2019, 186, 414–482. [Google Scholar] [CrossRef]
- Lammens, E.H.R.R.; Hoogenboezem, W. Diets and feeding behaviour. In Cyprinid Fishes; Winfield, I.J., Nelson, J.S., Eds.; Chapman & Hall: London, UK, 1991; pp. 353–376. [Google Scholar] [CrossRef]
- Yang, L.; Arunachalam, M.; Sado, T.; Levin, B.A.; Golubtsov, A.S.; Freyhof, J.; Friel, J.P.; Chen, W.J.; Hirt, M.V.; Manickam, R.; et al. Molecular phylogeny of the cyprinid tribe Labeonini (Teleostei: Cypriniformes). Mol. Phylogenet. Evol. 2012, 65, 362–379. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Sado, T.; Hirt, M.V.; Pasco-Viel, E.; Arunachalam, M.; Li, J.; Wang, X.; Freyhof, J.; Saitoh, K.; Simons, A.M.; et al. Phylogeny and polyploidy: Resolving the classification of cyprinine fishes (Teleostei: Cypriniformes). Mol. Phylogenet. Evol. 2015, 85, 97–116. [Google Scholar] [CrossRef] [PubMed]
- Lévêque, C.; Daget, J. Cyprinidae. In Check-List of the Freshwater Fishes of Africa. CLOFFA 1; Daget, J., Gosse, J.-P., Thys van den Audenaerde, D.F.E., Eds.; ISNB: Bruxelles, Belgium; MRAC: Tervuren, Belgium; ORSTOM: Paris, France, 1984; pp. 217–342. [Google Scholar]
- Froese, R.; Pauly, D. (Eds.) FishBase: Stockholm, Sweden, 2021. Available online: www.fishbase.org (accessed on 21 March 2021).
- Tsigenopoulos, C.S.; Kasapidis, P.; Berrebi, P. Phylogenetic relationships of hexaploid large-sized barbs (genus Labeobarbus, Cyprinidae) based on mtDNA data. Mol. Phylogenet. Evol. 2010, 56, 851–856. [Google Scholar] [CrossRef] [PubMed]
- Levin, B.A.; Golubtsov, A.S.; Dgebuadze, Y.Y.; Mugue, N.S. New evidence of homoplasy within the African genus Varicorhinus (Cyprinidae): An independent origin of specialized scraping forms in the adjacent drainage systems of Ethiopia inferred from mtDNA analysis. Afr. Zool. 2013, 48, 400–406. [Google Scholar] [CrossRef]
- Eschmeyer, W.N.; Fricke, R.; van der Laan, R. (Eds.) Catalog of Fishes: Genera, Species, References. 2021. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 28 June 2021).
- Oellermann, L.K.; Skelton, P.H. Hexaploidy in yellowfish species (Barbus, Pisces, Cyprinidae) from southern Africa. J. Fish Biol. 1990, 37, 105–115. [Google Scholar] [CrossRef]
- Golubtsov, A.S.; Krysanov, E.Y. Karyological study of some cyprinid species from Ethiopia. The ploidy differences between large and small Barbus of Africa. J. Fish Biol. 1993, 42, 445–455. [Google Scholar] [CrossRef]
- Guégan, J.-F.; Rab, P.; Machordom, A.; Doadrio, I. New evidence of hexaploidy in ’large’ African Barbus with some considerations on the origin of hexaploidy. J. Fish Biol. 1995, 47, 192–198. [Google Scholar] [CrossRef]
- Krysanov, E.Y.; Golubtsov, A.S. Karyotypes of some Ethiopian Barbus and Varicorhinus from the Nile Basin including Lake Tana morphotypes. Folia Zool. 1996, 45 (Suppl. 1), 67–75. [Google Scholar]
- Naran, D.; Skelton, P.H.; Villet, M.H. Karyology of three evolutionarily hexaploid southern African species of yellowfish, Labeobarbus Rüppell, 1836 (Cyprinidae). Afr. Zool. 2007, 42, 254–260. [Google Scholar] [CrossRef]
- Mina, M.V.; Mironovsky, A.N.; Golubtsov, A.S.; Dgebuadze, Y.Y. The ’Barbus’ intermedius species flock in Lake Tana (Ethiopia): II-Morphological diversity of" large barbs" from Lake Tana and neighbouring areas: Homoplasies or synapomorphies? Ital. J. Zool. 1998, 65 (Suppl. 1), 9–14. [Google Scholar] [CrossRef] [Green Version]
- Levin, B.A. New data on morphology of the African scraping feeder Varicorhinus beso (Osteichthyes: Cyprinidae) with the special reference to specialized traits. J. Ichthyol. 2012, 52, 908–923. [Google Scholar] [CrossRef]
- Banister, K. Three new species of Varicorhinus (Pisces, Cyprinidae) from Africa. Bull. Nat. Hist. Mus. Zool. 1984, 47, 273–282. [Google Scholar]
- Reid, G.M. A Revision of African Species of Labeo and Re-Definition of the Genus; Verlag von J. Cramer: Braunschweig, Germany, 1985. [Google Scholar]
- Golubtsov, A.S.; Dgebuadze, Y.Y.; Mina, M.V. Fishes of the Ethiopian Rift Valley. In Ethiopian Rift Valley Lakes; Tudorancea, C., Taylor, W.D., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2002; pp. 167–258. [Google Scholar]
- Mironovsky, A.N.; Mina, M.V.; Dgebuadze, Y.Y. Large African barbs with hypertrophied lips and their relationship with generalized forms of species of the genus Barbus (Labeobarbus auctorum). J. Ichthyol. 2019, 59, 327–335. [Google Scholar] [CrossRef]
- Hubbs, C.L.; Lagler, K.F. Fishes of the Great Lakes Region; Michigan University Press: Ann Arbor, MI, USA, 1958. [Google Scholar]
- Levin, B.A.; Rubenyan, A.R.; Salnikov, V.B. Phenetic diversity of khramulya Capoeta capoeta (Ostariophysi, Cyprinidae). J. Ichthyol. 2005, 45, 754–767. [Google Scholar]
- Mina, M.V.; Levin, B.A.; Mironovsky, A.N. On the possibility of using character estimates obtained by different operators in morphometric studies of fish. J. Ichthyol. 2005, 45, 284–294. [Google Scholar]
- Natarajan, A.V.; Jhingran, A.G. Index of preponderance—A method of grading the food elements in the stomach analysis of fishes. Indian J. Fish. 1961, 8, 54–59. [Google Scholar]
- Popova, O.A.; Reshetnikov, Y.S. On Complex Indices in Investigation of Fish Feeding. J. Ichthyol. 2011, 51, 686–691. [Google Scholar] [CrossRef]
- Post, D.M.; Layman, C.A.; Arrington, D.A.; Takimoto, G.; Quattrochi, J.; Montana, C.G. Getting to the fat of the matter: Models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecologia 2007, 152, 179–189. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development for R; RStudio: PBC, Boston, MA, USA; Available online: http://www.rstudio.com (accessed on 20 March 2021).
- Comtois, D. Summarytools: Tools to Quickly and Neatly Summarize Data. R Package Version 0.8. 72018. 2018. Available online: https://CRAN.R-project.org/package=summarytools (accessed on 20 March 2021).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Pohlert, T.; Pohlert, M.T. Package ‘PMCMR’. R Package Version. 2018. Available online: https://cran.r-project.org/web/packages/PMCMR/index.html (accessed on 13 August 2021).
- Ogle, D.; Ogle, M.D. Package ‘FSA’. CRAN Repos. 2017. Available online: https://cran.microsoft.com/snapshot/2017-08-14/web/packages/FSA/FSA.pdf (accessed on 13 August 2021).
- McCutchan, J.H., Jr.; Lewis, W.M., Jr.; Kendall, C.; McGrath, C.C. Variation in trophic shift for stable isotope ratios of carbon, nitrogen, and sulfur. Oikos 2003, 102, 378–390. [Google Scholar] [CrossRef]
- Wagner, C.E.; McIntyre, P.B.; Buels, K.S.; Gilbert, D.M.; Michel, E. Diet predicts intestine length in Lake Tanganyika’s cichlid fishes. Funct. Ecol. 2009, 23, 1122–1131. [Google Scholar] [CrossRef]
- Zandonà, E.; Auer, S.K.; Kilham, S.S.; Reznick, D.N. Contrasting population and diet influences on gut length of an omnivorous tropical fish, the Trinidadian guppy (Poecilia reticulata). PLoS ONE 2015, 10, e0136079. [Google Scholar] [CrossRef] [Green Version]
- Golubtsov, A.S. Fish ‘Species Flocks’ in Rivers and Lakes: Sympatric Divergence in Poor Fauna Fish Communities as Particular Modus of Evolution. In Relevant Problems of Contemporary Ichthyology (To 100 Jubilee of G. V. Nikolsky); Pavlov, D.S., Dgebuadze, Y.Y., Shatunovsky, M.I., Eds.; KMK Scientific Press: Moscow, Russia, 2010; pp. 96–123. [Google Scholar]
- Kisekelwa, T.; Snoeks, J.; Zamba, A.I.; Amzati, G.S.; Isumbisho, M.; Masilya, P.M.; Lemmens, P.; Vreven, E. Association between Labeobarbus spp. (Teleostei: Cyprinidae) and environmental variables in the Luhoho basin (Eastern Congo River basin; DRC). J. Fish Biol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Azim, M.E.; Beveridge, M.C.M.; van Dam, A.A.; Verdegem, M.C. Periphyton and aquatic production: An introduction. In Periphyton: Ecology, Exploitation and Management; Azim, M.E., Verdegem, M.C.J., van Dam, A.A., Beveridge, M.C.M., Eds.; Centre for Agriculture and Bioscience International: Wallingford, UK, 2005; pp. 1–13. [Google Scholar]
- Becker, E.W. Micro-algae as a source of protein. Biotechnol. Adv. 2007, 25, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Saikia, S.K. Review on periphyton as mediator of nutrient transfer in aquatic ecosystems. Ecol. Balk. 2011, 3, 65–78. [Google Scholar]
- Akin, S.; Turan, H.; Kaymak, N. Does diet variation determine the digestive tract length of Capoeta banarescui Turan, Kottelat, Ekmekci and Imamoglu, 2006. J. Appl. Ichthyol. 2016, 32, 883–892. [Google Scholar] [CrossRef]
- Beshera, K.A.; Harris, P.M.; Mayden, R.L. Novel evolutionary lineages in Labeobarbus (Cypriniformes; Cyprinidae) based on phylogenetic analyses of mtDNA sequences. Zootaxa 2016, 4093, 363–381. [Google Scholar] [CrossRef]
- Nikolskii, G.V. Special Ichthyology; Sov. Nauka: Moscow, Russia, 1950. [Google Scholar]
- Popov, A.V. Morphofunctional adaptations of the Pamir Schizopygopsis stolizkai Steind. in lake Yashil-kul. Vopr Ikhtiol 1968, 8, 15–30. (In Russian) [Google Scholar]
- Thompson, C.E.; Taylor, E.B.; McPhail, J.D. Parallel evolution of lake-stream pairs of threespine sticklebacks (Gasterosteus) inferred from mitochondrial DNA variation. Evolution 1997, 51, 1955–1965. [Google Scholar] [PubMed]
- Rüber, L.; Verheyen, E.; Meyer, A. Replicated evolution of trophic specializations in an endemic cichlid fish lineage from Lake Tanganyika. Proc. Nat. Acad. Sci. USA 1999, 96, 10230–10235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alekseyev, S.S.; Samusenok, V.P.; Matveev, A.N.; Pichugin, M.Y. Diversification, sympatric speciation, and trophic polymorphism of Arctic charr, Salvelinus alpinus complex, in Transbaikalia. Env. Biol. Fish. 2002, 64, 97–114. [Google Scholar] [CrossRef]
- Derome, N.; Bernatchez, L. The transcriptomics of ecological convergence between 2 limnetic coregonine fishes (Salmonidae). Mol. Biol. Evol. 2006, 23, 2370–2378. [Google Scholar] [CrossRef]
- Elmer, K.R.; Fan, S.; Kusche, H.; Luise Spreitzer, M.; Kautt, A.F.; Franchini, P.; Meyer, A. Parallel evolution of Nicaraguan crater lake cichlid fishes via non-parallel routes. Nat. Commun. 2014, 5, 5168. [Google Scholar] [CrossRef] [Green Version]
- Burress, E.D.; Holcomb, J.M.; Bonato, K.O.; Armbruster, J.W. Body size is negatively correlated with trophic position among cyprinids. R. Soc. Open Sci. 2016, 3, 150652. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ecomorphs (SL Range, mm) | Gut Length (n) | Osteology (n) | Gill Rakers (n) | Stable Isotopes (n) | Diet (n) |
|---|---|---|---|---|---|
| Scrapers within L. gananensis/jubae complex—Genale River (Indian Ocean basin) | |||||
| V-like (89–349) | 22 | 26 | 15 | 13 | 12 |
| S-like (82–360) | 22 | 13 | 14 | 14 | 12 |
| Scrapers within L. cf. intermedius complex—Gojeb River (Omo-Turkana basin) | |||||
| V-like (142–361) | 40 | 28 | 23 | 32 | 17 |
| S-like (251–256) | 2 | 1 | 1 | 2 | 1 |
| L. beso and L. cf. intermedius—Didessa River (Blue Nile basin) | |||||
| V-like scraper (L. beso) (65–230) | 21 | 17 | 16 | 24 | 20 |
| Non-scraper, omnivorous ecomorph (L. cf. intermedius) (90–378) | 10 | 8 | 9 | 23 | 3 |
| Total | 117 | 85 | 78 | 108 | 65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levin, B.A.; Komarova, A.S.; Rozanova, O.L.; Golubtsov, A.S. Unexpected Diversity of Feeding Modes among Chisel-Mouthed Ethiopian Labeobarbus (Cyprinidae). Water 2021, 13, 2345. https://doi.org/10.3390/w13172345
Levin BA, Komarova AS, Rozanova OL, Golubtsov AS. Unexpected Diversity of Feeding Modes among Chisel-Mouthed Ethiopian Labeobarbus (Cyprinidae). Water. 2021; 13(17):2345. https://doi.org/10.3390/w13172345
Chicago/Turabian StyleLevin, Boris A., Aleksandra S. Komarova, Oksana L. Rozanova, and Alexander S. Golubtsov. 2021. "Unexpected Diversity of Feeding Modes among Chisel-Mouthed Ethiopian Labeobarbus (Cyprinidae)" Water 13, no. 17: 2345. https://doi.org/10.3390/w13172345
APA StyleLevin, B. A., Komarova, A. S., Rozanova, O. L., & Golubtsov, A. S. (2021). Unexpected Diversity of Feeding Modes among Chisel-Mouthed Ethiopian Labeobarbus (Cyprinidae). Water, 13(17), 2345. https://doi.org/10.3390/w13172345