
Performance Assessment of Posidonia oceanica (L.) Delile Restoration Experiment on Dead matte Twelve Years after Planting—Structural and Functional Meadow Features


 ,
, 

Abstract
:1. Introduction
- Urban and industrial wastewater treatment and wastewater disposal.
- Legislative instruments, based on EU Directives, aimed at environmental assessment of projects (Environmental Impact Assessment—E.I.A.), plans and programs (Strategic Environmental Assessment–S.E.A).
- Establishment of parks, marine protected areas and reserves along the coastal zone.
2. Materials and Methods
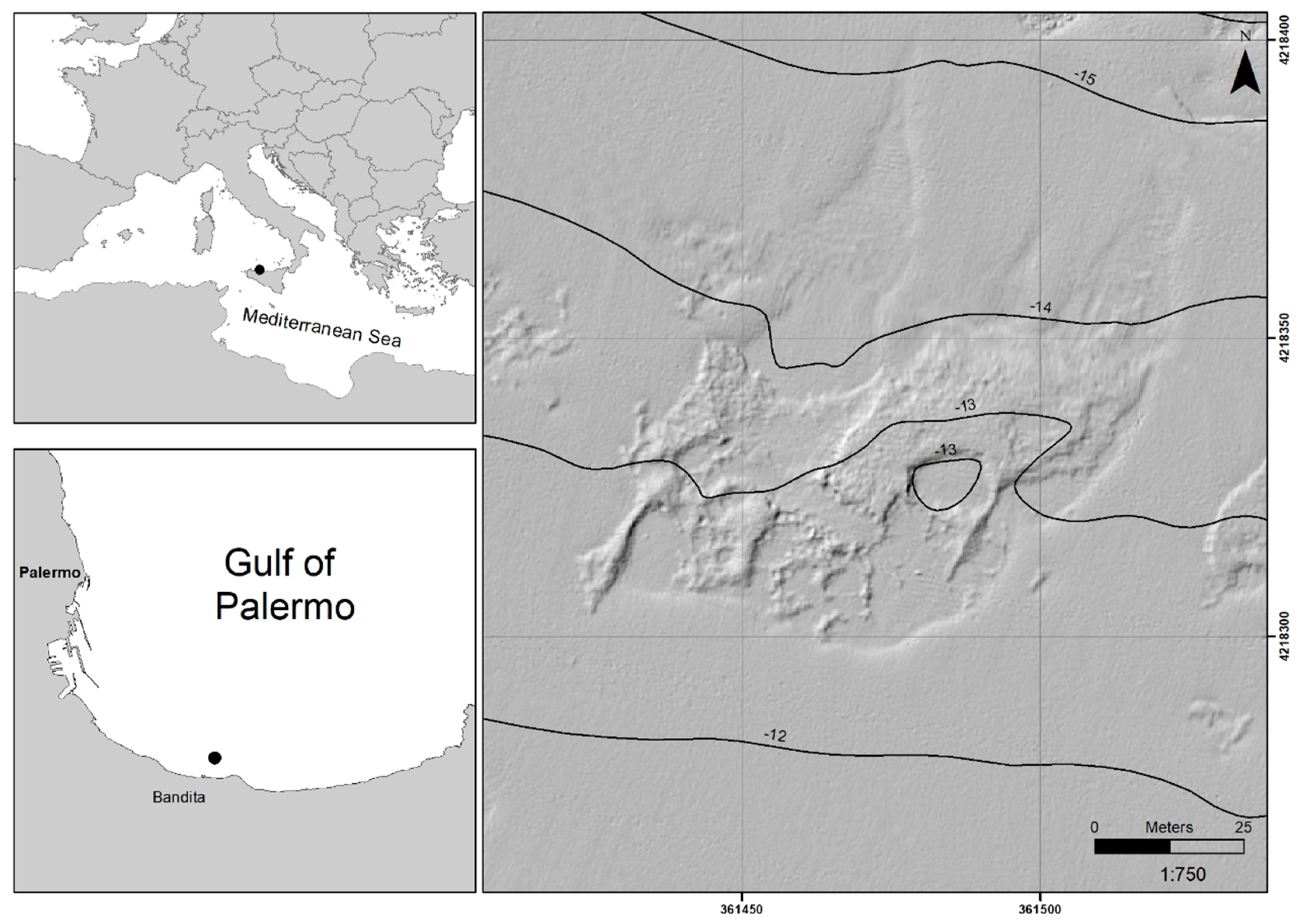
2.1. Study Area
2.2. Data Acquisitions, Field Measurements, Sampling and Laboratory Analysis
2.2.1. Image Acquisitions
2.2.2. Acoustic Survey
2.2.3. Density Assessment and Shoots Sampling
2.3. Data Processing and Statistical Analysis
2.3.1. Image and Acoustic Data
2.3.2. Biometry
3. Results
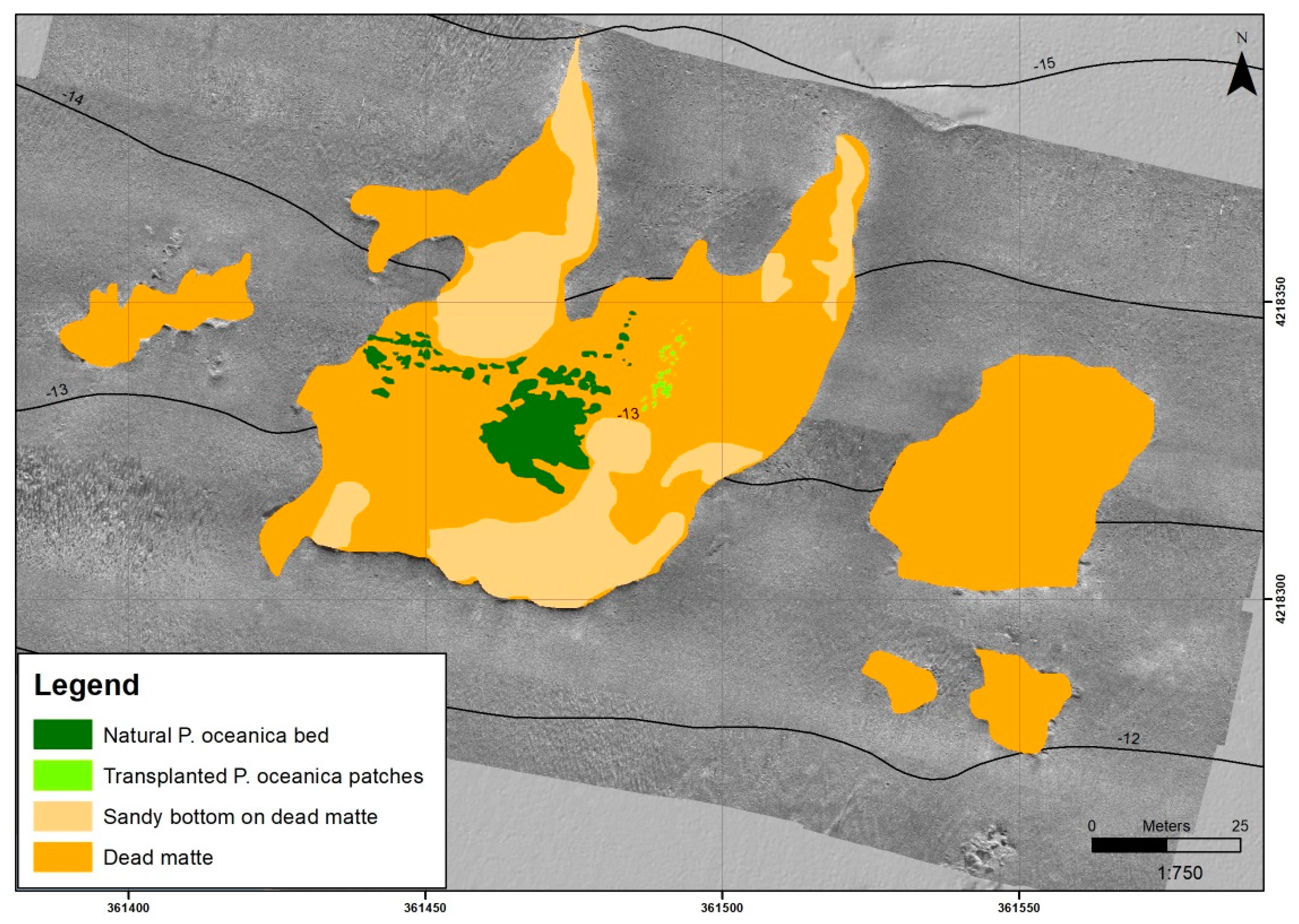
3.1. High-Resolution Sonar Data and Photo Mosaic
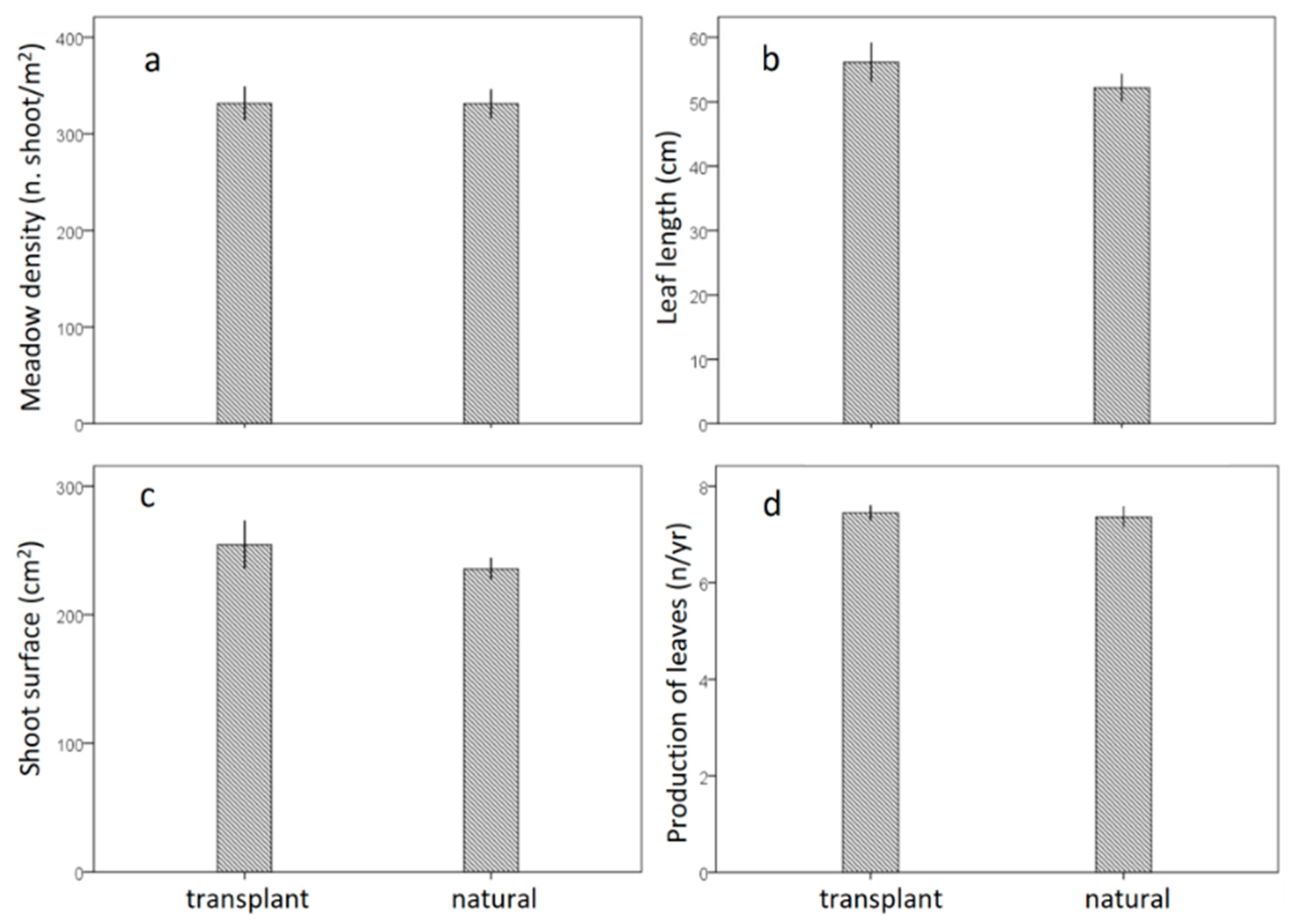
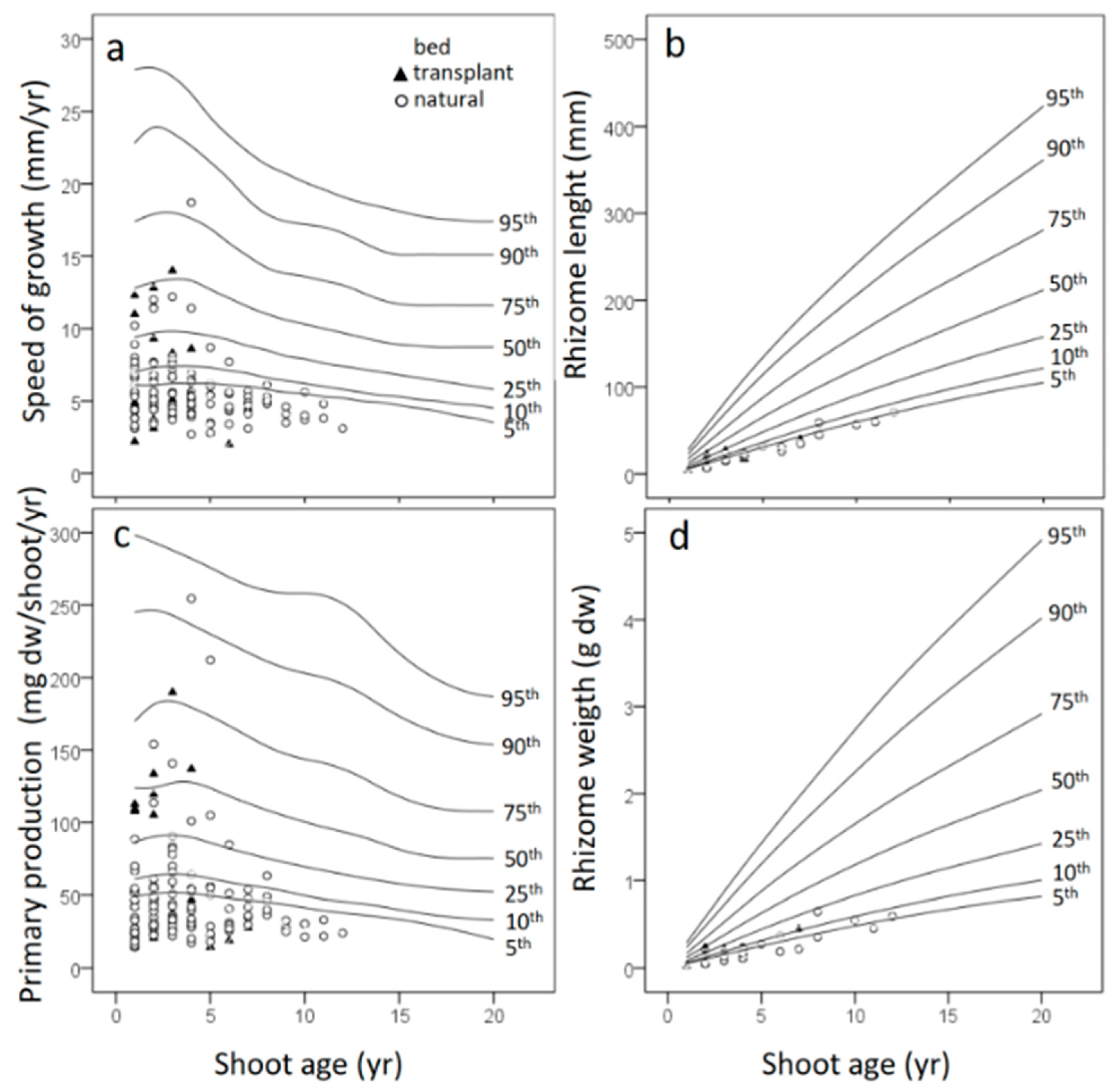
3.2. Leaf Biometry
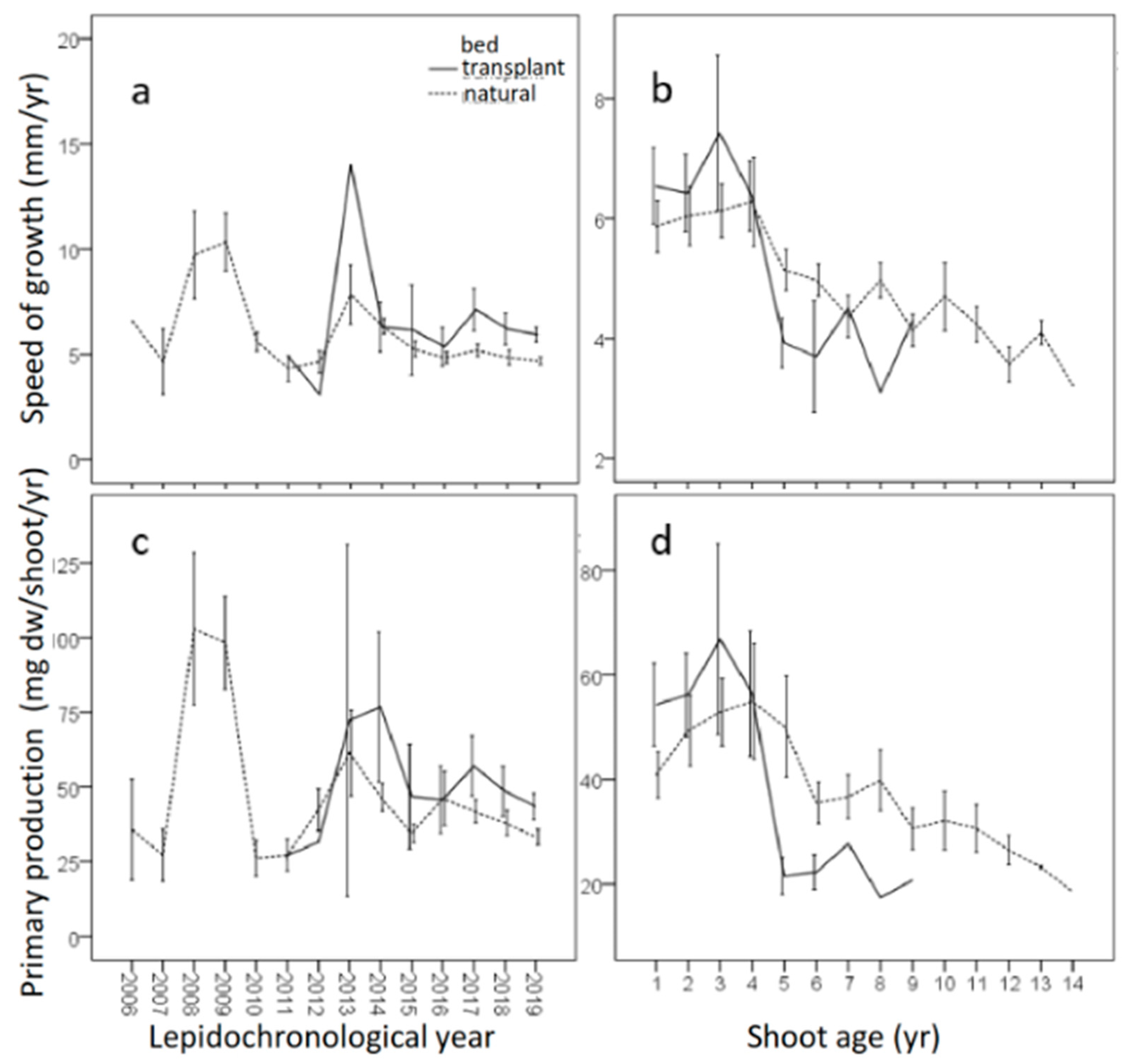
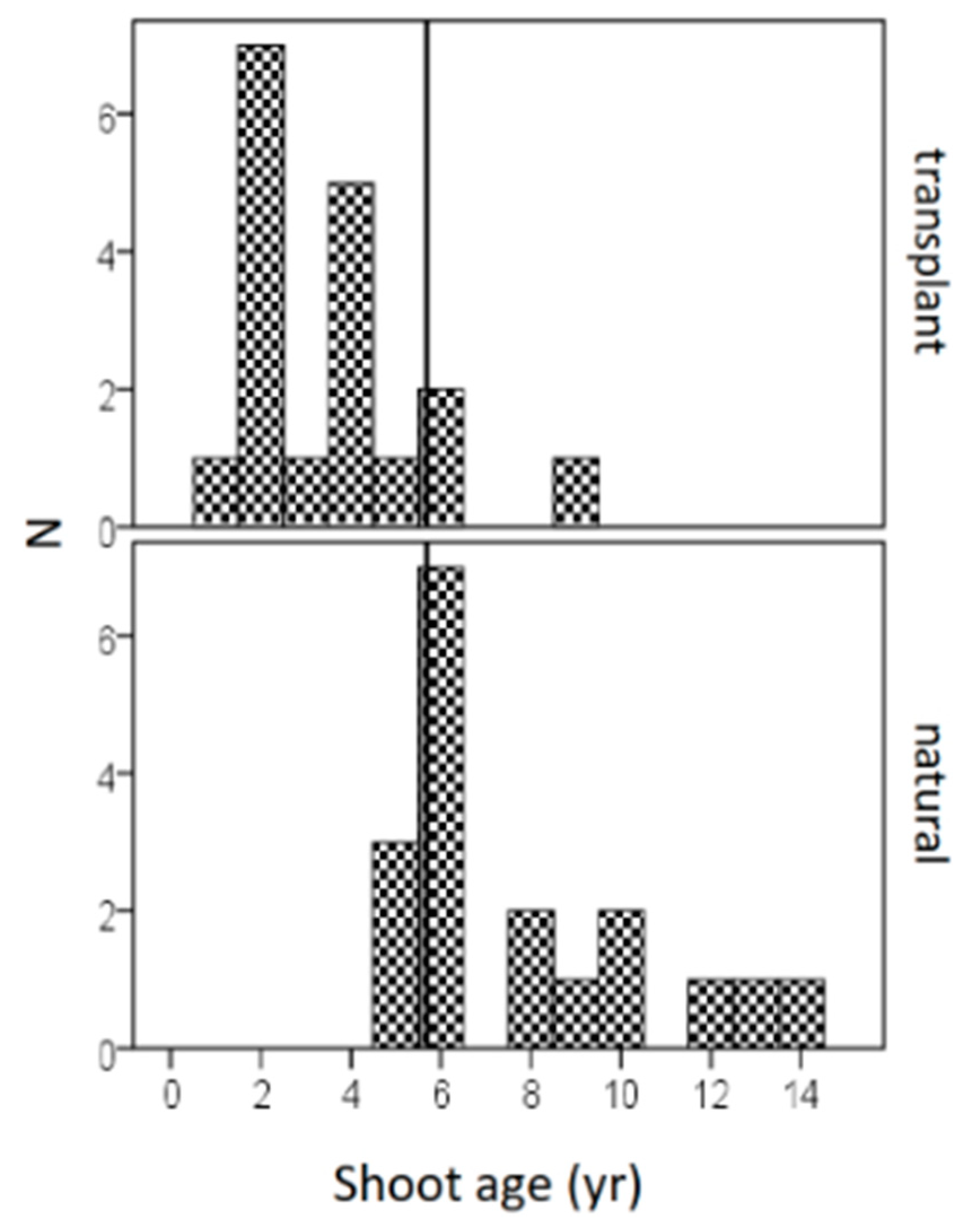
3.3. Dating Measures
3.4. Leaf Primary Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Waltham, N.J.; Elliott, M.; Lee, S.Y.; Lovelock, C.; Duarte, C.M.; Buelow, C.; Simenstad, C.; Nagelkerken, I.; Claassens, L.; Wen, C.K.-C.; et al. UN Decade on Ecosystem Restoration 2021–2030—What Chance for Success in Restoring Coastal Ecosystems? Front. Mar. Sci. 2020, 7. [Google Scholar] [CrossRef] [Green Version]
- Waycott, M.; Duarte, C.M.; Carruthers, T.J.; Orth, R.J.; Dennison, W.C.; Olyarnik, S.; Calladine, A.; Fourqurean, J.W.; Heck, K.L.; Hughes, A.R.; et al. Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sci. USA 2009, 106, 12377–12381. [Google Scholar] [CrossRef] [Green Version]
- Short, F.T.; Polidoro, B.; Livingstone, S.R.; Carpenter, K.E.; Bandeira, S.; Bujang, J.S.; Calumpong, H.P.; Carruthers, T.J.B.; Coles, R.G.; Dennison, W.C.; et al. Extinction risk assessment of the world’s seagrass species. Biol. Conserv. 2011, 144, 1961–1971. [Google Scholar] [CrossRef]
- Kemp, W.M.; Boynton, W.R.; Adolf, J.E.; Boesch, D.F.; Boicourt, W.C.; Brush, G.; Cornwell, J.C.; Fisher, T.R.; Glibert, P.M.; Hagy, J.D.; et al. Eutrophication of Chesapeake Bay: Historical trends and ecological interactions. Mar. Ecol. Prog. Ser. 2005, 303, 1–29. [Google Scholar] [CrossRef]
- González-Correa, J.M.; Bayle, J.T.; Sánchez-Lizaso, J.L.; Valle, C.; Sánchez-Jerez, P.; Ruiz, J.M. Recovery of deep Posidonia oceanica meadows degraded by trawling. J. Exp. Mar. Biol. Ecol. 2005, 320, 65–76. [Google Scholar] [CrossRef]
- Pergent-Martini, C.; Leoni, V.; Pasqualini, V.; Ardizzone, G.D.; Balestri, E.; Bedini, R.; Belluscio, A.; Belsher, T.; Borg, J.; Boudouresque, C.F.; et al. Descriptors of Posidonia oceanica meadows: Use and application. Ecol. Indic. 2005, 5, 213–230. [Google Scholar] [CrossRef]
- Orth, R.J.; Carruthers, T.J.; Dennison, W.C.; Duarte, C.M.; Fourqurean, J.W.; Heck, K.L.; Hughes, A.R.; Kendrick, G.A.; Kenworthy, W.J.; Olyarnik, S. A Global Crisis for Seagrass Ecosystems. Bioscience 2006, 56, 987–996. [Google Scholar] [CrossRef] [Green Version]
- Walker, D.I.; Kendrick, G.A.; McComb, A.J. Decline and Recovery of Seagrass Ecosystems—The Dynamics of Change. In Seagrasses: Biology, Ecology and Conservation; Larkum, A.W.D., Orth, R.J., Duarte, C.M., Eds.; Springer: Berlin, Germany, 2006; pp. 551–565. [Google Scholar]
- Díaz-Almela, E.; Marbà, N.; Álvarez, E.; Santiago, R.; Holmer, M.; Grau, A.; Mirto, S.; Danovaro, R.; Petrou, A.; Argyrou, M. Benthic input rates predict seagrass (Posidonia oceanica) fish farm-induced decline. Mar. Pollut. Bull. 2008, 56, 1332–1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boudouresque, C.F.; Bernard, G.; Pergent, G.; Shili, A.; Verlaque, M. Regression of Mediterranean seagrasses caused by natural processes and anthropogenic disturbances and stress: A critical review. Bot. Mar. 2009, 52, 395–418. [Google Scholar] [CrossRef]
- Pirrotta, M.; Tomasello, A.; Scannavino, A.; Di Maida, G.; Luzzu, F.; Bellissimo, G.; Bellavia, C.; Costantini, C.; Orestano, C.; Sclafani, G. Transplantation assessment of degraded Posidonia oceanica habitats: Site selection and long-term monitoring. Mediterr. Mar. Sci. 2015, 16, 591–604. [Google Scholar] [CrossRef] [Green Version]
- Marbà, N.; Díaz-Almela, E.; Duarte, C.M. Mediterranean seagrass (Posidonia oceanica) loss between 1842 and 2009. Biol. Conserv. 2014, 176, 183–190. [Google Scholar] [CrossRef]
- Telesca, L.; Belluscio, A.; Criscoli, A.; Ardizzone, G.; Apostolaki, E.T.; Fraschetti, S.; Gristina, M.; Knittweis, L.; Martin, C.S.; Pergent, G. Seagrass meadows (Posidonia oceanica) distribution and trajectories of change. Sci. Rep. 2015, 5, srep12505. [Google Scholar] [CrossRef] [Green Version]
- Duarte, C.M.; Middelburg, J.J.; Caraco, N. Major role of marine vegetation on the oceanic carbon cycle. Biogeosciences 2005, 2, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, H.; Björk, M. Seagrass Meadows. In The Management of Natural Coastal Carbon Sinks; Lafoley, D., Grimsditch, G., Eds.; IUCM: Gland, Switzerland, 2009; p. 23. [Google Scholar]
- Marbà, N.; Arias-Ortiz, A.; Masqué, P.; Kendrick, G.A.; Mazarrasa, I.; Bastyan, G.R.; Garcia-Orellana, J.; Duarte, C.M. Impact of seagrass loss and subsequent revegetation on carbon sequestration and stocks. J. Ecol. 2015, 103, 296–302. [Google Scholar] [CrossRef]
- Atwood, T.B.; Connolly, R.M.; Almahasheer, H.; Carnell, P.E.; Duarte, C.M.; Ewers Lewis, C.J.; Irigoien, X.; Kelleway, J.J.; Lavery, P.S.; Macreadie, P.I.; et al. Global patterns in mangrove soil carbon stocks and losses. Nat. Clim. Chang. 2017, 7, 523–528. [Google Scholar] [CrossRef]
- Jordà, G.; Marbà, N.; Duarte, C.M. Mediterranean seagrass vulnerable to regional climate warming. Nat. Clim. Chang. 2012, 2, 821–824. [Google Scholar] [CrossRef] [Green Version]
- González-Correa, J.M.; Sempere, J.T.B.; Sánchez-Jerez, P.; Valle, C. Posidonia oceanica meadows are not declining globally. Analysis of population dynamics in marine protected areas of the Mediterranean Sea. Mar. Ecol. Prog. Ser. 2007, 336, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Calvo, S.; Pirrotta, M.; Tomasello, A. Letter to the editor regarding the article “Taking advantage of seagrass recovery potential to develop novel and effective meadow rehabilitation methods” by Alagna et al., published in Marine Pollution Bulletin, 149: 2019 (110578). Mar. Pollut. Bull. 2020, 158, 111395. [Google Scholar] [CrossRef] [PubMed]
- Costanza, R.; d’Arge, R.; de Groot, R.; Farber, S.; Grasso, M.; Hannon, B.; Limburg, K.; Naeem, S.; O’Neill, R.V.; Paruelo, J.; et al. The value of the world’s ecosystem services and natural capital. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- Duarte, C.; Borum, J.; Short, F.T.; Walker, D.I. Seagrass Ecosystems: Their Global Status and Prospects. In Aquatic Ecosystems: Trends and Global Prospects; Cambridge University Press: Cambridge, UK, 2008; pp. 281–294. [Google Scholar] [CrossRef]
- Fonseca, M.S.; Kenworthy, W.J.; Courtney, F.X.; Hall, M.O. Seagrass Planting in the Southeastern United States: Methods for Accelerating Habitat Development. Restor. Ecol. 1994, 2, 198–212. [Google Scholar] [CrossRef]
- Campbell, M.L. An Empirical Evaluation of Posidonia australis (R. Br.) Hook f. Restoration in Western Australia: Development of a Decision-Based Restoration Framework. Ph.D. Thesis, Murdoch University, Perth, WA, Australia, 2000. [Google Scholar]
- Cunha, A.H.; Marbá, N.N.; Van Katwijk, M.M.; Pickerell, C.; Henriques, M.; Bernard, G.; Ferreira, M.A.; Garcia, S.; Garmendia, J.M.; Manent, P. Changing Paradigms in Seagrass Restoration. Restor. Ecol. 2012, 20, 427–430. [Google Scholar] [CrossRef]
- Calumpong, H.P.; Fonseca, M.S. Seagrass transplantation and other seagrass restoration methods. In Global Seagrass Research Methods; Short, F.T., Coles, R.G., Eds.; Elsevier Science: New York, NY, USA, 2001; pp. 425–442. [Google Scholar]
- Fonseca, M.S.; Kenworthy, W.J.; Thayer, G.W. Guidelines for the Conservation and Restoration of Seagrasses in the United States and Adjacent Waters; Science for Solution, NOAA Coastal Ocean Program, Decision Analysis Series No. 12; U.S. Department of Commerce, National Oceanic and Atmospheric Administration, NOAA Coastal Ocean Office: Silver Spring, MD, USA, 1998; p. 242. [Google Scholar]
- van Katwijk, M.M.; Bos, A.R.; de Jonge, V.N.; Hanssen, L.S.A.M.; Hermus, D.C.R.; de Jong, D.J. Guidelines for seagrass restoration: Importance of habitat selection and donor population, spreading of risks, and ecosystem engineering effects. Mar. Pollut. Bull. 2009, 58, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Paling, E.I.; Van Keulen, M.; Wheeler, K.D.; Phillips, J.; Dyhrberg, R. Influence of Spacing on Mechanically Transplanted Seagrass Survival in a High Wave Energy Regime. Restor. Ecol. 2003, 11, 56–61. [Google Scholar] [CrossRef]
- Bull, J.S.; Reed, D.C.; Holbrook, S.J. An Experimental Evaluation of Different Methods of Restoring Phyllospadix torreyi (Surfgrass). Restor. Ecol. 2004, 12, 70–79. [Google Scholar] [CrossRef]
- Meinesz, A.; Caye, G.; Loquès, F.; Molenaar, H. Polymorphism and Development of Posidonia oceanica Transplanted from Different Parts of the Mediterranean into the National Park of Port-Cros. Bot. Mar. 1993, 36, 209–216. [Google Scholar] [CrossRef]
- Campbell, M.L. Getting the Foundation Right: A Scientifically Based Management Framework to Aid in the Planning and Implementation of Seagrass Transplant Efforts. Bull. Mar. Sci. 2002, 71, 1405–1414. [Google Scholar]
- Short, F.T.; Davis, R.C.; Kopp, B.S.; Short, C.A.; Burdick, D.M. Site-selection model for optimal transplantation of eelgrass Zostera marina in the northeastern US. Mar. Ecol. Prog. Ser. 2002, 227, 253–267. [Google Scholar] [CrossRef]
- Lanuru, M.; Mashoreng, S.; Amri, K. Using site-selection model to identify suitable sites for seagrass transplantation in the west coast of South Sulawesi. J. Phys. Conf. Ser. 2018, 979, 012007. [Google Scholar] [CrossRef]
- Genchi, G.; Di Bernardo, F.; Lugaro, A.; Calvo, S.; Ragonese, S.; Riggio, S. Dystrophic and Eutrophic States of Coastal Sea-Water in Palermo Bay during the Summer Season (August 1981). In VI Journées Etud. Pollutions; CIESM: Cannes, France, 1982; pp. 635–639. [Google Scholar]
- Tomasello, A.; Calvo, S.; Di Maida, G.; Lovison, G.; Pirrotta, M.; Sciandra, M. Shoot age as a confounding factor on detecting the effect of human-induced disturbance on Posidonia oceanica growth performance. J. Exp. Mar. Biol. Ecol. 2007, 343, 166–175. [Google Scholar] [CrossRef]
- Vollenweider, R.A.; Giovanardi, F.; Montanari, G.; Rinaldi, A. Characterisation of the trophic conditions of marine coastal waters, with special reference to the NW Adriatic Sea: Proposal for trophic scale, turbidity and generalised water quality index. Envirometrics 1998, 9, 29–357. [Google Scholar] [CrossRef]
- ARPA Sicilia, Università degli Studi di Palermo. Studi Applicativi Finalizzati All’attivazione del Sistema di Monitoraggio Delle Acque Marino Costiere Della Regione Sicilia; Environmental Protection Agency: Sicilia, Italy, 2006; 227p. [Google Scholar]
- Tomasello, A.; Luzzu, F.; Di Maida, G.; Orestano, C.; Pirrotta, M.; Scannavino, A.; Calvo, S. Detection and Mapping of Posidonia oceanica Dead Matte by High-Resolution Acoustic Imaging. Ital. J. Remote Sens. 2009, 41, 139–146. [Google Scholar] [CrossRef]
- Rende, S.F.; Irving, A.D.; Bacci, T.; Parlagreco, L.; Bruno, F.; De Filippo, F.; Montefalcone, M.; Penna, M.; Trabucco, B.; Di Mento, R.; et al. Advances in micro-cartography: A two-dimensional photo mosaicing technique for seagrass monitoring. Estuar. Coast. Shelf Sci. 2015, 167, 475–486. [Google Scholar] [CrossRef]
- Rende, S.F.; Bosman, A.; Di Mento, R.; Bruno, F.; Lagudi, A.; Irving, A.D.; Dattola, L.; Di Giambattista, L.; Lanera, P.; Proietti, R.; et al. Ultra-High-Resolution Mapping of Posidonia oceanica (L.) Delile Meadows through Acoustic, Optical Data and Object-based Image Classification. J. Mar. Sci. Eng. 2020, 8, 647. [Google Scholar] [CrossRef]
- Giraud, G. Contribution à la Description et à la Phénologie Quantitative des Herbiers à Posidonia oceanica (L.) Delile. Ph.D. Thesis, Université Aix–Marseille II, Marseille, France, 1977. [Google Scholar]
- Giraud, G. Sur Une Méthode de Mesure et de Comptage Des Structures Foliaires de Posidonia oceanica (Linnaeus) Delile. Bull. Mus. Hist. Nat. Marseille 1979, 39, 33–39. [Google Scholar]
- Pergent, G. Lepidochronological analysis of the seagrass Posidonia oceanica (L.) Delile: A standardized approach. Aquat. Bot. 1990, 37, 39–54. [Google Scholar] [CrossRef]
- Pergent, G.; Pergent-Martini, C. Some Applications of Lepidochronological Analysis in the Seagrass Posidonia oceanica. Bot. Mar. 1990, 33, 299–310. [Google Scholar] [CrossRef]
- Tomasello, A.; Sciandra, M.; Muggeo, V.M.R.; Pirrotta, M.; Di Maida, G.; Calvo, S. Reference growth charts for Posidonia oceanica seagrass: An effective tool for assessing growth performance by age and depth. Ecol. Indic. 2016, 69, 50–58. [Google Scholar] [CrossRef]
- Pergent, G.; Boudouresque, C.-F.; Crouzet, A.; Meinesz, A. Cyclic Changes along Posidonia oceanica Rhizomes (Lepidochronology): Present State and Perspectives. Mar. Ecol. 1989, 10, 221–230. [Google Scholar] [CrossRef]
- Beaudoin, J.; Hughes Clarke, J.E.; Van den Ameele, E.J.; Gardner, J.V. Geometric and Radiometric Correction of Multibeam Backscatter Derived from Reson 8101 Systems. 2002. Available online: https://scholars.unh.edu/ccom/242 (accessed on 15 December 2020).
- Underwood, A.J. Experiments in Ecology; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Calvo, S.; Lovison, G.; Pirrotta, M.; Di Maida, G.; Tomasello, A.; Sciandra, M. Modelling the relationship between sexual reproduction and rhizome growth in Posidonia oceanica (L.) Delile. Mar. Ecol. 2006, 27, 361–371. [Google Scholar] [CrossRef]
- Vizzini, S.; Tomasello, A.; Di Maida, G.; Pirrotta, M.; Mazzola, A.; Calvo, S. Effect of explosive shallow hydrothermal vents on δ13C and growth performance in the seagrass Posidonia oceanica. J. Ecol. 2010, 98, 1284–1291. [Google Scholar] [CrossRef]
- Lovison, G.; Sciandra, M.; Tomasello, A.; Calvo, S. Modeling Posidonia oceanica growth data: From linear to generalized linear mixed models. Environmetrics 2011, 22, 370–382. [Google Scholar] [CrossRef]
- Di Maida, G.; Tomasello, A.; Sciandra, M.; Pirrotta, M.; Milazzo, M.; Calvo, S. Effect of different substrata on rhizome growth, leaf biometry and shoot density of Posidonia oceanica. Mar. Environ. Res. 2013, 87–88, 96–102. [Google Scholar] [CrossRef]
- Pergent, G.; Pergent-Martini, C. Leaf renewal cycle and primary production of Posidonia oceanica in the bay of Lacco Ameno (Ischia, Italy) using lepidochronological analysis. Aquat. Bot. 1991, 42, 49–66. [Google Scholar] [CrossRef]
- Marbá, N.; Duarte, C.M.; Cebrián, J.; Gallegos, M.E.; Olesen, B.; Sand-Jensen, K. Growth and Population Dynamics of Posidonia oceanica on the Spanish Mediterranean Coast: Elucidating Seagrass Decline. Mar. Ecol. Prog. Ser. 1996, 137, 203–213. [Google Scholar] [CrossRef]
- Duarte, C.M.; Uri, J.S.; Agawin, N.S.R.; Fortes, M.D.; Vermaat, J.E.; Marba, N. Flowering Frequency of Philippine Seagrasses. Bot. Mar. 1997, 40, 497–500. [Google Scholar] [CrossRef]
- La Loggia, G.; Calvo, S.; Ciraolo, G.; Mazzola, A.; Pirrotta, M.; Sara, G.; Tomasello, A.; Vizzini, S. Influence of Hydrodynamic Conditions on the Production and Fate of Posidonia oceanica in a Semi-Enclosed Shallow Basin (Stagnone Di Marsala, Western Sicily). Chem. Ecol. 2004, 20, 183–201. [Google Scholar] [CrossRef]
- Marbà, N.; Hemminga, M.A.; Mateo, M.A.; Duarte, C.M.; Mass, Y.E.; Terrados, J.; Gacia, E. Carbon and Nitrogen Translocation between Seagrass Ramets. Mar. Ecol. Prog. Ser. 2002, 226, 287–300. [Google Scholar] [CrossRef] [Green Version]
- Ralph, P.J.; Durako, M.J.; Enríquez, S.; Collier, C.J.; Doblin, M.A. Impact of light limitation on seagrasses. J. Exp. Mar. Biol. Ecol. 2007, 350, 176–193. [Google Scholar] [CrossRef]
- Buia, M.C.; Flagella, S.; Guala, I.; Gravina, T.; Flagella, M.M.; Cigliano, M. Interannual and Decadal Variability in Posidonia oceanica. In Proceedings of the 7th International Conference on the Mediterranean Coastl Environment MEDCOAST, Kusadasi, Turkey, 25–29 October 2005; Volume 5, pp. 407–418. [Google Scholar]
- Duarte, C.M. Allometric scaling of seagrass form and productivity. Mar. Ecol. Prog. Ser. Oldendorf 1991, 77, 289–300. [Google Scholar] [CrossRef]
- Duarte, C.M.; Fourqurean, J.W.; Krause-Jensen, D.; Olesen, B. Dynamics of Seagrass Stability and Change. In Seagrasses: Biology, Ecology and Conservation; Larkum, A.W.D., Orth, R.J., Duarte, C.M., Eds.; Springer: Berlin, Germany, 2006; pp. 271–294. [Google Scholar]
- Pergent, G.; Pergent-Martini, C.; Boudouresque, C.-F. Utilisation de l’herbier à Posidonia oceanica Comme Indicateur Biologique de la Qualité du Milieu Littoral en Méditerranée: État des Connaissances. Mésogée (Marseille) 1995, 54, 3–27. [Google Scholar]
- Thom, R.M.; Diefenderfer, H.L.; Vavrinec, J.; Borde, A.B. Restoring Resiliency: Case Studies from Pacific Northwest Estuarine Eelgrass (Zostera marina L.) Ecosystems. Estuaries Coasts 2012, 35, 78–91. [Google Scholar] [CrossRef]
- Meinesz, A.; Lefevre, J.R. Régénération d’un Herbier de Posidonia oceanica Quarante Années Après sa Destruction par une Bombe Dans la Rade de Villefranche (Alpes-Maritimes, France). In International Workshop on Posidonia oceanica Beds; GIS Posidonie Marseille: Marseille, France, 1984; Volume 2, pp. 39–44. [Google Scholar]
- Caye, G. Sur la Morphogénèse et le Cycle Végétatif de Posidonia oceanica (L.) Delile. Ph.D. Thesis, Universitè d’Aix-Marseille II, Marseille, France, 1980; pp. 1–121. [Google Scholar]
- de los Santos, C.B.; Krause-Jensen, D.; Alcoverro, T.; Marbà, N.; Duarte, C.M.; Van Katwijk, M.M.; Pérez, M.; Romero, J.; Sánchez-Lizaso, J.L.; Roca, G.; et al. Recent trend reversal for declining European seagrass meadows. Nat. Commun. 2019, 10, 3356. [Google Scholar] [CrossRef] [Green Version]
- Boudouresque, C.F. Posidonia oceanica Meadows. In Protection and Conservation of Posidonia Oceanica Meadows; Boudouresque, C.F., Bernard, G., Bonhomme, P., Charbonnel, E., Diviacco, G., Meinesz, A., Pergent, G., Pergent-Martini, C., Ruitton, S., Tunesi, L., Eds.; RAMOGE and RAC/SPA Publisher: Tunis, Tunisia, 2012; pp. 10–24. [Google Scholar]
- Moliner, R.; Picard, J. Recherches sur les Herbiers des Phenerogames Marines du Littoral Mediterraneen Francaise. Ann. Inst. Oceanogr. 1952, 27, 157–235. [Google Scholar]
- Boudouresque, C.F.; Jeudy De Grissac, A.; Meinesz, A. Chronologie de l’édification d’une Colline de Posidonies. Trav. Sci. Parc Nat. Rég. Rés. Nat. Corse 1986, 2, 3–12. [Google Scholar]
- Boudouresque, C.F.; Lefevre, J.R.; Meisenz, A. Cartographie du Carré Permanent de la Marina d’Elbu. Trav. Sci. Parc. Nat. Reg. Res. Nat. Corse 1986, 2, 24–35. [Google Scholar]
- Meinesz, A.; Boudouresque, C.; Lefevre, J. A Map of the Posidonia oceanica Beds of Marina ďElbu (Corsica, Mediterranean). Mar. Ecol. 1988, 9, 243–252. [Google Scholar] [CrossRef]
- Gobert, S.; Lepoint, G.; Pelaprat, C.; Remy, F.; Lejeune, P.; Richir, J.; Abadie, A. Temporal evolution of sand corridors in a Posidonia oceanica seascape: A 15-years study. Mediterr. Mar. Sci. 2016, 17, 777. [Google Scholar] [CrossRef] [Green Version]
- Tunesi, L.; Boudouresque, C.F. The Causes of Posidonia oceanica Meadows Regression. In Protection and Conservation of Posidonia oceanica Meadows; Boudouresque, C.F., Bernard, G., Bonhomme, P., Charbonnel, E., Diviacco, G., Meinesz, A., Pergent, G., Pergent-Martini, C., Ruitton, S., Tunesi, L., Eds.; RAMOGE and RAC/SPA Publisher: Tunis, Tunisia, 2012; pp. 32–47. [Google Scholar]
- Abadie, A.; Richir, J.; Lejeune, P.; LeDuc, M.; Gobert, S. Structural Changes of Seagrass Seascapes Driven by Natural and Anthropogenic Factors: A Multidisciplinary Approach. Front. Ecol. Evol. 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- Mateo, M.A.; Cebrìan, J.; Dunton, K.; Mutchler, T. Carbon Flux in Seagrass Ecosystem. In Seagrasses: Biology, Ecology and Conservation; Larkum, A.W.D., Orth, R.J., Duarte, C.M., Eds.; Springer: Berlin, Germany, 2006; pp. 159–192. [Google Scholar]
- Pergent, G.; Bazairi, H.; Bianchi, C.N.; Boudouresque, C.F.; Buia, M.C.; Calvo, S.; Clabaut, P.; Harmelin-Vivien, M.; Mateo, M.A.; Montefalcone, M. Climate change and Mediterranean seagrass meadows: A synopsis for environmental managers. Mediterr. Mar. Sci. 2014, 15, 462. [Google Scholar] [CrossRef] [Green Version]
- United Nations Framework Convention on Climate Change (UNFCCC). Report of the Conference of the Parties on Its Twenty-First Session, Held in Paris from 30 November to 13 December 2015; UNFCCC: Bonn, Germany, 2015; p. 42. [Google Scholar]
- Piano, R. Il Rammendo Delle Periferie. “Il Sole 24 ORE”. Available online: http://www.ilsole24ore.com/art/cultura/2014-06-18/il-rammendo-periferie-094517.shtml?uuid=ABBYPHSB (accessed on 26 January 2014).
- Piano, R. Building Workshop. Ricuciture Urbane e Periferie; Solferino: Milan, Italy, 2019; p. 143, EAN: 9788828202004. [Google Scholar]
- Balestri, E.; Vallerini, F.; Lardicci, C. Storm-generated fragments of the seagrass Posidonia oceanica from beach wrack—A potential source of transplants for restoration. Biol. Conserv. 2011, 144, 1644–1654. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Bed | t | p | |
|---|---|---|---|---|
| Transplant | Natural | |||
| Density (n shoot/m2) | 331.6 (17.7) | 331.2 (14.9) | 0.2 | ns |
| Leaf length (cm) | 56.1 (3.1) | 52.2 (2.2) | 0.1 | ns |
| Shoot surface (cm2) | 254.4 (18.7) | 235.6 (8.7) | 0.9 | ns |
| Shoot age (year) | 3.6 (0.5) | 7.8 (0.7) | 5.1 | *** |
| Production of leaves (n/shoot/year) | 7.4 (0.2) | 7.3 (0.2) | 0.3 | ns |
| Rhizome primary production (mg dw/shoot/year) | 58.7 (8.5) | 47.5 (3.2) | - | - |
| Speed of growth (mm/shoot/year) | 6.4 (0.6) | 5.7 (0.2) | - | - |
| Variables | Bed | |
|---|---|---|
| Transplant | Natural | |
| Leaf density (mg dw/cm) | 4.5 (0.22) | 4.2 (0.12) |
| Production of leaves (n/shoot/year) | 7.4 | 7.3 |
| Past sheet length (cm) | 3.43 (0.9) | 3.82 (0.8) |
| Theoretical past oldest leaf length (cm) | 39.3 | 48.1 |
| Leaf primary production (gr dw/shoot/year) | 1.3 | 1.5 |
| Rhizome primary production (g dw/shoot/year) | 0.06 | 0.05 |
| Total primary production (g dw/shoot/year) | 1.4 | 1.5 |
| Meadow density (n shoots/m2) | 331.6 | 331.2 |
| Covering (m2) | 24.0 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calvo, S.; Calvo, R.; Luzzu, F.; Raimondi, V.; Assenzo, M.; Cassetti, F.P.; Tomasello, A. Performance Assessment of Posidonia oceanica (L.) Delile Restoration Experiment on Dead matte Twelve Years after Planting—Structural and Functional Meadow Features. Water 2021, 13, 724. https://doi.org/10.3390/w13050724
Calvo S, Calvo R, Luzzu F, Raimondi V, Assenzo M, Cassetti FP, Tomasello A. Performance Assessment of Posidonia oceanica (L.) Delile Restoration Experiment on Dead matte Twelve Years after Planting—Structural and Functional Meadow Features. Water. 2021; 13(5):724. https://doi.org/10.3390/w13050724
Chicago/Turabian StyleCalvo, Sebastiano, Roberta Calvo, Filippo Luzzu, Vincenzo Raimondi, Mauro Assenzo, Federica Paola Cassetti, and Agostino Tomasello. 2021. "Performance Assessment of Posidonia oceanica (L.) Delile Restoration Experiment on Dead matte Twelve Years after Planting—Structural and Functional Meadow Features" Water 13, no. 5: 724. https://doi.org/10.3390/w13050724
APA StyleCalvo, S., Calvo, R., Luzzu, F., Raimondi, V., Assenzo, M., Cassetti, F. P., & Tomasello, A. (2021). Performance Assessment of Posidonia oceanica (L.) Delile Restoration Experiment on Dead matte Twelve Years after Planting—Structural and Functional Meadow Features. Water, 13(5), 724. https://doi.org/10.3390/w13050724