
Assessing the Ecotoxicity of Copper and Polycyclic Aromatic Hydrocarbons: Comparison of Effects on Paracentrotus lividus and Botryllus schlosseri, as Alternative Bioassay Methods





Abstract
:
1. Introduction
2. Materials and Methods
2.1. Ethics Statement for Paracentrotus lividus and Botryllus schlosseri
2.2. Experimental Procedures for P. lividus
2.3. Experimental procedures for Botryllus schlosseri
2.4. Collection Procedures
2.5. Toxicity Tests on Tunicates
2.6. Statistical Analyses
3. Results
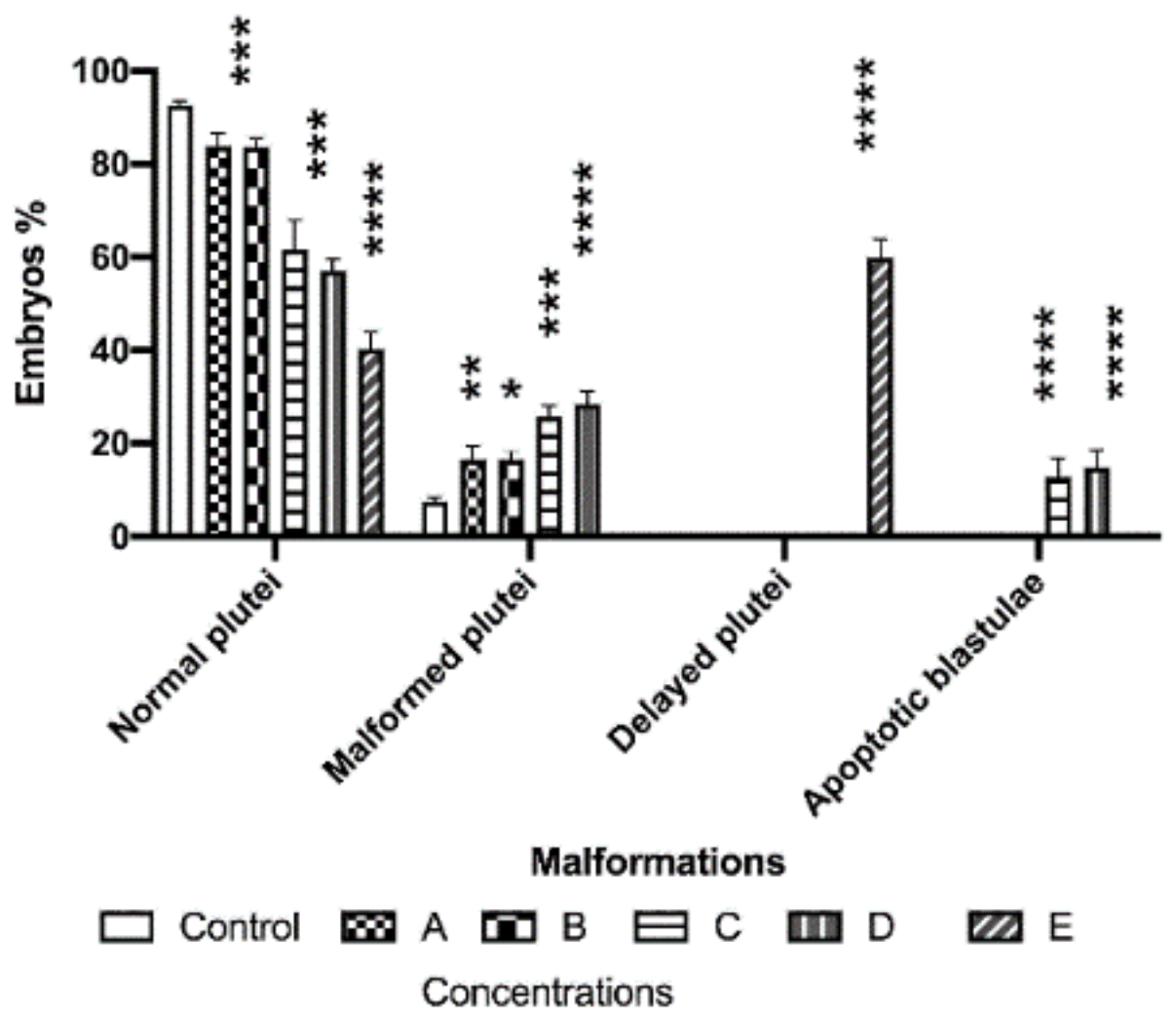
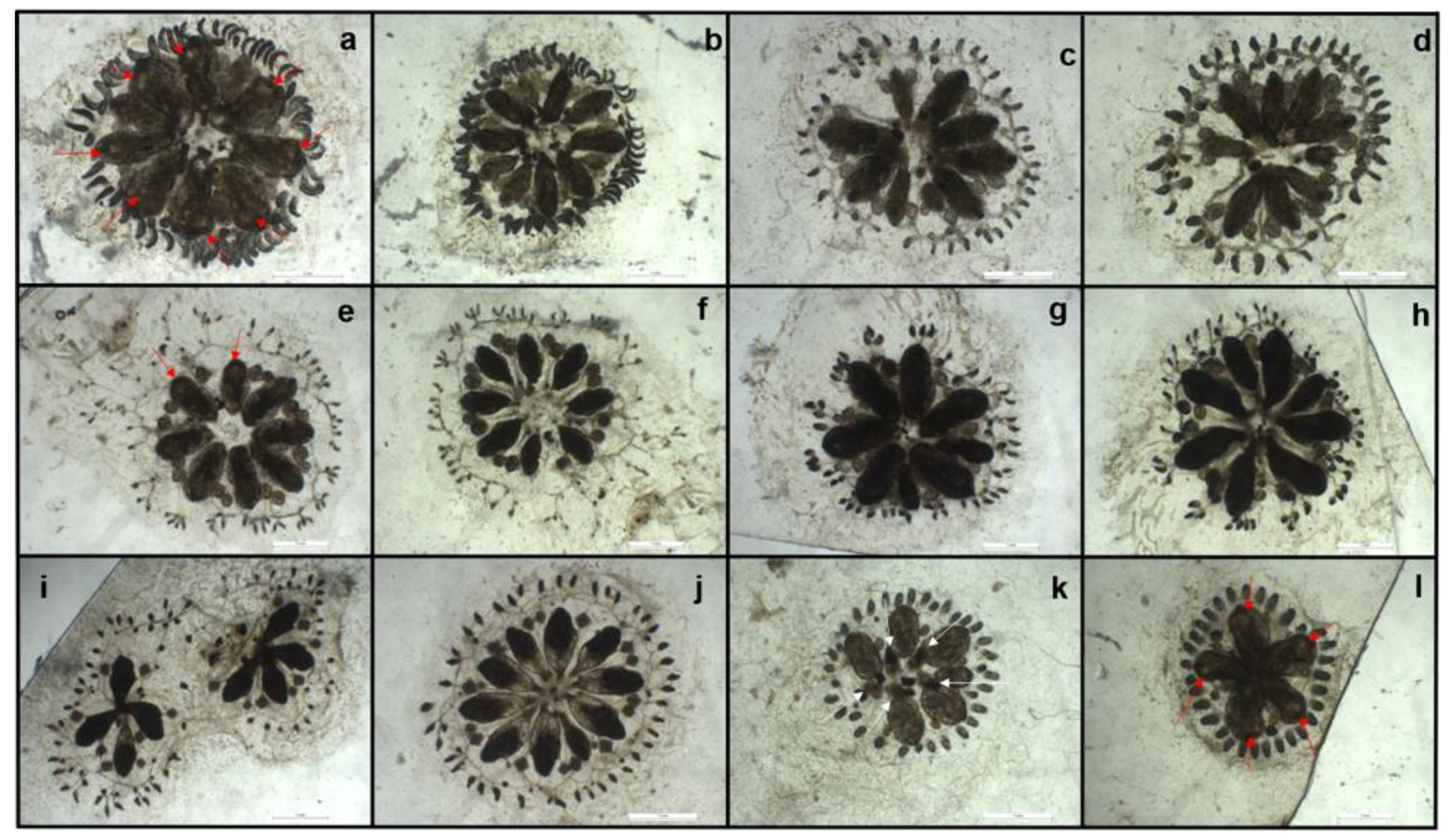
3.1. Morphological Effects of PAHs and Cu on Sea Urchin Embryos
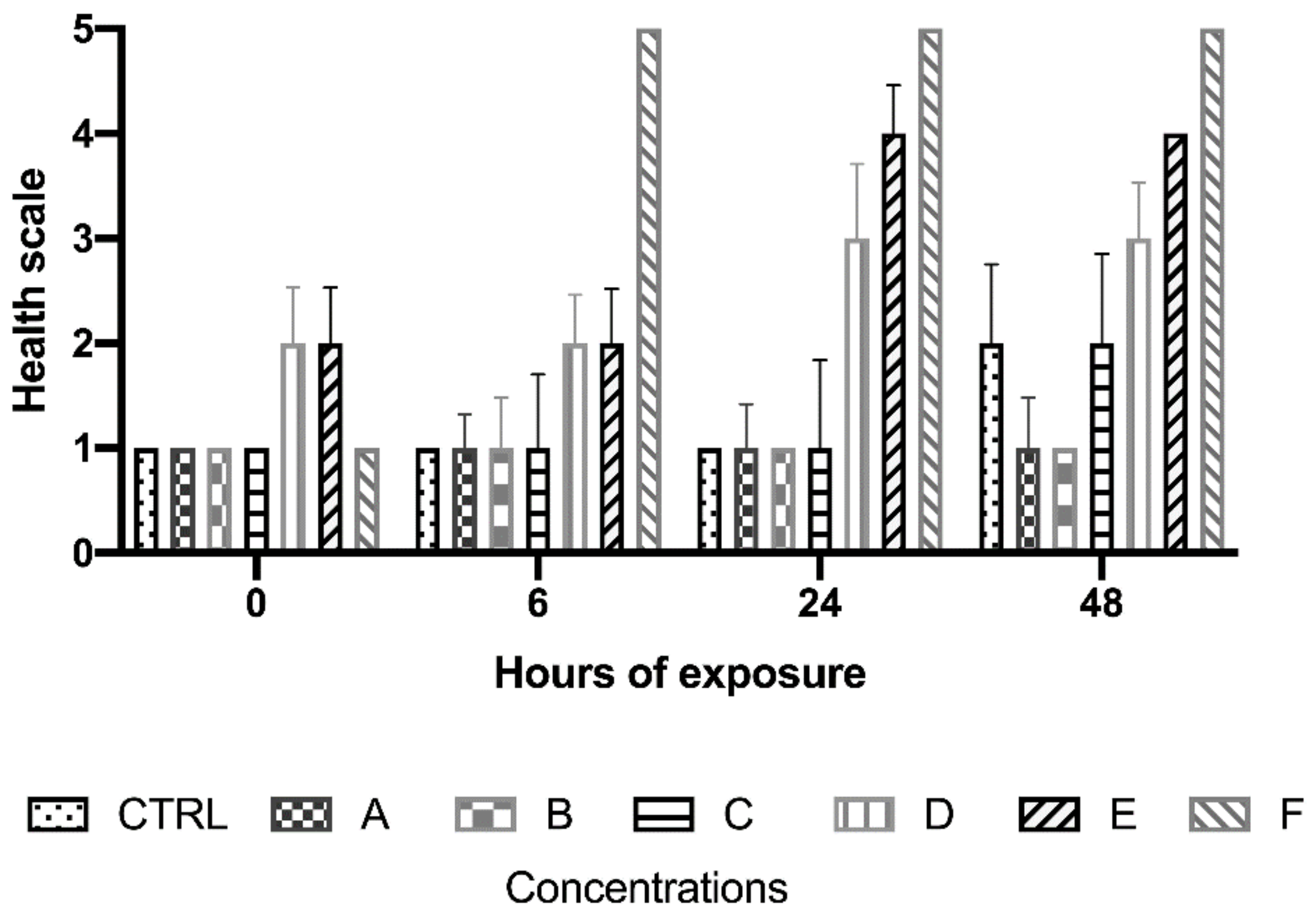
3.2. Morphological Effects of PAHs and Cu on Botryllus shlosseri
3.2.1. Chemical Descriptors of Media
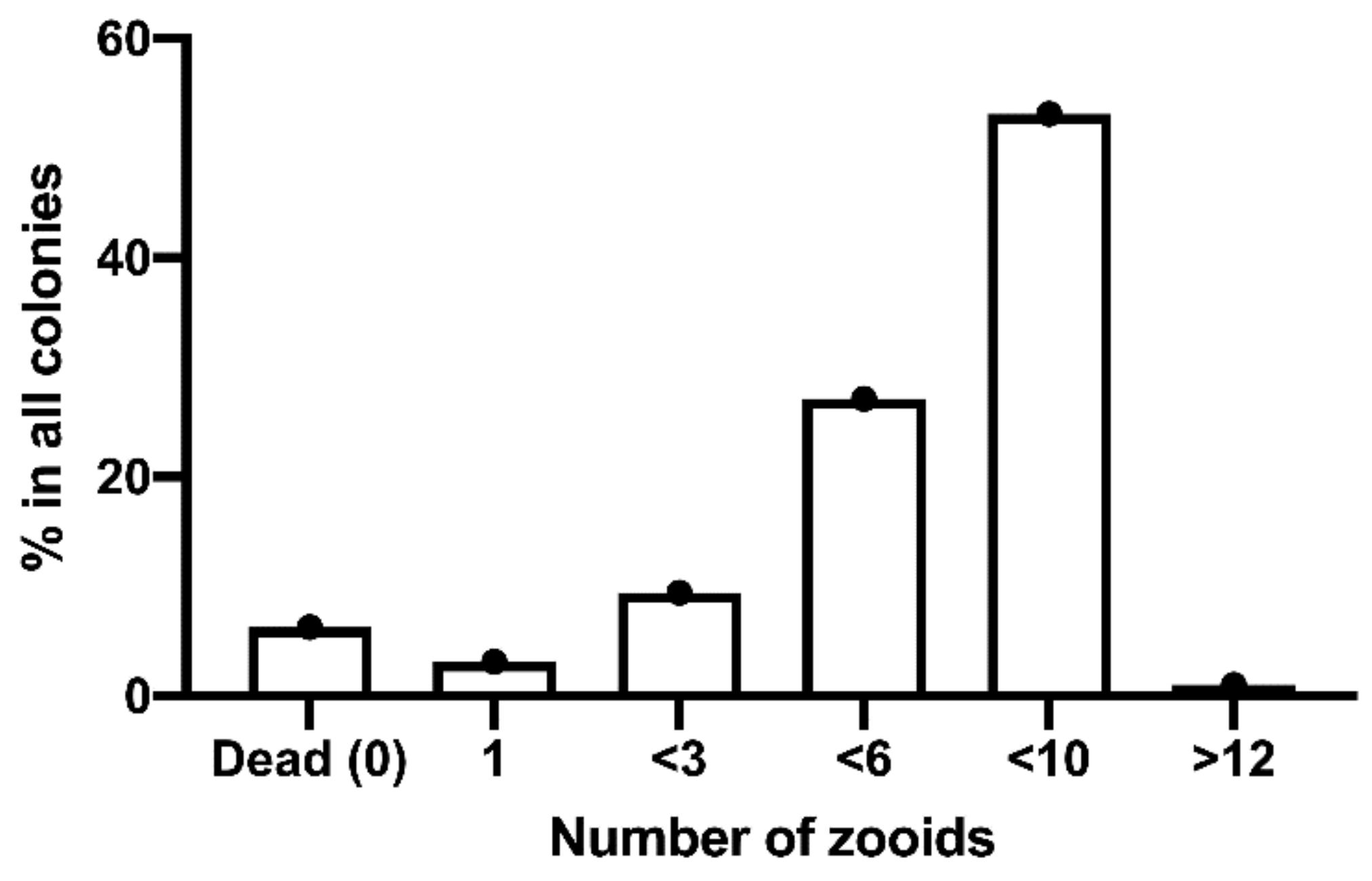
3.2.2. Development Times of Clutches
3.2.3. Effects of PAHs
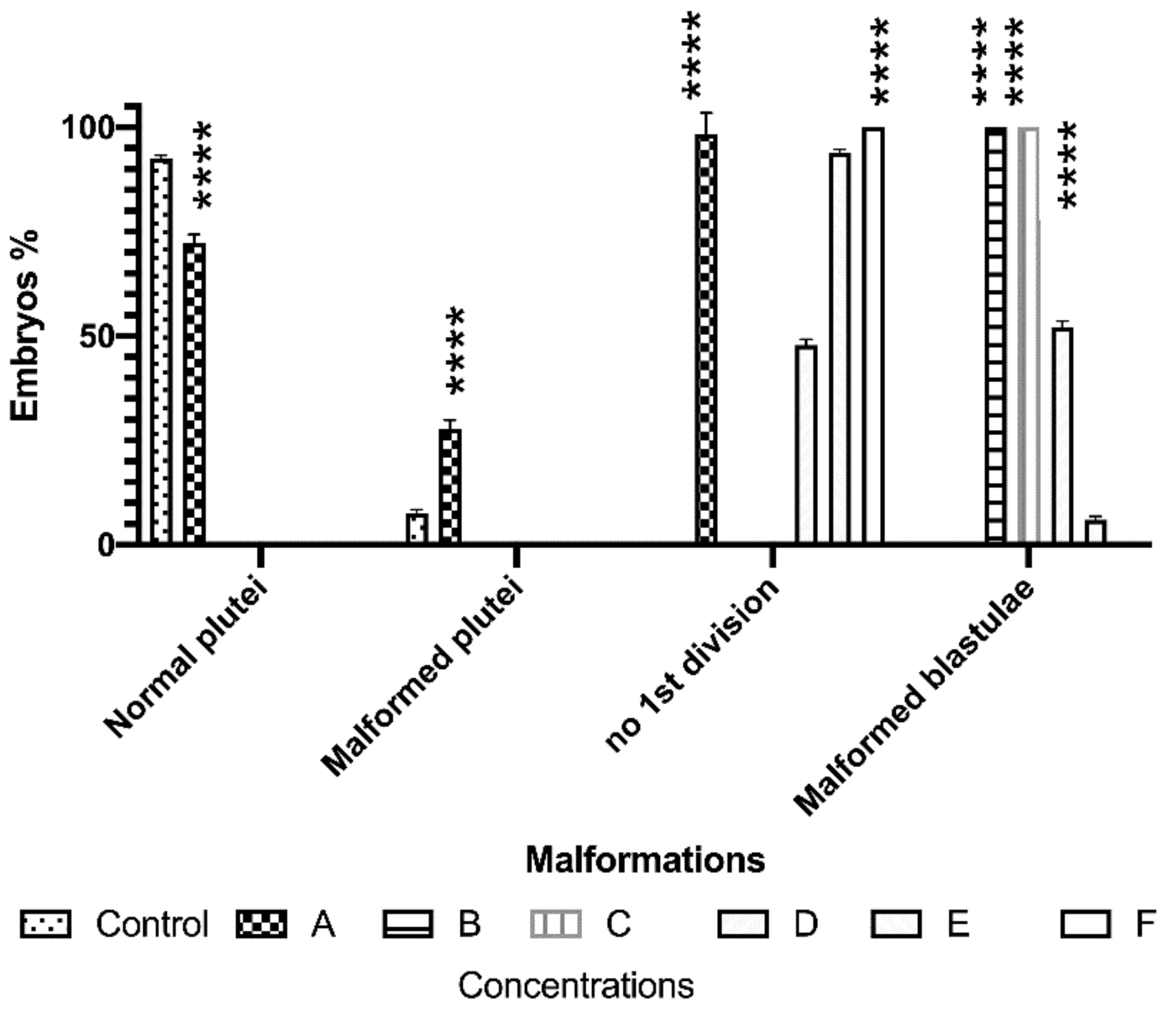
3.2.4. Effect of Copper
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sánchez, A.; Recillas, S.; Font, X.; Casals, E.; González, E.; Puntes, V. Ecotoxicity of, and remediation with, engineered inorganic nanoparticles in the environment. Trends Anal. Chem. 2011, 30, 507–516. [Google Scholar] [CrossRef] [Green Version]
- Zheng, N.; Wang, S.; Dong, W.; Hua, X.; Li, Y.; Song, X.; Chu, Q.; Hou, S.; Li, Y.Z.H.; Shi, J.; et al. The toxicological effects of mercury exposure in marine fish. Bull. Environ. Contam. Toxicol. 2019, 102, 714–720. [Google Scholar] [CrossRef]
- Dahms, H.-U.; Won, E.-J.; Kim, H.-S.; Han, J.; Park, H.G.; Souissi, S.; Raisuddin, S.; Lee, J.-S. Potential of the small cyclopoid copepod Paracyclopina nana as an invertebrate model for ecotoxicity testing. Aquat. Toxicol. 2016, 180, 282–294. [Google Scholar] [CrossRef]
- Qu, C.; Li, J.; Albanese, S.; Lima, A.; Wang, M.; Sacchi, M.; Molisso, F.; De Vivo, B. Polycyclic aromatic hydrocarbons in the sediments of the Gulfs of Naples and Salerno, Southern Italy: Status, sources and ecological risk. Ecotoxicol. Environ. Saf. 2018, 161, 156–163. [Google Scholar] [CrossRef]
- Ruocco, N.; Bertocci, I.; Munari, M.; Musco, M.; Caramiello, D.; Danovaro, R.; Zupo, V.; Costantini, M. Morphological and molecular responses of the sea urchin Paracentrotus lividus to highly contaminated marine sediments: The case study of Bagnoli-Coroglio brownfield (Mediterranean Sea). Mar. Environ. Res. 2020, 154, 104–165. [Google Scholar] [CrossRef] [PubMed]
- Kronenberg, M.; Trably, E.; Bernet, N.; Patureau, D. Biodegradation of polycyclic aromatic hydrocarbons: Using microbial bioelectrochemical systems to overcome an impasse. Environ. Pollut. 2017, 231, 509–523. [Google Scholar] [CrossRef] [PubMed]
- Yim, U.H.; Hong, S.H.; Shim, W.J.; Oh, J.R.; Chang, M. Spatio-temporal distribution and characteristics of PAHs in sediments from Masan Bay, Korea. Mar. Pollut. Bull. 2005, 50, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Myers, R.A.; Worm, B. Rapid worldwide depletion of predatory fish communities. Nature 2003, 423, 280–283. [Google Scholar] [CrossRef] [PubMed]
- Lotze, H.K.; Lenihan, H.S.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.G.; Kay, M.C.; Kidwell, S.M.; Kirby, S.M.; Peterson, C.H.; Jackson, J.B.C. Depletion, degradation and recovery potential of estuaries and coastal seas. Science 2006, 312, 1806–1809. [Google Scholar] [CrossRef]
- Halpern, B.S.; Walbridge, S.; Selkoe, K.A.; Kappel, C.V.; Micheli, F.; D’Agrosa, C.; Bruno, J.F.; Casey, K.S.; Ebert, C.; Fox, H.E.; et al. A global map of human impact on marine ecosystems. Science 2008, 319, 948–952. [Google Scholar] [CrossRef] [Green Version]
- Romano, E.; Bergamin, L.; Magno, M.C.; Pierfranceschi, G.; Ausili, A. Temporal changes of metal and trace element contamination in marine sediments due to a steel plant: The case study of Bagnoli (Naples, Italy). Appl. Geochem. 2017, 88, 85–94. [Google Scholar] [CrossRef]
- Cooper, E.L.; Arizza, V.; Cammarata, M.; Pellerito, L.; Parrinello, N. Tributyltin affects phagocytic activity of Ciona intestinalis hemocytes. Comp. Biochem. Physiol. 1995, 112, 285–289. [Google Scholar] [CrossRef]
- Cima, F.; Ballarin, L. Effects of antifouling paints alternative to organotin-based ones on macrofouling biocoenoses of hard substrates in the Lagoon of Venice. Fresen. Environ. Bull. 2008, 17, 1901–1908. [Google Scholar]
- Epelbaum, A.; Therriault, T.W.; Paulson, A.; Pearce, C.M. Botryllid tunicates: Culture techniques and experimental procedures. Aquat. Invasions 2009, 4, 111–120. [Google Scholar] [CrossRef]
- Gao, Y.; Li, S.; Zhan, A. Genome-wide single nucleotide polymorphisms (SNPs) for a model invasive ascidian Botryllus schlosseri. Genetica 2018, 146, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Epelbaum, A.; Herborg, L.M.; Therriault, T.W.; Pearce, C.M. Temperature and salinity effects on growth, survival, reproduction, and potential distribution of two non-indigenous botryllid ascidian in British Columbia. J. Exp. Mar. Biol. Ecol. 2009, 369, 43–52. [Google Scholar] [CrossRef]
- Cima, F.; Ballarin, L. A proposed integrated bioindex for the macrofouling biocoenosis of hard substrata in the lagoon of Venice. Estuar. Coast. Shelf Sci. 2013, 130, 190–201. [Google Scholar] [CrossRef]
- Reem, E.; Douek, J.; Paz, G.; Katzir, G.; Rinkevich, B. Phylogenetics, biogeography and population genetics of the ascidian Botryllus schlosseri in the Mediterranean Sea and beyond. Mol. Phylogenetics Evol. 2017, 107, 221–231. [Google Scholar] [CrossRef]
- Gittenberger, A.; Simeon Moons, J.J. Settlement and possible competition for space between the invasive violet tunicate Botrylloides violaceus and the native star tunicate Botryllus schlosseri in the Netherlands. Aquat. Invasions 2011, 6, 435–440. [Google Scholar] [CrossRef]
- Zega, G.; Pennati, R.; Candiani, S.; Pestarino, M.; De Bernardi, F. Solitary ascidians embryos (Chordata, Tunicata) as model organisms for testing coastal pollutant toxicity. Invertebr. Surviv. J. 2009, 6, S29–S34. [Google Scholar]
- Manni, L.; Gasparini, F.; Hotta, K.; Ishizuka, K.J.; Ricci, L.; Tiozzo, S.; Voskoboynik, A.; Dauga, D. Ontology for the Asexual Development and Anatomy of the Colonial Chordate Botryllus schlosseri. PLoS ONE 2014, 9, e96434. [Google Scholar] [CrossRef] [Green Version]
- Manni, L.; Burighel, P. Common and divergent pathways in alternative developmental processes of ascidians. BioEssays 2006, 28, 902–912. [Google Scholar] [CrossRef] [PubMed]
- Delsuc, F.; Brinkmann, H.; Chourrout, D.; Philippe, H. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 2006, 439, 965–968. [Google Scholar] [CrossRef] [PubMed]
- Voskoboynik, A.; Weissman, I.L. Botryllus schlosseri, an emerging model for the study of aging, stem cells, and mechanisms of regeneration. Invert. Reprod. Dev. 2015, 59, 33–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, C.C.; Lambert, G. Persistence and differential distribution of nonindigenous ascidians in harbours of the Southern California Bight. Mar. Ecol. Prog. Ser. 2003, 259, 145–161. [Google Scholar] [CrossRef] [Green Version]
- Carver, C.E.; Mallet, A.L.; Vercaemer, B. Biological synopsis of the solitary tunicate Ciona intestinalis. Can. Man. Rep. Fish. Aquat. Sci. 2006, 2746, 1–52. [Google Scholar]
- Naranjo, S.A.; Carballo, J.L.; Garcia-Gomez, J.C. Effects of environmental stress on ascidian populations in Algeciras Bay (southern Spain). Possible marine bioindicators? Mar. Ecol. Prog. Ser. 1996, 144, 119–131. [Google Scholar] [CrossRef] [Green Version]
- Brunetti, R.; Beghi, L.; Bressan, M.; Marin, M.G. Combined Effects of Temperature and Salinity on Colonies of Botryllus schlosseri and Botrylloides leachi (Ascidiacea) from the Venetian Lagoon. Mar. Ecol. Prog. Ser. 1980, 2, 303–314. [Google Scholar] [CrossRef]
- Bamforth, S.M.; Singleton, L. Bioremediation of polycyclicaromatic hydrocarbons, current knowledge and future directions. J. Chem. Technol. Biotechnol. 2005, 80, 723–736. [Google Scholar] [CrossRef]
- Balcioğlu, E.B. Potential effects of polycyclic aromatic hydrocarbons (PAHs) in marine foods on human health: A critical review. Tox. Rev. 2016, 35, 98–105. [Google Scholar] [CrossRef]
- Hylland, K. Polycyclic Aromatic Hydrocarbon (PAH) Ecotoxicology in Marine Ecosystems. J. Toxicol. Environ. Health Part A 2006, 69, 109–123. [Google Scholar] [CrossRef]
- Yu, W.; Liu, R.; Xu, F.; Shen, Z. Environmental risk assessments and spatial variations of polycyclic aromatic hydrocarbons in surface sediments in Yangtze River Estuary, China. Mar. Pollut. Bull. 2015, 100, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Albarano, L.; Lofrano, G.; Costantini, M.; Zupo, V.; Carraturo, F.; Guida, M.; Libralato, G. Comparison of in situ sediment remediation amendments: Risk perspectives from species sensitivity distribution. Environ. Pollut. 2021, 272, 115995. [Google Scholar] [CrossRef]
- Osborne, K.L.; Poynton, H.C. Copper pollution enhances the competitive advantage of invasive ascidians. Manag. Biol. Invasions 2019, 10, 641–656. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, D.H.; Sandhal, J.F.; Labenia, J.S.; Scholz, N.L. Sublethal effects of copper on Coho salmon: Impacts on nonoverlapping receptor pathways in the peripheral olfactory nervous system. Environ. Toxicol. Chem. 2003, 22, 2266–2274. [Google Scholar] [CrossRef] [PubMed]
- Gissi, F.; Stauber, J.; Reichelt-Brushett, A.; Harrison, P.L.; Jolley, D.F. Inhibition in fertilisation of coral gametes following exposure to nickel and copper. Ecotox. Environ. Saf. 2017, 145, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Maibam, C.; Fink, P.; Romano, G.; Buia, M.C.; Gambi, M.C.; Scipione, M.B.; Patti, F.P.; Lorenti, M.; Butera, E.; Zupo, V. Relevance of wound-activated compounds produced by diatoms as toxins and infochemicals for benthic invertebrates. Mar. Biol. 2014, 161, 1639–1652. [Google Scholar] [CrossRef]
- Lewis, C.; Ellis, R.; Vernon, E.; Elliot, K.; Newbatt, S.; Wilson, R.W. Ocean acidification increases copper toxicity differentially in two key marine invertebrates with distinct acid-base responses. Sci. Rep. 2016, 6, 21554. [Google Scholar] [CrossRef] [Green Version]
- Toussaint, M.W.; Shedd, T.R.; Van Der Schalie, W.H.; Leather, G.R. A comparison of standard acute toxicity tests with rapid-screening toxicity tests. Environ. Toxicol. Chem. 1995, 14, 907–915. [Google Scholar] [CrossRef]
- Morroni, L.; Pinsino, A.; Pellegrini, D.; Regoli, F.; Matranga, V. Development of a new integrative toxicity index based on an improvement of the sea urchin embryo toxicity test. Ecotoxicol. Environ. Saf. 2016, 123, 2–7. [Google Scholar] [CrossRef]
- Morroni, L.; Giuliani, S.; Pellegrini, D.; Sartori, D. In situ embryo toxicity test with sea urchin: Development of exposure chamber for test execution. Chemosphere 2017, 196, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Gregorin, C.; Musco, L.; Somma, E.; Zupo, V. Behavioural Responses of the Colonial Sea Squirt Botrylloides violaceus Oka to Suspended Food Micro-Particles in Laboratory Cultures. J. Mar. Sci. Eng. 2020, 8, 1021. [Google Scholar] [CrossRef]
- Suzuki, N.; Ogiso, S.; Yachiguchi, K.; Kawabe, K.; Makino, F.; Toriba, A.; Kiyomoto, M.; Sekiguchi, T.; Tabuchi, Y.; Kondo, T.; et al. Monohydroxylated polycyclic aromatic hydrocarbons influence spicule formation in the early development of sea urchins (Hemicentrotus pulcherrimus). Comp. Biochem. Physiol. C 2015, 171, 55–60. [Google Scholar] [CrossRef]
- Suzuki, T.; Sakata, K.; Mizuno, N.; Palikhe, S.; Yamashita, S.; Hattori, K.; Matsuda, N.; Hattori, Y. Different involvement of the MAPK family in inflammatory regulation in human pulmonary microvascular endothelial cells stimulated with LPS and IFN-γ. Immunobiology 2018, 223, 777–785. [Google Scholar] [CrossRef]
- Pillai, M.C.; Vines, C.A.; Wikramanayake, A.H.; Cherr, G.N. Polycyclic aromatic hydrocarbons disrupt axial development in sea urchin embryos through a beta-catenin dependent pathway. Toxicology 2003, 186, 93–108. [Google Scholar] [CrossRef]
- Kobayashi, N.; Okamura, H. Effects of heavy metals on sea urchin embryo development. Part 2. Interactive toxic effects of heavy metals in synthetic mine effluents. Chemosphere 2005, 61, 1198–1203. [Google Scholar] [CrossRef] [PubMed]
- Bielmyer, G.K.; Brix, K.V.; Tom, T.C.; Capop, T.; Grosell, M. The effects of metals on embryo-larval and adult life stages of the sea urchin, Diadema antillarum. Aquat. Toxicol. 2005, 74, 254–263. [Google Scholar] [CrossRef]
- His, E.; Heyvang, I.; Geffard, O.; de Montaudouin, X.A. comparison between oyster (Crassostrea gigas) and sea urchin (Paracentrotus lividus) larval bioassays for toxicological studies. Water Res. 1999, 33, 1706–1718. [Google Scholar] [CrossRef]
- Lotufo, G.R.; Farrara, J.D.; Biedenbach, J.M.; Lairda, J.G.; Krasnec, M.O.; Lay, C.; Morris, J.M.; Gielazync, M.L. Effects of sediment amended with Deepwater Horizon incident slick oil on the infaunal amphipod Leptocheirus plumulosus. Mar. Pollut. Bull. 2016, 109, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Cima, F.; Ballarin, L. Immunotoxicity in ascidians: Antifouling compounds alternative to organotins III—The case of copper(I) and Irgarol 1051. Chemosphere 2012, 89, 19–29. [Google Scholar] [CrossRef]
- Cima, F.; Bragadin, M.; Ballarin, L. Toxic effects of new antifouling compounds on tunicate haemocytes I. Sea-Nine 211TM and chlorothalonil. Aquat. Toxicol. 2008, 86, 299–312. [Google Scholar] [CrossRef]
- Menin, A.; Ballarin, L.; Bragadin, M.; Cima, F. Immunotoxicity in ascidians: Antifouling compounds alternative to organotins—II. the case of Diuron and TCMS pyridine. J. Environ. Sci. Health 2008, 43B, 644–654. [Google Scholar] [CrossRef]
- Cima, F.; Ballarin, L.; Caicci, F.; Franchi, N.; Gasparini, F.; Rigon, F.; Schiavon, F.; Manni, L. Life history and ecological genetics of the colonial ascidian Botryllus schlosseri. Zool. Anz. 2015, 257, 54–70. [Google Scholar] [CrossRef] [Green Version]
- Kamer, I.; Rinkevich, B. In vitro application of the comet assay for aquatic genotoxicity: Considering a primary culture versus a cell line. Toxicol Vitr. 2002, 16, 177–184. [Google Scholar] [CrossRef]
- Boudouresque, C.F.; Verlaque, M. Ecology of Paracentrotus lividus. In Edible Sea Urchins: Biology and Ecology; Lawrence, J.M., Ed.; Elsevier: Amsterdam, The Netherlands, 2001; pp. 177–216. [Google Scholar]
- Bellas, J.; Beiras, R.; Vazquez, E. A standardisation of Ciona intestinalis (Chordata, Ascidiacea) embryo-larval bioassay for ecotoxicological studies. Water Res. 2003, 37, 4613–4622. [Google Scholar] [CrossRef]
- Bellas, J.; Fernandez, N.; Lorenzo, I.; Beiras, R. Integrative assessment of coastal pollution in a Ria coastal system (Galicia, NW Spain): Correspondence between sediment chemistry and toxicity. Chemosphere 2008, 72, 826–835. [Google Scholar] [CrossRef]
- Bellas, J.; Thor, P. Effects of selected PAHs on reproduction and survival of the calanoid copepod Acartia tonsa. Ecotoxicology 2007, 16, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Lorenti, M.; Gambi, M.C.; Guglielmo, R.; Patti, F.P.; Scipione, M.B.; Zupo, V.; Buia, M.C. Soft-bottom macrofaunal assemblages in the Gulf of Salerno, Tyrrhenian Sea, Italy, an area affected by the invasion of the seaweed Caulerpa racemosa var. cylindracea. Mar. Ecol. 2011, 32, 320–334. [Google Scholar] [CrossRef]
- Piola, R.F.; Johnston, E.L. Differential tolerance to metals among populations of the introduced bryozoan Bugula neritina. Mar. Biol. 2006, 148, 997–1010. [Google Scholar] [CrossRef]
- Zupo, V.; Fresi, E. A study on the food web of the Posidonia oceanica (L.) Delile ecosystem: Analysis of the gut contents of decapod crustaceans. Rapp. Comm. Int. Mer Medit. 1985, 29, 189–192. [Google Scholar]
- Ruiz, G.M.; Carlton, J.T.; Grosholz, E.D.; Hines, A.H. Global invasions of marine and estuarine habitats by non-indigenous species: Mechanisms, extent, and consequences. Am. Zool. 1997, 37, 621–632. [Google Scholar] [CrossRef]
- Ruiz, G.M.; Fofonof, P.; Hines, A.H.; Grosholz, E.D. Nonindigenous species as stressors in estuarine and marine communities: Assessing invasion impacts and interactions. Limnol. Oceanogr. 1999, 44, 950–972. [Google Scholar] [CrossRef] [Green Version]
- Osborne, K.L.; Hannigan, R.E.; Poynton, H.C. Differential copper toxicity in invasive and native ascidians of New England provides support for enhanced invader tolerance. Mar. Ecol. Prog. Ser. 2018, 595, 135–147. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentrations | PAHs (μg/L) | Cu (mg/L) |
|---|---|---|
| A | 192 | 0.01 |
| B | 19.2 × 102 | 0.05 |
| C | 19.2 × 103 | 0.1 |
| D | 19.2 × 104 | 0.3 |
| E | 19.2 × 105 | 1 |
| F | - | 5 |
| Scores | Aspect and Morphology |
|---|---|
| 1 | Zooids in healthy state, transparent zooids and ampullae, vessels in normal shape, open siphons, good hemolymph circulation |
| 2 | Zooids in healthy state but slightly pigmented and covered by microalgae, some closed oral siphons, not active filtering |
| 3 | Zooids and ampullae pigmented, some suffering zooids partially detached from the center, thick tunic, but normal vessels and ampullae normally distant from the colony |
| 4 | Zooids, ampullae, and vessels very dark and strict, zooids detached from the center and suffering, small and black, ruined shape, closed oral siphons. Peripheral circulation is limited, heartbeat (bpm) slows, and central circulation keeps the colony alive. |
| 5 | Zooids are dead, no heartbeat, and no circulation. Colony is darkish. |
| Time | Salinity (PSU) | pH | NO2– -N (mg/L) | NH3 -N (mg/L) |
|---|---|---|---|---|
| t0 | 38.0 ± 0.0 | 8.14 ± 0.01 | 0.008 ± 0.002 | 0.06 ± 0.012 |
| t48 | 40.0 ± 0.0 | 8.02 ± 0.06 | 0.004 ± 0.001 | 0.76 ± 0.84 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gregorin, C.; Albarano, L.; Somma, E.; Costantini, M.; Zupo, V. Assessing the Ecotoxicity of Copper and Polycyclic Aromatic Hydrocarbons: Comparison of Effects on Paracentrotus lividus and Botryllus schlosseri, as Alternative Bioassay Methods. Water 2021, 13, 711. https://doi.org/10.3390/w13050711
Gregorin C, Albarano L, Somma E, Costantini M, Zupo V. Assessing the Ecotoxicity of Copper and Polycyclic Aromatic Hydrocarbons: Comparison of Effects on Paracentrotus lividus and Botryllus schlosseri, as Alternative Bioassay Methods. Water. 2021; 13(5):711. https://doi.org/10.3390/w13050711
Chicago/Turabian StyleGregorin, Chiara, Luisa Albarano, Emanuele Somma, Maria Costantini, and Valerio Zupo. 2021. "Assessing the Ecotoxicity of Copper and Polycyclic Aromatic Hydrocarbons: Comparison of Effects on Paracentrotus lividus and Botryllus schlosseri, as Alternative Bioassay Methods" Water 13, no. 5: 711. https://doi.org/10.3390/w13050711
APA StyleGregorin, C., Albarano, L., Somma, E., Costantini, M., & Zupo, V. (2021). Assessing the Ecotoxicity of Copper and Polycyclic Aromatic Hydrocarbons: Comparison of Effects on Paracentrotus lividus and Botryllus schlosseri, as Alternative Bioassay Methods. Water, 13(5), 711. https://doi.org/10.3390/w13050711