
Invertebrate Responses to Restoration across Benthic and Hyporheic Stream Compartments



Abstract
:
1. Introduction
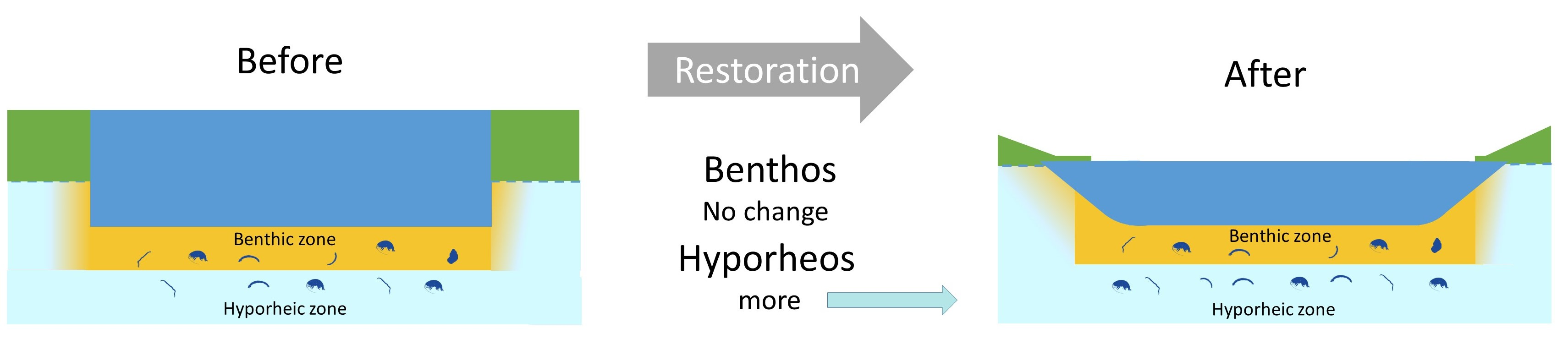
- The hyporheos in the impact reach will differ more strongly in diversity and composition pre- and post-restoration than the control reach, since we expect weir notching will result in an increase in exchange processes between the surface stream and hyporheic zone.
- The responses of the hyporheos to the restoration will differ from the benthos because they are discrete communities [27] driven by differing environmental variables.
2. Methods
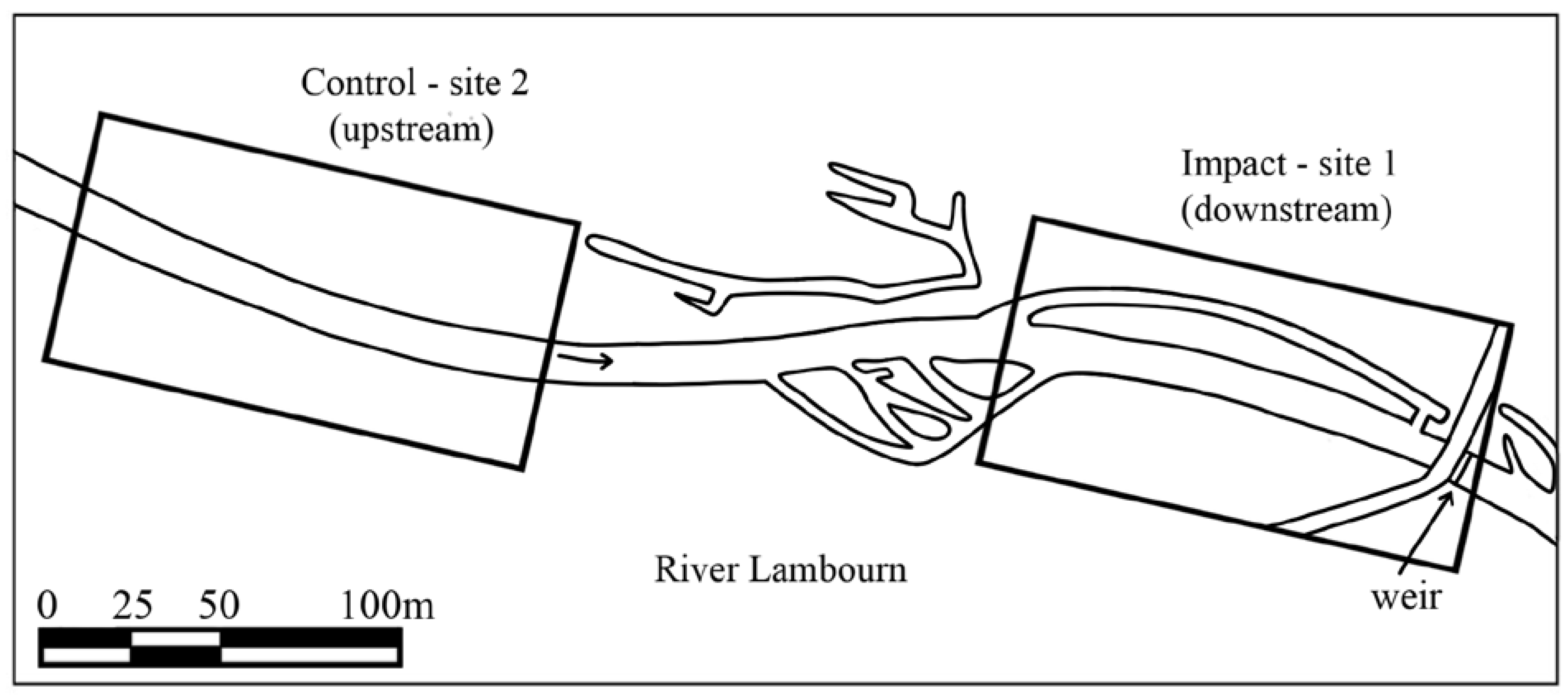
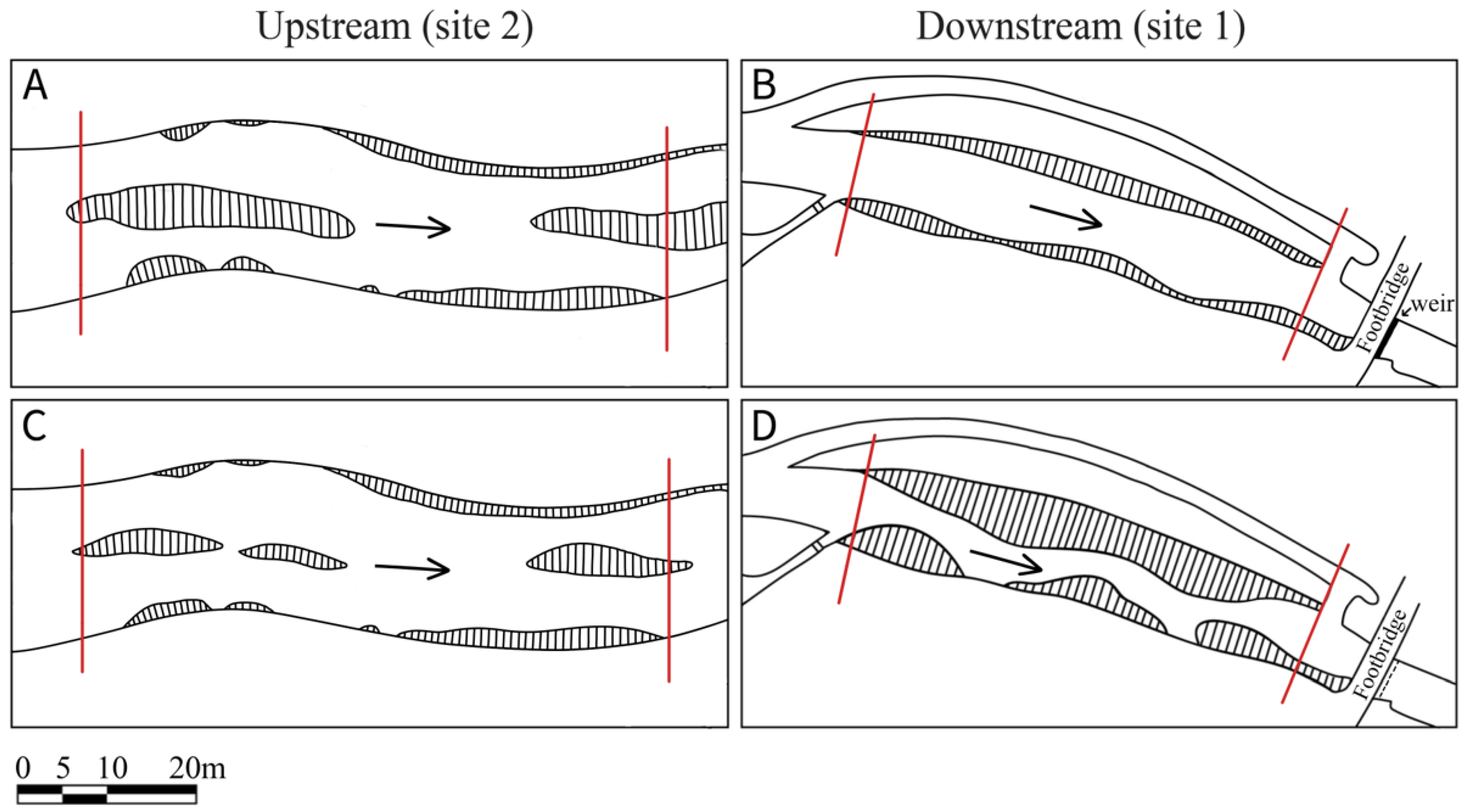
2.1. Restoration Scheme
2.2. Sampling
2.3. Analysis
3. Results
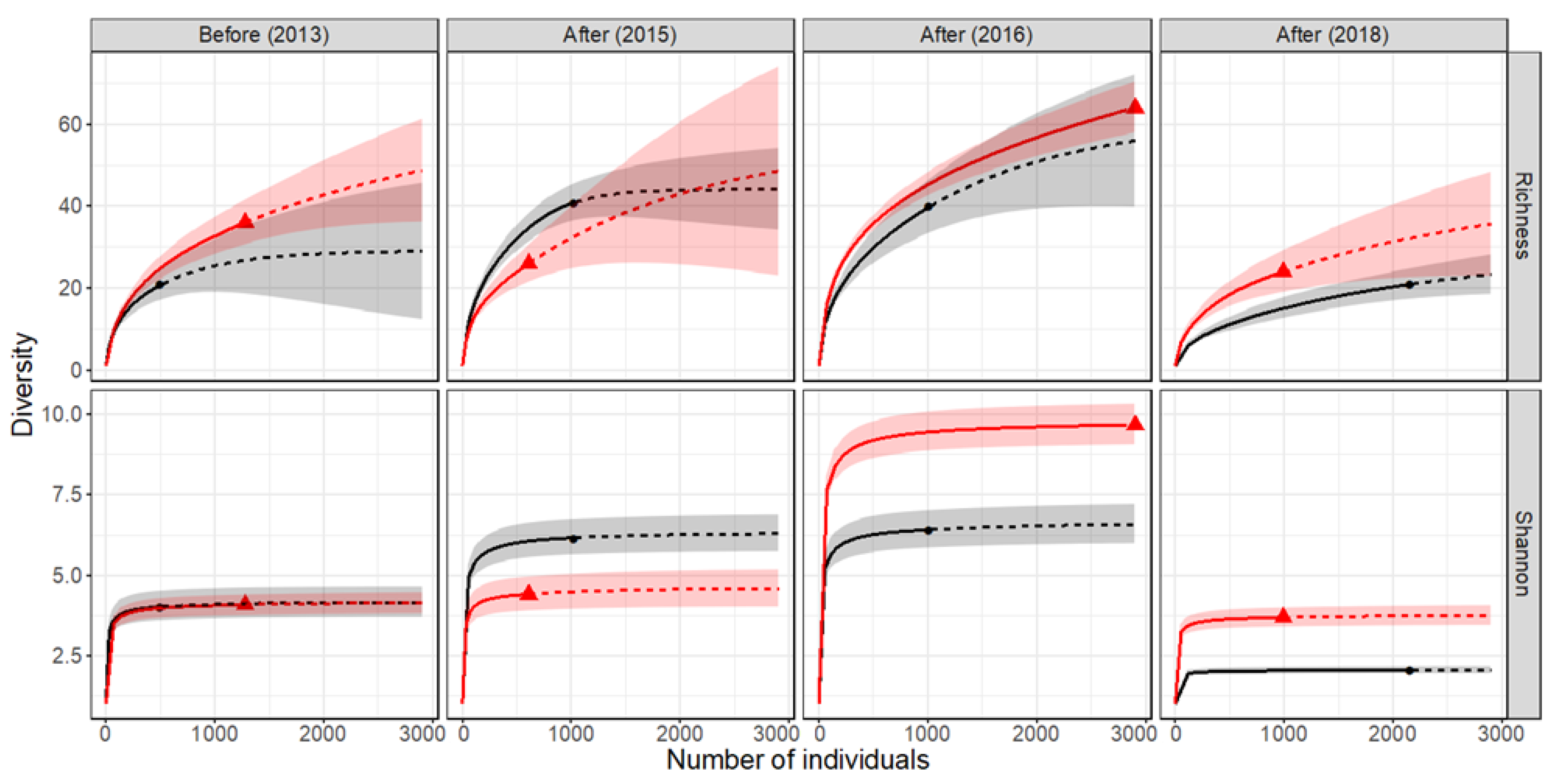
3.1. Hyporheos
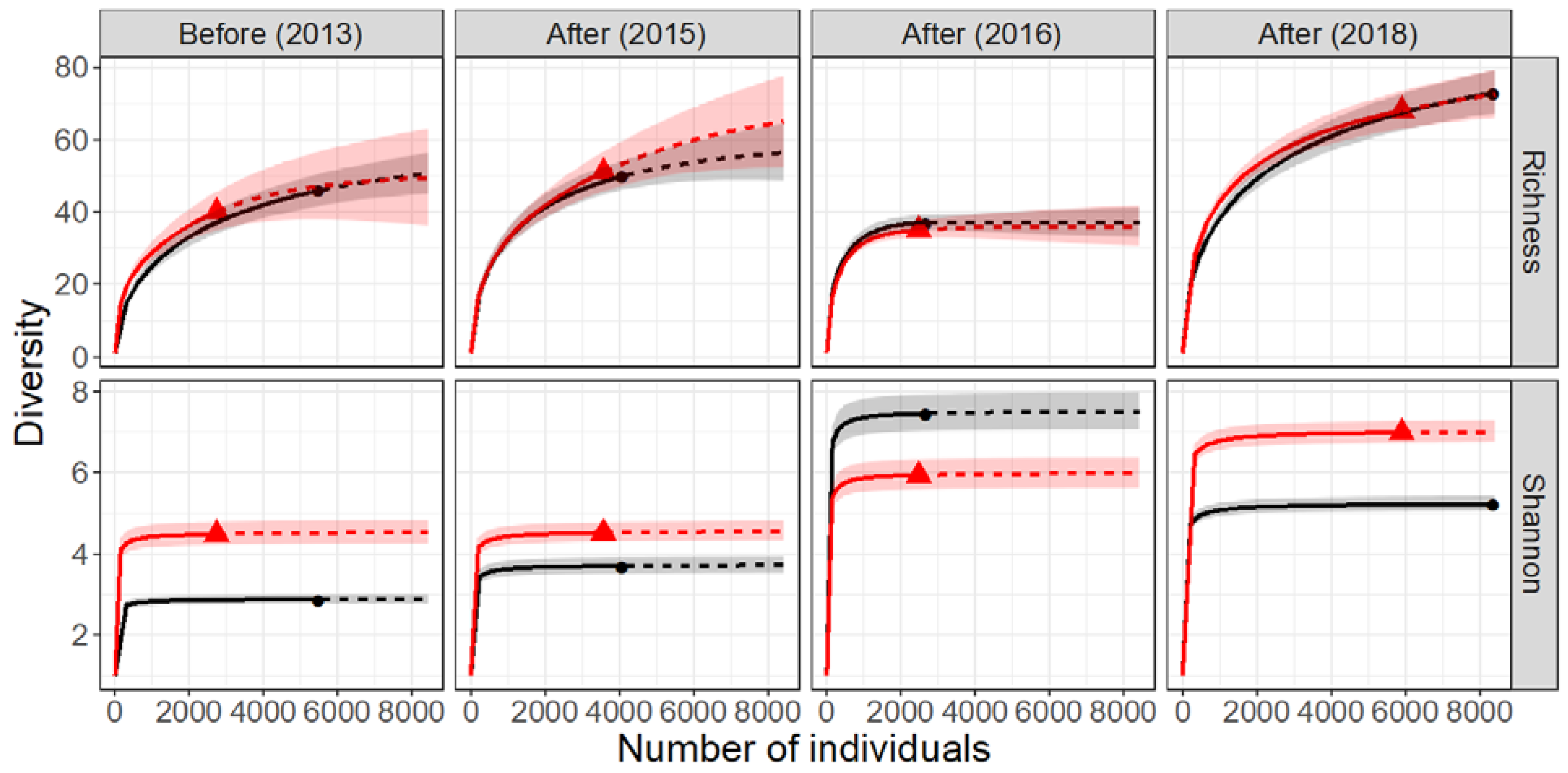
3.2. Benthos
3.3. Coupling of Hyporheic and Benthic Assemblages
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wohl, E.; Lane, S.N.; Wilcox, A.C. The science and practice of river restoration. Water Resour. Res. 2015, 51, 5974–5997. [Google Scholar] [CrossRef] [Green Version]
- Palmer, M.A.; Ambrose, R.F.; Poff, N.L. Ecological theory and community restoration ecology. Restor. Ecol. 1997, 5, 291–300. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, A.W.; Haase, P.; Januschke, K.; Sundermann, A.; Hering, D. Revisiting restored river reaches—Assessing change of aquatic and riparian communities after five years. Sci. Total Environ. 2018, 613–614, 1185–1195. [Google Scholar] [CrossRef] [PubMed]
- Beechie, T.J.; Sear, D.A.; Olden, J.D.; Pess, G.R.; Buffington, J.M.; Moir, H.; Roni, P.; Pollock, M.M. Process-based principles for restoring river ecosystems. Bioscience 2010, 60, 209–222. [Google Scholar] [CrossRef] [Green Version]
- Palmer, M.A.; Hondula, K.L.; Koch, B.J. Ecological Restoration of Streams and Rivers: Shifting Strategies and Shifting Goals. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 247–269. [Google Scholar] [CrossRef] [Green Version]
- Boulton, A.J.; Datry, T.; Kasahara, T.; Mutz, M.; Stanford, J.A. Ecology and management of the hyporheic zone: Stream–groundwater interactions of running waters and their floodplains. J. N. Am. Benthol. Soc. 2010, 29, 26–40. [Google Scholar] [CrossRef] [Green Version]
- Stegen, J.C.; Fredrickson, J.K.; Wilkins, M.J.; Konopka, A.E.; Nelson, W.C.; Arntzen, E.V.; Chrisler, W.B.; Chu, R.K.; Danczak, R.E.; Fansler, S.J.; et al. Groundwater–surface water mixing shifts ecological assembly processes and stimulates organic carbon turnover. Nat. Commun. 2016, 7, 11237. [Google Scholar] [CrossRef] [Green Version]
- Merill, L.; Tonjes, D.J. A review of the hyporheic zone stream restoration, and means to enhance denitrification. Crit. Rev. Environ. Sci. Technol. 2014, 44, 2337–2379. [Google Scholar] [CrossRef]
- Lewandowski, J.; Putschew, A.; Schwesig, D.; Neumann, C.; Radke, M. Fate of organic micropollutants in the hyporheic zone of a eutrophic lowland stream: Results of a preliminary field study. Sci. Total Environ. 2011, 409, 1824–1835. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, C.; Nelson, W.C.; Shi, L.; Xu, F.; Liu, Y.; Yan, A.; Zhong, L.; Thompson, C.; Fredrikson, J.K.; et al. Effect of water chemistry and hydrodynamics on nitrogen transformation activity and microbial community functional potential inhyporheic zone sediment columns. Environ. Sci. Technol. 2017, 51, 4877–4886. [Google Scholar] [CrossRef]
- Robertson, A.L.; Wood, P.J. Ecology of the hyporheic zone: Origins, current knowledge and future directions. Fundam. Appl. Limnol. 2010, 176, 279–289. [Google Scholar] [CrossRef]
- Brunke, M.; Gonser, T. The ecological significance of exchange processes between rivers and groundwater. Freshw. Biol. 1997, 37, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Rutherford, J.E.; Hynes, H.B.N. Dissolved organic carbon in streams and groundwater. Hydrobiologia 1987, 154, 33–48. [Google Scholar] [CrossRef]
- Boano, F.; Harvey, J.W.; Marion, A.; Packman, A.I.; Revelli, R.; Ridolfi, L.; Wörman, A. Hyporheic flow and transport processes: Mechanisms, models, and biogeochemical implications. Rev. Geophys. 2014, 52, 603–679. [Google Scholar] [CrossRef]
- Hancock, P. The Effects of River Stage Fluctuations on the Hyporheic and Parafluvial Ecology of the Hunter River, New South Wales. Ph.D. Thesis, University of New England, Armidale, NSW, Australia, 2004. [Google Scholar]
- Arthington, A.H.; Pusey, B.J. Flow restoration and protection in Australian rivers. River Res. Appl. 2003, 19, 377–395. [Google Scholar] [CrossRef]
- Buss, S.; Cai, Z.; Cardenas, B.; Fleckenstein, J.; Hannah, D.; Heppell, K.; Hulme, P.; Ibrahim, T.; Kaeser, D.; Krause, S.; et al. The Hyporheic Handbook: A Handbook on the Groundwater-Surface-Water Interface and Ecology of the Hyporheic Zone for Environmental Managers; 2009 Environment Agency Science Report SC0 50070; Environment Agency: Bristol, UK, 2009.
- Palmer, M.A.; Bely, A.E.; Berg, K.E. Response of invertebrates to lotic disturbance: A test of the hyporheic refuge hypothesis. Oecologia 1992, 89, 182–194. [Google Scholar] [CrossRef]
- Lafont, M.; Vivier, A.; Nogueira, S.; Namour, P.; Breil, P. Surface and hyporheic oligochaete assemblages in a French suburban stream. Hydrobiologia 2006, 564, 183–193. [Google Scholar] [CrossRef] [Green Version]
- James, A.B.W.; Dewson, Z.S.; Death, R.G. Do stream macroinvertebrates use instream refugia in response to severe short-term flow reduction in New Zealand streams? Freshwat. Biol. 2008, 53, 1316–1334. [Google Scholar] [CrossRef]
- Descloux, S.; Datry, T.; Marmonier, P. Benthic and hyporheic invertebrate assemblages along a gradient of increasing streambed colmation by fine sediment. Aquat. Sci. 2013, 75, 493–507. [Google Scholar] [CrossRef]
- Descloux, S.; Datry, T.; Usseglio-Polatera, P. Trait-based structure of invertebrates along a gradient of sediment colmation: Benthos versus hyporheos responses. Sci. Total Environ. 2014, 466, 265–276. [Google Scholar] [CrossRef]
- Stubbington, R.; Boulton, A.J.; Little, S.; Wood, P.J. Changes in invertebrate assemblage composition in benthic and hyporheic zones during a severe supraseasonal drought. Freshwat. Sci. 2015, 34, 344–354. [Google Scholar] [CrossRef] [Green Version]
- Magliozzi, C.; Usseglio-Polatera, P.; Meyer, A.; Grabowski, R.C. Functional traits of hyporheic and benthic invertebrates reveal importance of wood-driven geomorphological processes in rivers. Funct. Ecol. 2019, 33, 1758–1770. [Google Scholar] [CrossRef]
- Peralta-Maraver, I.; Galloway, J.; Posselt, M.; Arnon, S.; Reiss, J.; Lewandowski, J.; Robertson, A. Environmental filtering and community delineation in the streambed ecotone. Sci. Rep. 2019, 8, 15871. [Google Scholar] [CrossRef]
- Negishi, J.N.; Terui, A.; Nessa, B.; Miura, K.; Oiso, T.; Sumitomo, K.; Kyuka, T.; Yonemoto, M.; Nakamura, F. High resilience of aquatic community to a 100-year flood in a gravel-bed river. Landsc. Ecol. Eng. 2019, 15, 143–154. [Google Scholar] [CrossRef]
- Peralta-Maraver, I.; Reiss, J.; Robertson, A.L. Interplay of hydrology, community ecology and pollution attenuation in the hyporheic zone. Sci. Total Environ. 2018, 610–611, 267–275. [Google Scholar] [CrossRef] [Green Version]
- Bou, C.; Rouch, R. Un nouveau champ de recherches sur la faune aquatique souterraine. CR Acad. Sci. 1967, 265, 369–370. [Google Scholar]
- Boulton, A.J.; Dole-Olivier, M.-J.; Marmonier, P. Optimizing a sampling strategy for assessing hyporheic invertebrate biodiversity using the Bou-Rouch method: Within site replication and sample volume. Archiv. Hydrobiol. 2003, 156, 431–456. [Google Scholar] [CrossRef]
- Kibichii, S.; Baars, J.R.; Kelly-Quinn, M. Optimising sample volume and replicates using the Bou-Rouch method for the rapid assessment of hyporheic fauna. Mar. Freshw. Res. 2009, 60, 83–96. [Google Scholar] [CrossRef]
- Pretty, J.L.; Hilldrew, A.G.; Trimmer, M. Nutrient dynamics in relation to surface-groundwater hydrological exchange in a groundwater fed chalk stream. J. Hydrol. 2006, 330, 84–100. [Google Scholar] [CrossRef]
- Davy-Bowker, J.-D.; Sweeting, W.; Wright, N.; Clark, R.T.; Arnott, S. The distribution of benthic and hyporheic macroinvertebrates from the heads and tails of riffles. Hydrobiologia 2006, 563, 109–123. [Google Scholar] [CrossRef]
- Pacioglu, O.; Robertson, A.L. The invertebrate community of the chalk stream hyporheic zone: Spatio-temporal distribution patterns. Knowl. Manag. Aquat. Ecosyst. 2017, 418, 10. [Google Scholar] [CrossRef] [Green Version]
- Murray-Bligh, J.A.D.; Furse, M.T.; Jones, F.H.; Gunn, R.J.M.; Dines, R.A.; Wright, J.F. Procedure for Collecting and Analysing Macroinvertebrate Samples for RIVPACS; Joint publication by the Institute of Freshwater Ecology and the Environment Agency; Institute of Freshwater Ecology and the Environment Agency: Wareham, UK, 1997; p. 162. [Google Scholar]
- Tachet, H.; Richoux, P.; Bournaud, M.; Usseglio-Polatera, P. Invertebres d’eau douce. Systematique, biologie, ecologie. CNRS Editions: Paris, France, 2010; p. 607. [Google Scholar]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef] [Green Version]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Zou, Y.; Axmacher, J.C. The Chord-Normalized Expected Species Shared (CNESS)-distance represents a superior measure of species turnover patterns. Methods Ecol. Evol. 2019, 11, 273–280. [Google Scholar] [CrossRef]
- Hancock, P. Human impacts on the stream–groundwater exchange zone. Environ. Manag. 2002, 29, 761–781. [Google Scholar] [CrossRef]
- Rillig, M.P.; Ziersch, L.; Hempel, S. Microplastic transport in soil by earthworms. Sci. Rep. 2017, 7, 1362. [Google Scholar] [CrossRef] [PubMed]
- Re, V. Shedding light on the invisible: Addressing the potential for groundwater contamination by plastic microfibers. Hydrogeol. J. 2019, 27, 2719–2727. [Google Scholar] [CrossRef] [Green Version]
- Tullos, D.; Penrose, D.; Jennings, G.; Cope, W. Analysis of functional traits in reconfigured channels: Implications for the bioassessment and disturbance of river restoration. J. N. Am. Benthol. Soc. 2009, 28, 80–92. [Google Scholar] [CrossRef]
- Franken, R.J.M.; Storey, R.G.; Dudley Williams, D. Biological, chemical and physical characteristics of downwelling and upwelling zones in the hyporheic zone of a north-temperate stream. Hydrobiologia 2001, 444, 183–195. [Google Scholar] [CrossRef]
- Sear, D.A.; Armitage, P.D.; Dawson, F.H. Groundwater dominated rivers. Hydrol. Process. 1999, 13, 255–276. [Google Scholar] [CrossRef]
- Kasahara, T.; Hill, A.R. Instream restoration: Its effectson lateral stream-subsurface water exchange in urban and agricultural streams in Southern Ontario. River Res. Appl. 2007, 23, 801–814. [Google Scholar] [CrossRef]
- Rubin, Z.; Kondolf, G.M.; Rios-Touma, B. Evaluating Stream Restoration Projects: What Do We Learn from Monitoring? Water 2017, 9, 174. [Google Scholar] [CrossRef] [Green Version]
- Bernhardt, E.S.; Palmer, M.; Allan, J.D.; Alexander, G.; Barnas, K.; Brooks, S.; Carr, J.; Clayton, S.; Dahm, C.; Follstad-Shah, J.; et al. Synthesizing US river restoration efforts. Science 2005, 308, 636–637. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, E.S.; Sudduth, E.B.; Palmer, M.A.; Allan, J.D.; Meyer, J.L.; Alexander, G.; Follastad-Shah, J.; Hassett, B.; Jenkinson, R.; Lave, R.; et al. Restoring rivers one reach at a time: Results from a survey of US river restoration practitioners. Restor. Ecol. 2007, 15, 482–493. [Google Scholar] [CrossRef] [Green Version]
- England, J.; Naura, M.; Mant, J.; Skinner, K. Seeking river restoration appraisal best practice: Supporting wider national and international environmental goals. Water Environ. J. 2019, 34, 1003–1011. [Google Scholar] [CrossRef]
- Louhi, P.; Mykrä, H.; Paavola, R.; Huusko, A.; Vehanen, T.; Mäki-Petäys, A.; Muotka, T. Twenty years of stream restoration in Finland: Little response by benthic macroinvertebrate communities. Ecol. Appl. 2011, 21, 1950–1961. [Google Scholar] [CrossRef]
- Haase, P.; Hering, D.; Jahnig, S.C.; Lorenz, A.W.; Sundermann, A. The impact of hydromorphological restoration on river ecological status: A comparison of fish, benthic invertebrates and macrophytes. Hydrobiologica 2013, 704, 475–488. [Google Scholar] [CrossRef]
- Sarriquet, P.E.; Bordenave, P.; Marmonier, P. Effects of bottom sediment restoration on interstitial habitat characteristics and benthic macroinvertebrate assemblages in a headwater stream. River Res. Appl. 2007, 23, 815–828. [Google Scholar] [CrossRef]
- Knott, J.; Mueller, M.; Pander, J.; Geist, J. Effectiveness of catchment erosion protection measures and scale-dependent response of stream biota. Hydrobiologia 2019, 830, 77–92. [Google Scholar] [CrossRef]
- Tonkin, J.D.; Stoll, S.; Sundermann, A.; Haase, P. Dispersal distance and the pool of taxa, but not barriers, determine the colonisation of restored river reaches by benthic invertebrates. Freshwat. Biol. 2014, 59, 1843–1855. [Google Scholar] [CrossRef]
- Palmer, M.; Ruhi, A. Linkages between flow regime, biota, and ecosystem processes: Implications for river restoration. Science 2019, 365, eaaw2087. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Period | Year | Reach | Individuals | Richness_Rarerified | 95% CI | Richness_Extrapolated | 95% CI | Dissimilarity_Min | Dissimilarity_Max | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hyporheos | Before | 2013 | Control | 519 | 21.00 | 16.92 | to | 25.08 | 26.83 | 17.33 | to | 36.33 | 0.06 | 0.57 |
| Restored | 1279 | 25.29 | 22.00 | to | 28.58 | 36.00 | 30.71 | to | 41.29 | |||||
| After | 2015 | Control | 1037 | 34.72 | 31.46 | to | 37.97 | 41.00 | 36.71 | to | 45.29 | 0.75 | 0.57 | |
| Restored | 610 | 26.00 | 21.00 | to | 31.00 | 32.80 | 24.34 | to | 41.25 | |||||
| After | 2016 | Control | 1026 | 40.00 | 34.16 | to | 45.85 | 55.92 | 40.79 | to | 71.06 | 0.25 | 0.47 | |
| Restored | 2906 | 45.60 | 42.09 | to | 49.12 | 64.00 | 55.77 | to | 72.23 | |||||
| After | 2018 | Control | 2173 | 15.11 | 12.74 | to | 17.47 | 21.00 | 16.85 | to | 25.15 | 0.16 | 0.51 | |
| Restored | 993 | 24.00 | 18.20 | to | 29.80 | 32.31 | 21.64 | to | 42.97 | |||||
| Benthos | Before | 2013 | Control | 5537 | 37.08 | 33.61 | to | 40.55 | 46.00 | 40.99 | to | 51.01 | 0.11 | 0.45 |
| Restored | 2733 | 40.00 | 35.39 | to | 44.61 | 47.18 | 38.06 | to | 56.29 | |||||
| After | 2015 | Control | 4148 | 48.31 | 44.10 | to | 52.52 | 50.00 | 44.90 | to | 55.10 | 0.13 | 0.47 | |
| Restored | 3569 | 51.00 | 44.45 | to | 57.55 | 53.58 | 46.85 | to | 60.32 | |||||
| After | 2016 | Control | 2737 | 36.95 | 35.17 | to | 38.73 | 37.00 | 35.22 | to | 38.78 | 0.39 | 0.41 | |
| Restored | 2475 | 35.00 | 33.01 | to | 36.99 | 35.19 | 32.67 | to | 37.72 | |||||
| After | 2018 | Control | 8419 | 67.40 | 62.58 | to | 72.23 | 73.00 | 67.48 | to | 78.52 | 0.13 | 0.41 | |
| Restored | 5898 | 68.00 | 62.48 | to | 73.52 | 72.45 | 65.41 | to | 79.50 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robertson, A.L.; Perkins, D.M.; England, J.; Johns, T. Invertebrate Responses to Restoration across Benthic and Hyporheic Stream Compartments. Water 2021, 13, 996. https://doi.org/10.3390/w13070996
Robertson AL, Perkins DM, England J, Johns T. Invertebrate Responses to Restoration across Benthic and Hyporheic Stream Compartments. Water. 2021; 13(7):996. https://doi.org/10.3390/w13070996
Chicago/Turabian StyleRobertson, Anne L., Daniel M. Perkins, Judy England, and Tim Johns. 2021. "Invertebrate Responses to Restoration across Benthic and Hyporheic Stream Compartments" Water 13, no. 7: 996. https://doi.org/10.3390/w13070996
APA StyleRobertson, A. L., Perkins, D. M., England, J., & Johns, T. (2021). Invertebrate Responses to Restoration across Benthic and Hyporheic Stream Compartments. Water, 13(7), 996. https://doi.org/10.3390/w13070996