
Influence of Intensive Agriculture on Benthic Macroinvertebrate Assemblages and Water Quality in the Aconcagua River Basin (Central Chile)



Abstract
:1. Introduction
2. Material and Methods
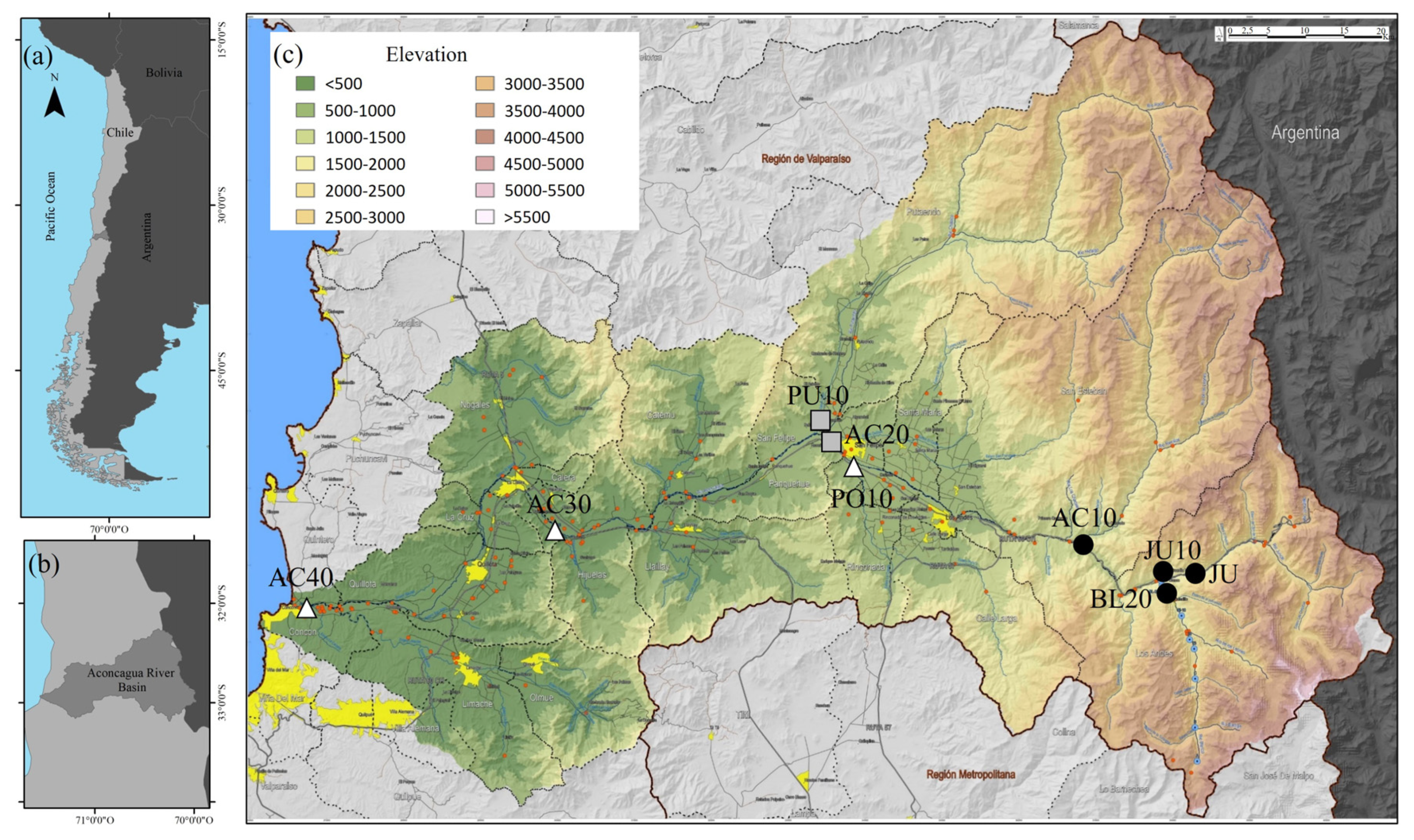
2.1. Study Area
2.2. Land Use and Sampling Sites
2.3. Field Sampling and Data Collection
2.4. Data Analyses
3. Results
3.1. Environmental Variables and Water Quality
3.2. Macroinvertebrate Assemblages
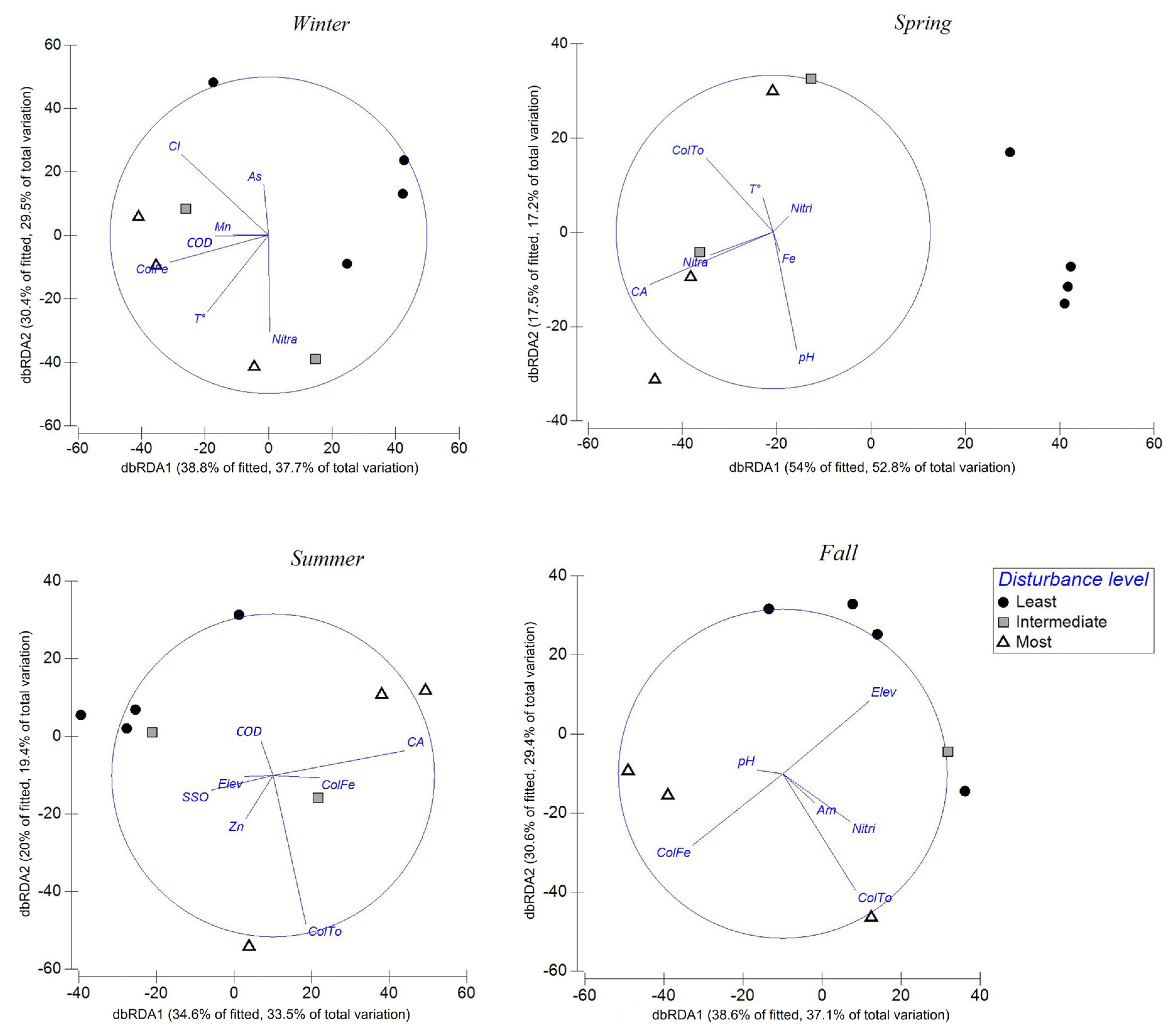
3.3. Multivariate Analysis: Environmental Variables and Macroinvertebrate Assemblages
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| JU | BL 20 | JU 10 | AC 10 | AC 20 | PU 10 | PO 10 | AC 30 | AC 40 | |
|---|---|---|---|---|---|---|---|---|---|
| Temperature (°C) | 6.3 (3–10.3) | 8.5 (4.5–14.9) | 10.2 (6.9–12.6) | 13.4 (10–16.2) | 17 (10.9–21.7) | 15.6 (12.1–17.7) | 15.2 (7.3–22.8) | 16.7 (10.2–20.4) | 14.4 (5.4–18.9) |
| Electrical conductivity (S/cm) | 367.8 (204.2–560) | 720.8 (362–1122) | 303.2 (235.2–375) | 326.5 (257.2–432) | 643 (452–794) | 653.7 (558–721) | 493 (379–585) | 621.3 (594–663) | 938.5 (813–1089) |
| pH | 7.1 (6.3–8) | 6.9 (6–7.5) | 7.4 (6.6–8) | 8 (7.5–8.4) | 7.9 (7–8.6) | 7.6 (7.4–7.8) | 7.5 (6.9–8) | 8.2 (7.9–8.6) | 7.6 (7.3–8.3) |
| Turbidity (NTU) | 32 (11–55) | 36.8 (5–100) | 14.5 (2–47) | 47.8 (5–170) | 50.5 (6–111) | 3 (1–5) | 85.3 (45–132) | 5.3 (4–9) | 16.3 (3–44) |
| Dissolved oxygen (mg/L) | 10.4 (9.6–11.6) | 9.3 (8.4–11.8) | 9.9 (9.5–10.3) | 9.9 (8.8–10.8) | 10.6 (9.6–11.2) | 9.7 (7.9–10.8) | 9.8 (8.5–11.8) | 11.3 (10.4–12) | 10.2 (6–13.2) |
| BDO5 (mg/L) | 1.1 (1–1.3) | 5.8 (2.6–7.8) | 1.1 (1–1.4) | 1 (1–1.1) | 2.9 (2.2–3.4) | 1 (1–1.1) | 2 (1.2–2.6) | 1.4 (1–1.6) | 1.5 (1.1–1.9) |
| COD (mg/L) | 5.6 (2.5–8.5) | 15.5 (9.7–24.1) | 5.5 (2.5–7.6) | 6.3 (2.5–11.5) | 14.7 (3.3–23.9) | 5.2 (2.7–8.5) | 9.8 (4–12.3) | 8.6 (5.4–12.3) | 10.5 (6.8–14.2) |
| Organic suspended solids (mg/L) | 1.6 (1.2–2.5) | 22 (1–3.8) | 2.5 (1–7) | 1.8 (1–3.5) | 7.1 (3.1–11.8) | 1 | 11.8 (6.3–24) | 1.1 (1–1.3) | 2.5 (1–4.2) |
| Inorganic suspended solids (mg/L) | 18.9 (10.1–35.8) | 8 (1–15.7) | 4.7 (1–11.8) | 16.5 (2.6–57.4) | 37.3 (3.1–79.2) | 1 (1–1.1) | 48.3 (31.3–62.8) | 2.2 (1.1–3.9) | 15.2 (2–42.2) |
| Total phosphorus (g/L) | 67.5 (20–120) | 62.5 (10–150) | 35 (10–70) | 132.5 (10–420) | 200 (70–270) | 10 | 187.5 (120–240) | 50 (10–90) | 65 (20–120) |
| Ammonium (g/L) | 20 | 32.5 (20–70) | 25 (20–40) | 20 | 1067.5 (60–1920) | 20 | 142.5 (20–320) | 22.5 (20–30) | 57.5 (30–90) |
| Nitrite (g/L) | 22.5 (15–45) | 133.5 (15–478) | 24.5 (15–53) | 26.3 (15–60) | 852.5 (114–1889) | 33.3 (15–0) | 87.3 (36–174) | 36.5 (15–59) | 153.5 (24–366) |
| Nitrate (mg/L) | 1 (0.9–1.2) | 4.4 (1.4–11.9) | 1.4 (0.9–1.8) | 1.4 (1–2.2) | 3.1 (1.8–5.9) | 17.7 (11.9–23.8) | 2.6 (1.2–4.4) | 11.7 (7.7–17) | 5.7 (0.4–13.7) |
| Al dissolved (g/L) | 55 (13–128) | 18 (5–57) | 76.5 (5–195) | 63 (30–90) | 52.8 (8–140) | 47.7 (27–71) | 47.8 (22–93) | 50.8 (5–81) | 26.8 (5–83) |
| As dissolved (g/L) | 3.5 (1–5) | 3.7 (1.4–7.1) | 4.2 (3–5.7) | 4.3 (2.2–7.5) | 1.8 (1–2.7) | 0.7 (0.2–1) | 3.2 (2.7–3.7) | 1.9 (0.5–3) | 2.3 (1.4–3.5) |
| Cu dissolved (g/L) | 182.4 (0.5–617.4) | 38.2 (13.1–63.8) | 3.5 (0.5–8.9) | 8.6 (0.5–13.8) | 10.7 (5.7–16.2) | 3.7 (0.5–6.9) | 10.2 (4.3–17.5) | 10.2 (4.2–15.3) | 6.5 (0.5–13.8) |
| Fe dissolved (g/L) | 21.5 (3–35) | 16 (1–38) | 30.3 (18–45) | 18.3 (1–27) | 17 (5–36) | 12.7 (1–22) | 28.5 (17–38) | 11.8 (1–32) | 27.5 (5–70) |
| Mn dissolved (g/L) | 27.5 (6–47) | 235.3 (68–570) | 4.5 (1–7) | 36 (4–94) | 30.3 (20–46) | 8 (4–11) | 42.5 (3–129) | 20.3 (5–43) | 31.8 (3–63) |
| Mb dissolved (g/L) | 3.3 (1–5) | 20.3 (1–65) | 2.3 (1–6) | 2.3 (1–5) | 2.5 (1–6) | 1.3 (1–2) | 3.8 (1–7) | 1.8 (1–3) | 1.8 (1–3) |
| Zn dissolved (g/L) | 18.5 (6.7–34.9) | 28–9 (7.2–67.6) | 11.5 (2.5–24.3) | 7.7 (5–13.8) | 13.5 (0.9–34.5) | 9.8 (0.2–22.5) | 10 (5.1–17.1) | 9.9 (3.1–21.9) | 13 (0.2–39.5) |
| Sulfate (mg/L) | 70.7 (51–102) | 186 (93.5–337.6) | 60.4 (50–66.7) | 89.2 (55.4–116.8) | 176.2 (127–234.3) | 167.4 (152.7–176.4) | 161.3 (116.9–223.9) | 155.5 (148.2–161.4) | 248.6 (215.2–266.3) |
| Total hardness (mg/L) | 580.3 (103.6–1875.5) | 231.9 (177–324.1) | 160.4 (125.1–187.1) | 157.9 (106–217.3) | 288.6 (200.4–367) | 372.5 (330.9–404.7) | 233.1 (175.4–315) | 325.8 (279.6–362) | 800.2 (388.3–1961.5) |
| Alkalinity (mg/L) | 52.3 (45–65) | 46 (36–55) | 69.5 (65–73) | 51 (43–56) | 124.5 (95–145) | 137.8 (97–172.5) | 59.4 (50–72.5) | 138.8 (115–155) | 123.4 (52.5–159) |
| Chlorides (mg/L) | 18.3 (10–24.8) | 64.3 (24.1–125.1) | 19.9 (15.9–25.1) | 15.7 (9.1–23.9) | 49.1 (25.8–79.5) | 26.4 (26.2–26.6) | 42.1 (20.6–54.8) | 20.4 (20–20.8) | 53.6 (39.4–66.9) |
| Fecal coliforms (Nmp/100 mL) | 25.1 (4.5–46) | 190.8 (33–330) | 90.2 (7.8–220) | 44 (2–130) | 16.1 (4.5–26) | 1776.6 (240–4600) | 3025 (1100–7900) | 512.5 (170–920) | 892.3 (79–1700) |
| Total coliforms (Nmp/100 mL) | 65.4 (4.5–130) | 1780.8 (33–3300) | 1760 (280–4600) | 116.5 (17–330) | 493.3 (13–1300) | 5410 (330–11,000) | 39475 (7900–79,000) | 493.3 (13–1300) | 5280 (330–17,000) |
References
- Ormerod, S.; Rundle, S.; Wilkinson, S.; Daly, G.; Dale, K.; Juttner, I. Altitudinal trends in the diatoms, bryophytes, macroinvertebrates and fish of a Nepalese river system. Freshw. Biol. 1994, 32, 309–322. [Google Scholar] [CrossRef]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The river continuum concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Fu, L.; Jiang, Y.; Ding, J.; Liu, Q.; Peng, Q.Z.; Kang, M.Y. Impacts of land use and environmental factors on macroinvertebrate functional feeding groups in the Dongjiang River basin, southeast China. J. Freshw. Ecol. 2016, 31, 21–35. [Google Scholar] [CrossRef] [Green Version]
- Allan, J.D. Landscapes and riverscapes: The influence of land use on stream ecosystems. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 257–284. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, J.E.; Lunde, K.B.; Mazor, R.D.; Bêche, L.A.; McElravy, E.P.; Resh, V.H. Long-term macroinvertebrate responses to climate change: Implications for biological assessment in mediterranean-climate streams. J. N. Am. Benthol. Soc. 2010, 29, 1424–1440. [Google Scholar] [CrossRef] [Green Version]
- Fierro, P.; Valdovinos, C.; Arismendi, I.; Díaz, G.; Jara-Flores, A.; Habit, E.; Vargas-Chacoff, L. Examining the influence of human stressors on benthic algae, macroinvertebrate, and fish assemblages in Mediterranean streams of Chile. Sci. Total Environ. 2019, 686, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Martins, F.S.; Formigo, N.; Antunes, S.C. Can be the environmental and biotic factors responsible for macroinvertebrate communities’ alterations in Portuguese alpine ponds? Limnologica 2020, 83, 125782. [Google Scholar] [CrossRef]
- Firmiano, K.R.; Castro, D.M.; Linares, M.S.; Callisto, M. Functional responses of aquatic invertebrates to anthropogenic stressors in riparian zones of Neotropical savanna streams. Sci. Total Environ. 2021, 753, 141865. [Google Scholar] [CrossRef]
- Chen, K.; Hughes, R.M.; Xu, S.; Zhang, J.; Cai, D.; Wang, B. Evaluating performance of macroinvertebrate-based adjusted and unadjusted multi-metric indices (MMI) using multi-season and multi-year samples. Ecol. Indic. 2014, 36, 142–151. [Google Scholar] [CrossRef]
- Miserendino, M.; Brand, C.; Di Prinzio, C. Assessing urban impacts on water quality, benthic communities and fish in streams of the Andes Mountains, Patagonia (Argentina). Water Air Soil Pollut. 2008, 194, 91–110. [Google Scholar] [CrossRef]
- Fierro, P.; Bertrán, C.; Tapia, J.; Hauenstein, E.; Peña-Cortés, F.; Vergara, C.; Cerna, C.; Vargas-Chacoff, L. Effects of local land-use on riparian vegetation, water quality, and the functional organization of macroinvertebrate assemblages. Sci. Total Environ. 2017, 609, 724–734. [Google Scholar] [CrossRef]
- Hunke, P.; Mueller, E.N.; Schröder, B.; Zeilhofer, P. The Brazilian Cerrado: Assessment of water and soil degradation in catchments under intensive agricultural use. Ecohydrology 2015, 8, 1154–1180. [Google Scholar] [CrossRef]
- Brysiewicz, A.; Czerniejewski, P.; Bonisławska, M. Effect of Diverse Abiotic Conditions on the Structure and Biodiversity of Ichthyofauna in Small, Natural Water Bodies Located on Agricultural Lands. Water 2020, 12, 2674. [Google Scholar] [CrossRef]
- Atapattu, S.S.; Kodituwakku, D.C. Agriculture in South Asia and its implications on downstream health and sustainability: A review. Agric. Water Manag. 2009, 96, 361–373. [Google Scholar] [CrossRef]
- Bonada, N.; Prat, N.; Resh, V.H.; Statzner, B. Developments in aquatic insect biomonitoring: A comparative analysis of recent approaches. Annu. Rev. Entomol. 2006, 51, 495–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novoa, V.; Ahumada-Rudolph, R.; Rojas, O.; Munizaga, J.; Sáez, K.; Arumí, J.L. Sustainability assessment of the agricultural water footprint in the Cachapoal River basin, Chile. Ecol. Indic. 2019, 98, 19–28. [Google Scholar] [CrossRef]
- Gerth, W.J.; Li, J.; Giannico, G.R. Agricultural land use and macroinvertebrate assemblages in lowland temporary streams of the Willamette Valley, Oregon, USA. Agric. Ecosyst. Environ. 2017, 236, 154–165. [Google Scholar] [CrossRef] [Green Version]
- Horak, C.N.; Assef, Y.A.; Grech, M.G.; Miserendino, M.L. Agricultural practices alter function and structure of macroinvertebrate communities in Patagonian piedmont streams. Hydrobiologia 2020, 847, 3659–3676. [Google Scholar] [CrossRef]
- Webb, M.J.; Winter, J.M.; Spera, S.A.; Chipman, J.W.; Osterberg, E.C. Water, agriculture, and climate dynamics in central Chile’s Aconcagua River Basin. Phys. Geogr. 2020, 1, 1–21. [Google Scholar] [CrossRef]
- Amigo, J.; Ramírez, C. A bioclimatic classification of Chile: Woodland communities in the temperate zone. Plant Ecol. 1998, 136, 9–26. [Google Scholar] [CrossRef]
- DGA. Diagnóstico y Clasificación de los Cursos y Cuerpos de agua Según Objetivos de Calidad: Cuenca del río Aconcagua; DGA: Valparaiso, Indiana, 2004. [Google Scholar]
- Carter, D.; Aguirre, L. Structural Geology of Aconcagua province and its relationship to the central Valley Graben, Chile. GSA Bull. 1965, 76, 651–664. [Google Scholar] [CrossRef]
- Wilgat, T.; Kosik, A.; Wojcirchowski, K. Hidrología de la cuenca Aconcagua selección de ejemplos. Inv. Geogr. 1968, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Ribbe, L.; Delgado, P.; Salgado, E.; Flügel, W.A. Nitrate pollution of surface water induced by agricultural non-point pollution in the Pocochay watershed, Chile. Desalination 2008, 226, 13–20. [Google Scholar] [CrossRef]
- Gaete, H.; Aránguiz, F.; Cienfuegos, G.; Tejos, M. Metales pesados y toxicidad de aguas del río Aconcagua en Chile. Quim. Nova 2007, 30, 885–891. [Google Scholar] [CrossRef] [Green Version]
- Olivares, Y.; Gaete, H.; Neaman, A. Evaluación de la fitotoxicidad y la genotoxicidad de suelos agrícolas de zonas con actividades mineras de cobre de la cuenca del río Aconcagua (Chile central). Rev. Int. Contam. Ambient. 2015, 31, 237–243. [Google Scholar]
- Hernández, A.; Miranda, M.D.; Arellano, E.C.; Dobbs, C. Landscape trajectories and their effect on fragmentation for a Mediterranean semi-arid ecosystem in Central Chile. J. Arid Environ. 2016, 127, 74–81. [Google Scholar] [CrossRef]
- ESRI. GIS and Mapping Software. 2007. Available online: http://www.esri.com/ (accessed on 9 February 2021).
- Domínguez, E.; Fernández, H.R. Macroinvertebrados bentónicos sudamericanos. Sistemática y biología. Fundación Miguel Lillo, Tucumán, Argentina 2009, 103, 695. [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Wastewater; American Public Health Association (APHA): Washington, DC, USA, 2005. [Google Scholar]
- Elgueta, A.; Górski, K.; Thoms, M.; Fierro, P.; Toledo, B.; Manosalva, A.; Habit, E. Interplay of geomorphology and hydrology drives macroinvertebrate assemblage responses to hydropeaking. Sci. Total Environ. 2021, 768, 144262. [Google Scholar] [CrossRef]
- Anderson, M.J.; Robinson, J. Generalized discriminant analysis based on distances. Aust. NZ J. Stat. 2003, 45, 301–318. [Google Scholar] [CrossRef]
- Anderson, M.; Gorley, R.; Clarke, K.P. For PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Heino, J. Taxonomic surrogacy, numerical resolution and responses of stream macroinvertebrate communities to ecological gradients: Are the inferences transferable among regions? Ecol. Indic. 2014, 36, 186–194. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing [Computer Software Manual]; Team, R.C.: Vienna, Austria, 2016. [Google Scholar]
- Alavaisha, E.; Lyon, S.W.; Lindborg, R. Assessment of water quality across irrigation schemes: A case study of wetland agriculture impacts in Kilombero Valley, Tanzania. Water 2019, 11, 671. [Google Scholar] [CrossRef] [Green Version]
- Burdon, F.J.; Munz, N.A.; Reyes, M.; Focks, A.; Joss, A.; Räsänen, K.; Altermatt, F.; Eggen, R.I.L.; Stamm, C. Agriculture versus wastewater pollution as drivers of macroinvertebrate community structure in streams. Sci. Total Environ. 2019, 659, 1256–1265. [Google Scholar] [CrossRef]
- Usseglio-Polatera, P.; Beisel, J.N. Longitudinal changes in macroinvertebrate assemblages in the Meuse River: Anthropogenic effects versus natural change. River Res. Appl. 2002, 18, 197–211. [Google Scholar] [CrossRef]
- Jacobsen, D.; Wiberg-Larsen, P.; Brodersen, K.P.; Hansen, S.B.; Lindegaard, C.; Friberg, N.; Dall, P.C.; Kirkegaard, J.; Skriver, J.; Toman, M. Macroinvertebrate communities along the main stem and tributaries of a pre-Alpine river: Composition responds to altitude, richness does not. Limnologica 2020, 84, 125816. [Google Scholar] [CrossRef]
- Ruaro, R.; Gubiani, É.A.; Cunico, A.M.; Moretto, Y.; Piana, P.A. Comparison of fish and macroinvertebrates as bioindicators of Neotropical streams. Environ. Monit. Assess. 2016, 188, 45. [Google Scholar] [CrossRef]
- Harrington, R.A.; Poff, N.L.; Kondratieff, B.C. Aquatic insect β-diversity is not dependent on elevation in Southern Rocky Mountain streams. Freshw. Biol. 2016, 61, 195–205. [Google Scholar] [CrossRef]
- Molineri, C.; Nieto, C.; Dos Santos, D.A.; Emmerich, D.; Zúñiga, M.d.C.; Fierro, P.; Pessacq, P.; Gomez, D.; Márquez, J.A.; Príncipe, R.E.; et al. Do mayflies (Ephemeroptera) support a biogeographic transition zone in South America? J. Biogeogr. 2020, 47, 1980–1993. [Google Scholar] [CrossRef]
- Scheibler, E.E.; Fernández Campón, F.; Lagos Silnik, S.; Wellnitz, T. Seasonal influence and local factors affecting macroinvertebrate structure in a high-altitude Andean stream. J. Mt. Sci. 2020, 17, 1374–1386. [Google Scholar] [CrossRef]
- Dos Santos, D.A.; Molineri, C.; Nieto, C.; Zuñiga, M.C.; Emmerich, D.; Fierro, P.; Pessacq, P.; Rios-Touma, B.; Márquez, J.; Gomez, D.; et al. Cold/Warm stenothermic freshwater macroinvertebrates along altitudinal and latitudinal gradients in Western South America: A modern approach to an old hypothesis with updated data. J. Biogeogr. 2018, 45, 1571–1581. [Google Scholar] [CrossRef]
- Gil, M.; Tripole, S.; Vallania, E. Feeding habits of Smicridea (Rhyacophylax) dythyra Flint, 1974 (Trichoptera: Hydropsychidae) larvae in the Los Molles stream (San Luis–Argentina). Acta Limnol. Bras. 2008, 20, 1–4. [Google Scholar]
- Príncipe, R.E.; Gualdoni, C.M.; Raffani, G.B.; Corigliano, M.C. Spatial-temporal patterns of functional feeding groups in mountain streams of Córdoba, Argentina. Ecol. Austral 2010, 20, 257–268. [Google Scholar]
- Fierro, P.; Arismendi, I.; Hughes, R.M.; Valdovinos, C.; Jara-Flores, A. A benthic macroinvertebrate multimetric index for Chilean Mediterranean streams. Ecol. Indic. 2018, 91, 13–23. [Google Scholar] [CrossRef]
- Rodrigues, V.; Estrany, J.; Ranzini, M.; de Cicco, V.; Martín-Benito, J.M.T.; Hedo, J.; Lucas-Borja, M.E. Effects of land use and seasonality on stream water quality in a small tropical catchment: The headwater of Córrego Água Limpa, São Paulo (Brazil). Sci. Total Environ. 2018, 622, 1553–1561. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhang, Y.; Tan, X.; Zheng, Y.; Zhang, Q. Do water quality, land use, or benthic diatoms drive macroinvertebrate functional feeding groups in a subtropical mountain stream? Inland Waters 2020, 1, 1–11. [Google Scholar] [CrossRef]
- Monaghan, K.A.; Peck, M.R.; Brewin, P.; Masiero, M.; Zarate, E.; Turcotte, P.; Ormerod, S. Macroinvertebrate distribution in Ecuadorian hill streams: The effects of altitude and land use. Arch. Hydrobiol. 2000, 149, 421–440. [Google Scholar] [CrossRef]
- Crespo, S.A.; Lavergne, C.; Fernandoy, F.; Muñoz, A.A.; Cara, L.; Olfos-Vargas, S. Where does the chilean aconcagua river come from? Use of natural tracers for water genesis characterization in glacial and periglacial environments. Water 2020, 12, 2630. [Google Scholar] [CrossRef]
- Climent, M.J.; Sánchez-Martín, M.J.; Rodríguez-Cruz, M.S.; Pedreros, P.; Urrutia, R.; Herrero-Hernández, E. Determination of pesticides in river surface waters of central Chile using SPE-GC-MS multi-residue method. J. Chil. Chem. Soc. 2018, 63, 4023–4031. [Google Scholar] [CrossRef] [Green Version]
- Cornejo, A.; Pérez, J.; López-Rojo, N.; Tonin, A.M.; Rovira, D.; Checa, B.; Jaramillo, N.; Correa, K.; Villarreal, A.; Villarreal, V.; et al. Agriculture impairs stream ecosystem functioning in a tropical catchment. Sci. Total Environ. 2020, 745, 140950. [Google Scholar] [CrossRef] [PubMed]
- Shuman, T.C.; Smiley, P.C.; Gillespie, R.B.; Gonzalez, J.M. Influence of Physical and Chemical Characteristics of Sediment on Macroinvertebrate Communities in Agricultural Headwater Streams. Water 2020, 12, 2976. [Google Scholar] [CrossRef]
- Wang, L.; Robertson, D.M.; Garrison, P.J. Linkages between nutrients and assemblages of macroinvertebrates and fish in wadeable streams: Implication to nutrient criteria development. Environ. Manag. 2007, 39, 194–212. [Google Scholar] [CrossRef]
- Heino, J.; Mykrä, H.; Hämäläinen, H.; Aroviita, J.; Muotka, T. Responses of taxonomic distinctness and species diversity indices to anthropogenic impacts and natural environmental gradients in stream macroinvertebrates. Freshw. Biol. 2007, 52, 1846–1861. [Google Scholar] [CrossRef]
- Córdova, S.; Gaete, H.; Aránguiz, F.; Figueroa, R. Evaluación de la calidad de las aguas del estero Limache (Chile central), mediante bioindicadores y bioensayos. Lat. Am. J. Aquat. Res. 2009, 37, 199–209. [Google Scholar] [CrossRef]
- Pizarro, J.; Vergara, P.M.; Rodríguez, J.A.; Valenzuela, A.M. Heavy metals in northern Chilean rivers: Spatial variation and temporal trends. J. Hazard. Mater. 2010, 181, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Suresh, G.; Ramasamy, V.; Meenakshisundaram, V.; Venkatachalapathy, R.; Ponnusamy, V. Influence of mineralogical and heavy metal composition on natural radionuclide concentrations in the river sediments. Appl. Radiat. Isot. 2011, 69, 1466–1474. [Google Scholar] [CrossRef]
- Alvial, I.E.; Orth, K.; Durán, B.C.; Alvarez, E.; Squeo, F.A. Importance of geochemical factors in determining distribution patterns of aquatic invertebrates in mountain streams south of the Atacama Desert, Chile. Hydrobiologia 2013, 709, 11–25. [Google Scholar] [CrossRef]




| Site Code | Elev (m asl) | CA (ha) | Stream Order | Substrate Type | % Agriculture | % Native Forest | % Shrublands | % Forest Plantation | % Urban-and-Industrial | % Water Body | % No Vegetation |
|---|---|---|---|---|---|---|---|---|---|---|---|
| JU | 1657 | 15779 | 3 | Bo/Co | 0.0 | 0.4 | 19.1 | 0.0 | 0.0 | 17.0 | 63.5 |
| BL20 | 1521 | 38729 | 3 | Bo/Co | 0.0 | 1.2 | 10.3 | 0.0 | 4.6 | 25.1 | 58.9 |
| JU10 | 1448 | 17312 | 3 | Bo/Co | 0.0 | 1.2 | 10.3 | 0.0 | 4.6 | 25.1 | 58.9 |
| AC10 | 1096 | 44945 | 4 | Gra/Co | 0.2 | 4.0 | 16.1 | 0.0 | 1.7 | 18.1 | 60.0 |
| AC20 | 650 | 164162 | 5 | Gra/Co | 6.8 | 7.3 | 29.6 | 0.0 | 1.8 | 14.6 | 40.0 |
| PU10 | 604 | 103816 | 4 | Gra | 7.5 | 10.5 | 29.0 | 0.0 | 1.1 | 10.7 | 41.2 |
| PO10 | 600 | 43814 | 4 | Gra/Co | 19.1 | 17.6 | 55.9 | 0.0 | 2.6 | 1.2 | 3.5 |
| AC30 | 188 | 398540 | 5 | Gra | 10.8 | 15.9 | 30.0 | 0.1 | 1.6 | 10.1 | 31.6 |
| AC40 | 12 | 538908 | 5 | Gra | 13.0 | 21.3 | 28.9 | 0.4 | 2.4 | 8.2 | 25.8 |
| Variable | Abbreviation | JU | BL20 | JU 10 | AC 10 | AC 20 | PU 10 | PO 10 | AC 300 | AC 40 |
|---|---|---|---|---|---|---|---|---|---|---|
| Temperature (°C) | T° | 6.3 | 8.5 | 17 | 13.4 | 17.0 | 15.6 | 15.2 | 16.7 | 14.4 |
| Conductivity (µS/cm) | Cond | 367.8 | 720.8 | 303.2 | 326.5 | 643.0 | 653.4 | 493.0 | 621.3 | 938.5 |
| pH | pH | 7.1 | 6.9 | 7.4 | 8.0 | 7.9 | 7.6 | 7.5 | 8.2 | 7.6 |
| Turbidity (NTU) | Tur | 32.0 | 36.8 | 14.5 | 47.8 | 50.5 | 3.0 | 85.3 | 5.3 | 16.3 |
| Dissolved oxygen (mg/L) | DO | 10.4 | 9.3 | 9.9 | 9.9 | 10.6 | 9.7 | 9.8 | 11.3 | 10.2 |
| BDO (mg/L) | BOD | 1.1 | 5.8 | 1.1 | 1.0 | 2.9 | 1.0 | 2.0 | 1.4 | 1.5 |
| COD (mg/L) | COD | 5.6 | 15.5 | 5.5 | 6.3 | 14.7 | 5.2 | 9.8 | 11.3 | 10.2 |
| OSS (mg/L) | SSO | 1.6 | 22.0 | 2.5 | 1.8 | 7.1 | 1.0 | 11.8 | 1.1 | 2.5 |
| ISS (mg/L) | SSI | 18.9 | 8.0 | 4.7 | 16.5 | 37.3 | 1.0 | 48.3 | 2.2 | 15.2 |
| Total phosphorus (µg/L) | TP | 67.5 | 62.5 | 35.0 | 132.5 | 200.0 | 10.0 | 187.5 | 50.0 | 65.0 |
| Ammonium (µg/L) | Am | 20.0 | 32.5 | 25.0 | 20.0 | 1067.5 | 20.0 | 142.5 | 22.5 | 57.5 |
| Nitrite (µg/L) | Nitri | 22.5 | 133.5 | 24.5 | 26.3 | 852.5 | 33.3 | 87.3 | 36.5 | 153.5 |
| Nitrate (mg/L) | Nitra | 1.0 | 4.4 | 1.4 | 1.4 | 3.1 | 17.7 | 2.6 | 11.7 | 5.7 |
| Al dissolved (µg/L) | Al | 55.0 | 18.0 | 76.5 | 63.0 | 52.8 | 47.7 | 47.8 | 50.8 | 26.8 |
| As dissolved (µg/L) | As | 3.5 | 3.7 | 4.2 | 4.3 | 1.8 | 0.7 | 3.2 | 1.9 | 2.3 |
| Cu dissolved (µg/L) | Cu | 182.4 | 38.2 | 3.5 | 8.6 | 10.7 | 3.7 | 10.2 | 10.2 | 6.5 |
| Fe dissolved (µg/L) | Fe | 21.5 | 16.0 | 30.3 | 18.3 | 17.0 | 12.7 | 28.5 | 11.8 | 27.5 |
| Mn dissolved (µg/L) | Mn | 27.5 | 235.3 | 4.5 | 36.0 | 30.3 | 8.0 | 42.5 | 20.3 | 31.8 |
| Mb dissolved (µg/L) | Mb | 3.3 | 20.3 | 2.3 | 2.3 | 2.5 | 1.3 | 3.8 | 1.8 | 1.8 |
| Zn dissolved (µg/L) | Zn | 18.5 | 28.9 | 11.5 | 7.7 | 13.5 | 9.8 | 10.0 | 9.9 | 13.0 |
| Sulfate (mg/L) | Sulf | 70.7 | 186.0 | 60.4 | 89.2 | 176.2 | 167.4 | 161.3 | 155.5 | 248.6 |
| Total hardness (mg/L) | TH | 580.3 | 231.9 | 160.4 | 157.9 | 288.6 | 372.5 | 233.1 | 225.8 | 800.2 |
| Alkalinity (mg/L) | Alk | 52.3 | 46.0 | 69.5 | 51.0 | 124.5 | 137.8 | 59.4 | 138.8 | 123.4 |
| Chlorides (mg/L) | Cl | 18.3 | 64.3 | 19.9 | 15.7 | 49.1 | 26.4 | 42.1 | 20.4 | 53.6 |
| Fecal coliforms (Nmp/100 mL) | ColFe | 25.1 | 190.8 | 90.2 | 44.0 | 16.1 | 1776.6 | 3025 | 512.5 | 892.3 |
| Total coliforms (Nmp/100 mL) | ColTo | 65.4 | 1780.8 | 1760 | 116.5 | 493.3 | 5410.0 | 39,475.0 | 493.3 | 5280.0 |
| Percentage Correct Classifications | Among Land Use Types Differences | ||||
|---|---|---|---|---|---|
| CAP | Least | Intermediate | Most | Trace | P |
| Environmental variables | |||||
| Winter | 100 | 100 | 66.67 | 0.587 | 0.064 |
| Spring | 100 | 100 | 66.67 | 0.566 | 0.055 |
| Summer | 100 | 100 | 66.67 | 0.566 | 0.057 |
| Fall | 100 | 100 | 66.67 | 0.583 | 0.079 |
| Macroinvertebrate assemblages | |||||
| Winter | 50 | 0 | 100 | 1.444 | 0.080 |
| Spring | 100 | 100 | 100 | 1.943 | 0.002 |
| Summer | 100 | 0 | 66.67 | 1.490 | 0.008 |
| Fall | 75 | 0 | 66.67 | 1.216 | 0.094 |
| Variable | Adjust.R | Pseudo-F | P | Cumulative Proportion |
|---|---|---|---|---|
| Winter | ||||
| CDOM | 0.289 | 2.856 | 0.013 | 0.289 |
| Temperature | 0.528 | 3.029 | 0.012 | 0.528 |
| Fecal coliforms | 0.657 | 1.884 | 0.052 | 0.657 |
| As | 0.75 | 1.646 | 0.13 | 0.757 |
| Cl | 0.843 | 1.651 | 0.211 | 0.843 |
| Nitrates | 0.911 | 1.546 | 0.267 | 0.912 |
| Mn | 0.969 | 1.902 | 0.322 | 0.970 |
| Spring | ||||
| Temperature | 0.425 | 5.174 | 0.005 | 0.425 |
| pH | 0.589 | 2.401 | 0.018 | 0.589 |
| Nitrates | 0.697 | 1.776 | 0.139 | 0.697 |
| Catchment area | 0.793 | 1.868 | 0.152 | 0.793 |
| Total coliforms | 0.857 | 1.971 | 0.144 | 0.875 |
| Fe | 0.931 | 1.634 | 0.274 | 0.931 |
| Nitrites | 0.977 | 2.091 | 0.33 | 0.977 |
| Summer | ||||
| Elevation | 0.268 | 2.569 | 0.01 | 0.268 |
| Total coliforms | 0.443 | 1.888 | 0.024 | 0.443 |
| ColFe | 0.586 | 1.726 | 0.08 | 0.586 |
| SSO | 0.706 | 1.632 | 0.171 | 0.706 |
| Catchment area | 0.812 | 1.704 | 0.176 | 0.812 |
| Zn | 0.891 | 1.439 | 0.301 | 0.891 |
| COD | 0.97 | 2.635 | 0.277 | 0.97 |
| Fall | ||||
| Elevation | 0.2810 | 2.3453 | 0.0210 | 0.2810 |
| Total coliforms | 0.5211 | 2.5050 | 0.0310 | 0.5211 |
| Nitrites | 0.7083 | 2.5670 | 0.0410 | 0.7083 |
| pH | 0.8193 | 1.8430 | 0.1220 | 0.8193 |
| Ammonium | 0.8982 | 1.5480 | 0.2620 | 0.8982 |
| Fecal coliforms | 0.9599 | 1.5400 | 0.4050 | 0.9599 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fierro, P.; Valdovinos, C.; Lara, C.; Saldías, G.S. Influence of Intensive Agriculture on Benthic Macroinvertebrate Assemblages and Water Quality in the Aconcagua River Basin (Central Chile). Water 2021, 13, 492. https://doi.org/10.3390/w13040492
Fierro P, Valdovinos C, Lara C, Saldías GS. Influence of Intensive Agriculture on Benthic Macroinvertebrate Assemblages and Water Quality in the Aconcagua River Basin (Central Chile). Water. 2021; 13(4):492. https://doi.org/10.3390/w13040492
Chicago/Turabian StyleFierro, Pablo, Claudio Valdovinos, Carlos Lara, and Gonzalo S. Saldías. 2021. "Influence of Intensive Agriculture on Benthic Macroinvertebrate Assemblages and Water Quality in the Aconcagua River Basin (Central Chile)" Water 13, no. 4: 492. https://doi.org/10.3390/w13040492
APA StyleFierro, P., Valdovinos, C., Lara, C., & Saldías, G. S. (2021). Influence of Intensive Agriculture on Benthic Macroinvertebrate Assemblages and Water Quality in the Aconcagua River Basin (Central Chile). Water, 13(4), 492. https://doi.org/10.3390/w13040492