
Microbial Detoxification of Dimethoate and Methomyl Residues in Aqueous Media

 and
and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental
2.1.1. Chemicals
2.1.2. Growth Medium
2.1.3. Isolation by Enrichment Culture
2.1.4. Identification of the Isolated Microbes
2.1.5. Determination of Microbial Growth
2.1.6. Biodegradation of the Tested Insecticides
2.1.7. HPLC Analysis
2.1.8. Calculation of Biodegradation Rate and Half-Life Time
2.2. Toxicity Test
2.2.1. Ethical Statement
2.2.2. Animals
2.2.3. Animal Treatment
2.2.4. Histopathological Examination
2.2.5. Biochemical Assays
2.2.6. Statistical Analysis
3. Results
3.1. Identification of the Tested Microbes
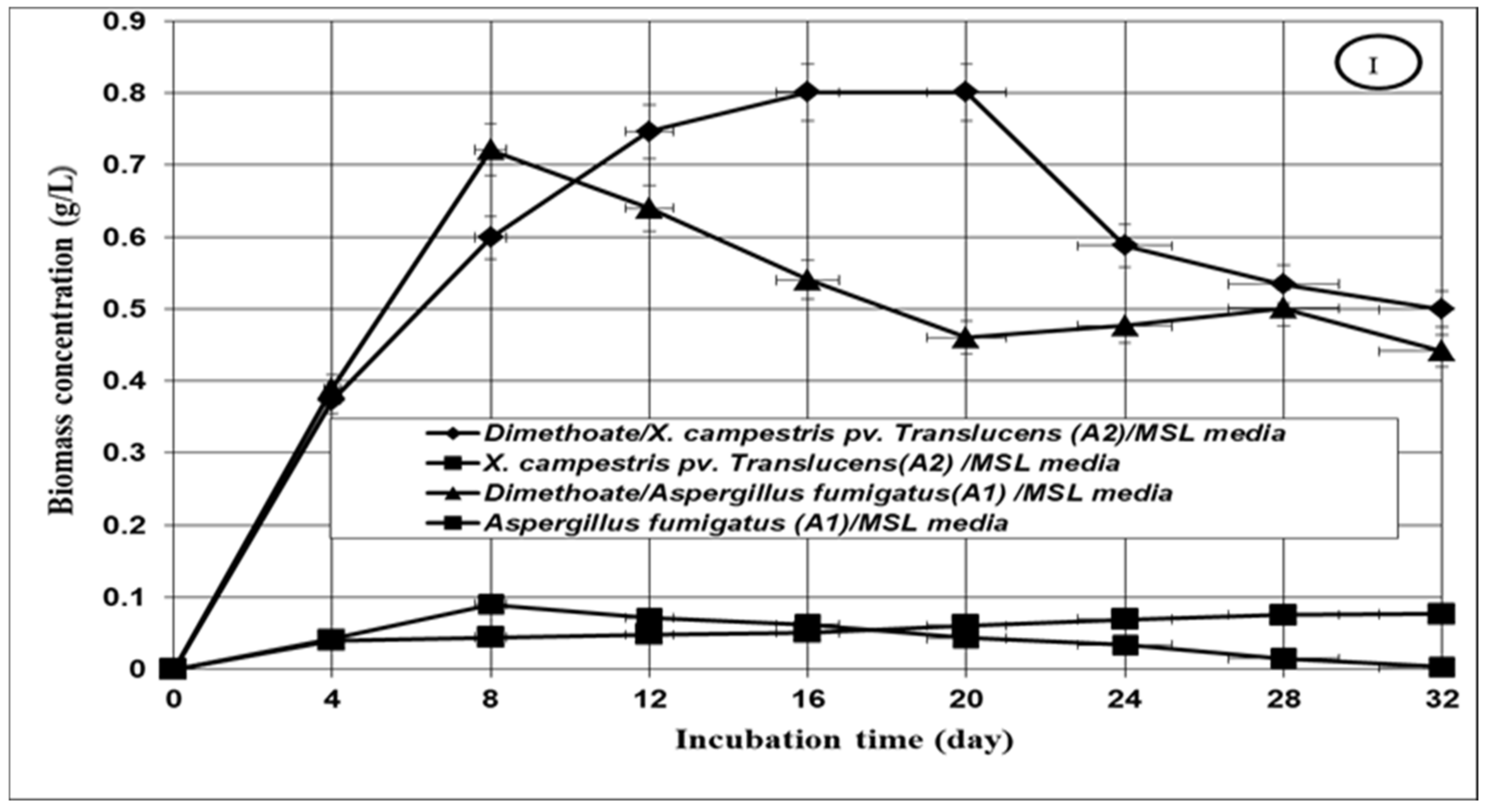
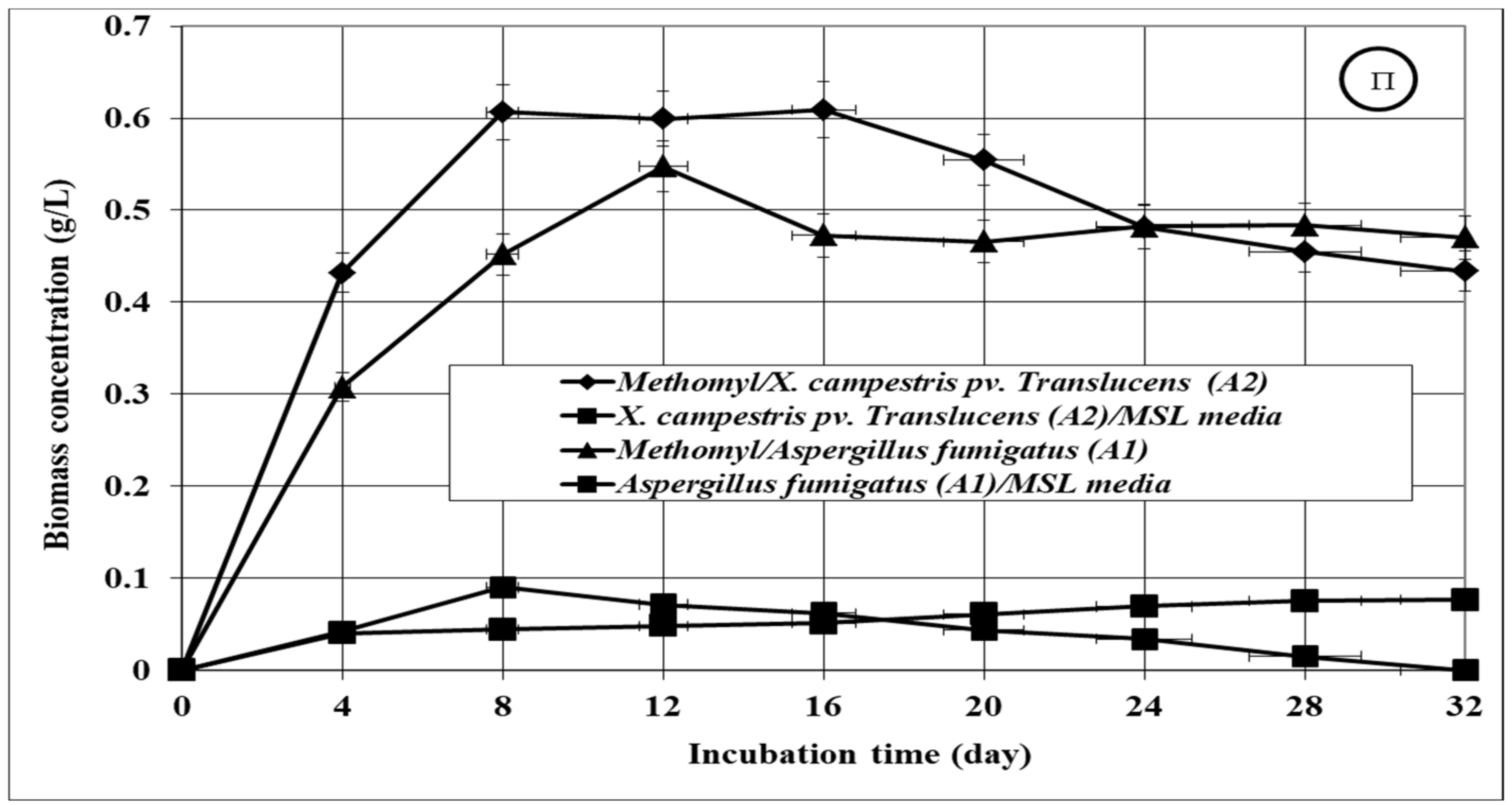
3.2. Growth of the Tested Microbial Isolates
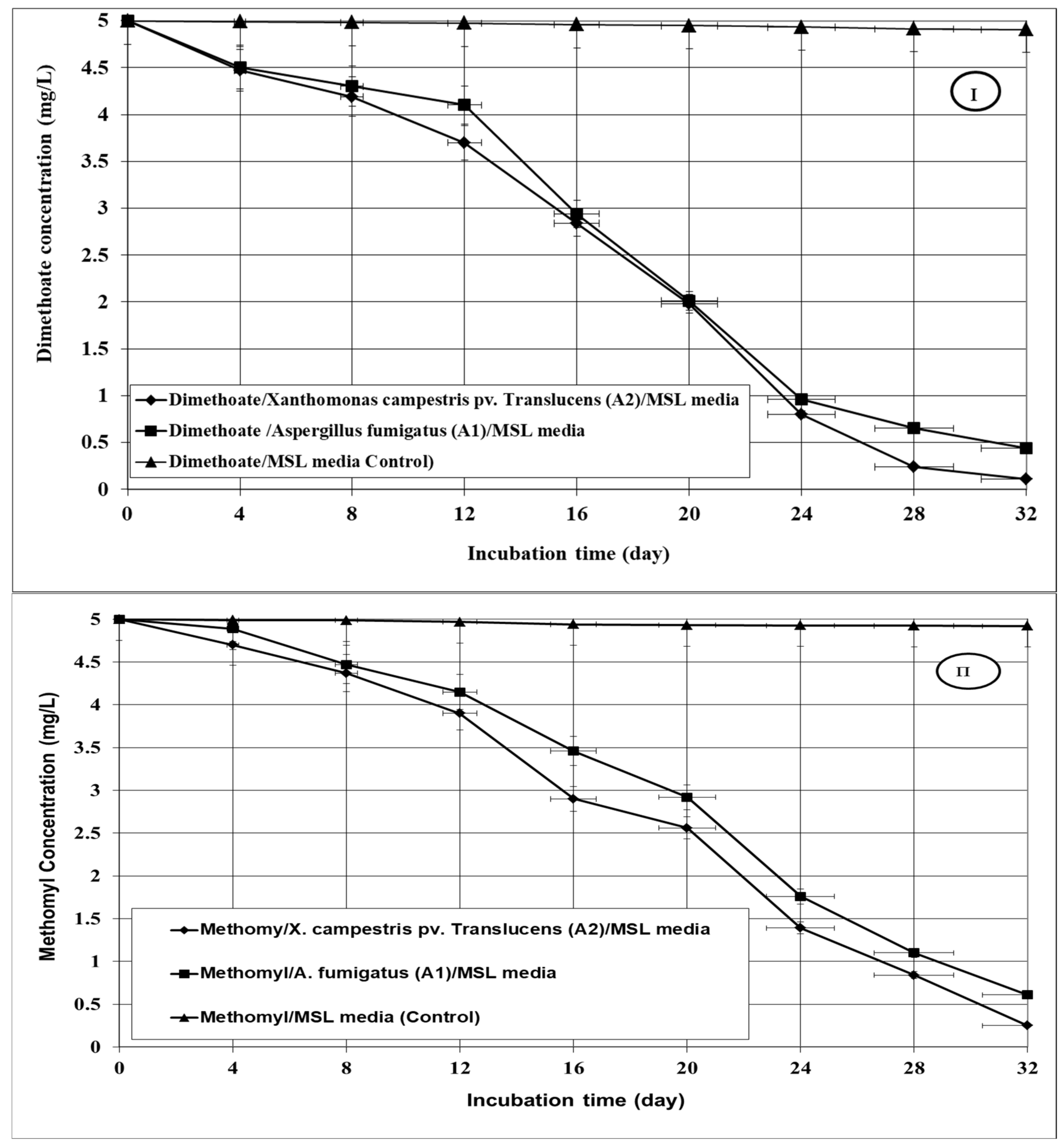
3.3. Bioremediation of the Tested Insecticides in Aqueous Media
3.4. Toxicity Assessment
3.4.1. Biochemical Parameters
3.4.2. Histopathological Changes
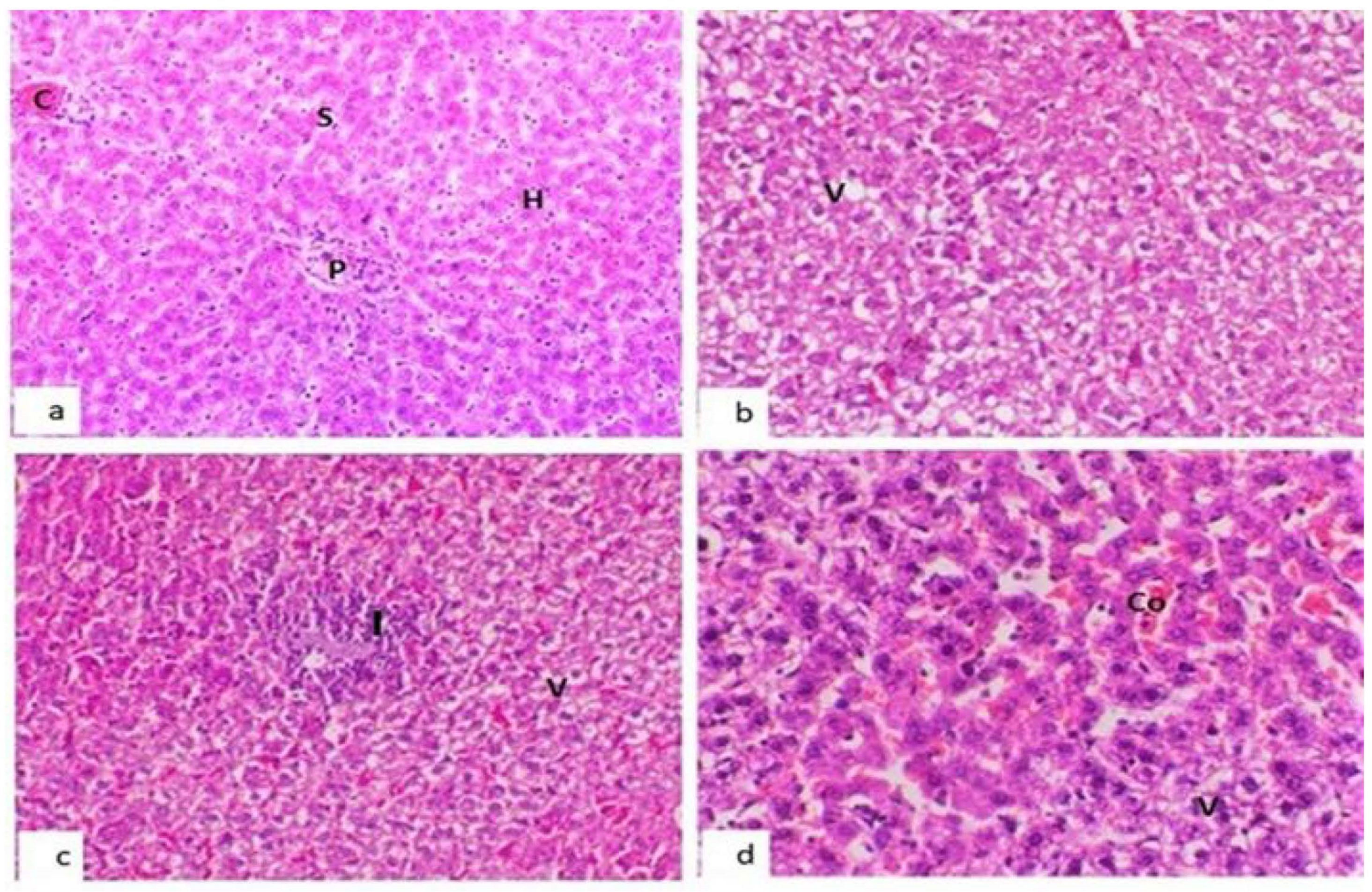
- LiverThe liver of the control group showed a normal hepatic lobule with a centrally located central vein and hepatocytes arranged in cords separated from each other by hepatic sinusoids (Figure 3a). However, the liver of group 1 showed the same morphology as that in the control group but with slight hepatocellular cytoplasmic vacuolation (Figure 3b), while the liver of group 2 showed moderate hepatocellular cytoplasmic hydropic degeneration and focal portal mononuclear cell infiltration (Figure 3c), and the liver of group 3 showed slight sinusoidal congestion and hepatocellular vacuolation (Figure 3d).
- KidneyThe normal structure of kidney tissue with an intact renal corpuscle and renal tubules in the control group is shown in Figure 4a. Kidneys in both groups 1 and 2 showed almost the same histologic structure as that in the control group except slight changes in the form of renal tubule epithelial cell swelling in group 1 (Figure 4b) and the presence of homogeneous acidophilic material in the lumen of renal tubules in group 2 (Figure 4c). The kidneys of group 3 showed mild vacuolation in the epithelial lining of renal tubules (Figure 4d).
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Directive 60/EC of the European Parliament and of the Council on the assessment and management of flood risks. Off. J. Eur. Union 2007, 1, 186–193.
- European Comission. Water Scarcity and Drought—In Depth Assessment; European Comission: Luxembourg, 2006. [Google Scholar]
- Campo, J.; Masiá, A.; Blasco, C.; Picó, Y. Occurrence and removal efficiency of pesticides in sewage treatment plants of four Mediterranean River Basins. J. Hazard. Mater. 2013, 263, 146–157. [Google Scholar] [CrossRef]
- Castillo, M.D.P.; Torstensson, L.; Stenström, J. Biobeds for Environmental Protection from Pesticide Use—A Review. J. Agric. Food Chem. 2008, 56, 6206–6219. [Google Scholar] [CrossRef]
- Omirou, M.; Dalias, P.; Costa, C.; Papastefanou, C.; Dados, A.; Ehaliotis, C.; Karpouzas, D. Exploring the potential of biobeds for the depuration of pesticide-contaminated wastewaters from the citrus production chain: Laboratory, column and field studies. Environ. Pollut. 2012, 166, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Karanasios, E.; Tsiropoulos, N.G.; Karpouzas, D.G. On-farm biopurification systems for the depuration of pesticide wastewaters: Recent biotechnological advances and future perspectives. Biodegradation 2012, 23, 787–802. [Google Scholar] [CrossRef]
- Sniegowski, K.; Springael, D. Establishment of multiple pesticide biodegradation capacities from pesticide-primed materials in on-farm biopurification system microcosms treating complex pesticide-contaminated wastewater. Pest Manag. Sci. 2015, 71, 986–995. [Google Scholar] [CrossRef]
- Deshpande, N.; Sarnaik, S.; Paranjpe, S.; Kanekar, P. Optimization of dimethoate degradation by Brevundimonas sp. MCM B-427 using factorial design: Studies on interactive effects of environmental factors. World J. Microbiol. Biotechnol. 2004, 20, 455–462. [Google Scholar] [CrossRef]
- Pappas, C.J.; Kyriakidis, N.V. A comparison of dimethoate degradation in lemons and mandarins on the trees with two GC systems. Food Chem. 2003, 80, 23–28. [Google Scholar] [CrossRef]
- Hassal, A.K. Organophosphorous insecticides. In The Biochemistry and Uses; Wiley-VCH: Weinhem, Germany, 1990. [Google Scholar]
- López-Carillo, L.; López-Cervantes, M. Effect of exposure to organophosphate pesticides on serum cholinesterase levels. Arch. Environ. Health Int. J. 1993, 48, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Al-Jaghbir, M.T.; Salhab, A.S.; Hamarsheh, F.A. Dermal and inhalation exposure to dimethoate. Arch. Environ. Contam. Toxicol. 1992, 22, 358–361. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, M.; Raizada, R. Development effect of technical dimethoate in rats: Maternal and fetal toxicity evaluation. Indian J. Exp. Biol. 1996, 34, 329–333. [Google Scholar] [PubMed]
- Andreozzi, R.; Ialongo, G.; Marotta, R.; Sanchirico, R. The thermal decomposition of dimethoate. J. Hazard. Mater. 1999, 64, 283–294. [Google Scholar] [CrossRef]
- Tomlin, D.S. The Pesticide Manual, 13th ed.; BCPC Publications: Alton, UK, 2003; Volume 1344. [Google Scholar]
- Strathmann, T.J.; Stone, A.T. Reduction of the carbamate pesticides oxamyl and methomyl by dissolved FeII and CuI. Environ. Sci. Technol. 2001, 35, 2461–2469. [Google Scholar] [CrossRef] [PubMed]
- Bhushan, R.; Thapar, S.; Mathur, R. Accumulation pattern of pesticides in tropical fresh waters. Biomed. Chromatogr. 1997, 11, 143–150. [Google Scholar] [CrossRef]
- Thapar, S.; Bhushan, R.; Mathur, R. Degradation of organophosphorus and carbamate pesticides in soils—HPLC determination. Biomed. Chromatogr. 1995, 9, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Yang, C.; Qu, H.; Liu, Z.; Fu, Q.; Qiao, C. Cloning of a novel aldo-keto reductase gene from Klebsiella sp. strain F51-1-2 and its functional expression in Escherichia coli. Appl. Environ. Microbiol. 2007, 73, 4959–4965. [Google Scholar] [CrossRef]
- DebMandal, M.; Mandal, S.; Pal, N.K.; Aich, A. Potential metabolites of dimethoate produced by bacterial degradation. World J. Microbiol. Biotechnol. 2008, 24, 69–72. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Chen, X.; Shi, Y.; Su, Z.C. Bacterial degradation of chlorpyrifos by Bacillus cereus. Adv. Mater. Res. 2011, 356–360, 676–680. [Google Scholar] [CrossRef]
- Chaudhry, G.R.; Ali, A. Bacterial metabolism of carbofuran. Appl. Environ. Microbiol. 1988, 54, 1414–1419. [Google Scholar] [CrossRef]
- Vincent, J. Manual for the Practical Study of Root-Nodule Bacteria; Blackwell of Oxford: Oxford, UK, 1970. [Google Scholar]
- Maheswari, N.; Ramya, P. Biodegradation of organophosphate and dimethoate by Aspergillus niger in Vigna mungo. Int. J. Univ. Pharm. Bio Sci. 2013, 2, 177–185. [Google Scholar]
- Ashry, M.; Bayoumi, O.; El-Fakharany, I.; Derbalah, A.; Ismail, A. Monitoring and removal of pesticides residues in drinking water collected from Kafr El-Sheikh governorate, Egypt. J. Agric. Res. Tanta Univ. 2006, 32, 691–704. [Google Scholar]
- Hassan, H. Fungal utilization of organophosphorus pesticides and their degradation by Aspergillus flavus and A. sydowii in soil. Folia Microbiol. 1999, 44, 77–84. [Google Scholar] [CrossRef]
- Subramanian, G.; Sekar, S.; Sampoornam, S. Biodegradation and utilization of organophosphorus pesticides by cyanobacteria. Int. Biodeterior. Biodegrad. 1994, 33, 129–143. [Google Scholar] [CrossRef]
- Gams, W.; Christensen, M.; Onions, A.H.; Pitt, J.I.; Samson, R.A. Infrageneric taxa of Aspergillus. In Advances in Penicillium and Aspergillus Systematics; Springer: Berlin/Heidelberg, Germany, 1986; pp. 55–62. [Google Scholar]
- Okuda, T.; Klich, M.; Seifert, K.; Ando, K.; Samson, R.; Pitt, J. Integration of Modern Taxonomic Methods for Penicillium and Aspergillus Classification; Hardwood Academic Publishers: Reading, UK, 2000; pp. 83–100. [Google Scholar]
- McClenny, N. Laboratory detection and identification of Aspergillus species by microscopic observation and culture: The traditional approach. Med. Mycol. 2005, 43, S125–S128. [Google Scholar] [CrossRef]
- Diba, K.; Kordbacheh, P.; Mirhendi, S.; Rezaie, S.; Mahmoudi, M. Identification of Aspergillus species using morphological characteristics. Pak. J. Med. Sci. 2007, 23, 867. [Google Scholar]
- Barragan-Huerta, B.E.; Costa-Pérez, C.; Peralta-Cruz, J.; Barrera-Cortés, J.; Esparza-García, F.; Rodríguez-Vázquez, R. Biodegradation of organochlorine pesticides by bacteria grown in microniches of the porous structure of green bean coffee. Int. Biodeterior. Biodegrad. 2007, 59, 239–244. [Google Scholar] [CrossRef]
- Li, R.; Zheng, J.; Wang, R.; Song, Y.; Chen, Q.; Yang, X.; Li, S.; Jiang, J. Biochemical degradation pathway of dimethoate by Paracoccus sp. Lgjj-3 isolated from treatment wastewater. Int. Biodeterior. Biodegrad. 2010, 64, 51–57. [Google Scholar] [CrossRef]
- Antachopoulos, C.; Meletiadis, J.; Roilides, E.; Sein, T.; Walsh, T.J. Rapid susceptibility testing of medically important zygomycetes by XTT assay. J. Clin. Microbiol. 2006, 44, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Antachopoulos, C.; Meletiadis, J.; Sein, T.; Roilides, E.; Walsh, T.J. Use of high inoculum for early metabolic signalling and rapid susceptibility testing of Aspergillus species. J. Antimicrob. Chemother. 2007, 59, 230–237. [Google Scholar] [CrossRef]
- Molina-Ramirez, C.; Castro, M.; Osorio, M.; Torres-Taborda, M.; Gomez, B.; Zuluaga, R.; Gomez, C.; Ganan, P.; Rojas, O.J.; Castro, C. Effect of Different Carbon Sources on Bacterial Nanocellulose Production and Structure Using the Low pH Resistant Strain Komagataeibacter Medellinensis. Materials 2017, 10, 639. [Google Scholar] [CrossRef] [PubMed]
- Derbalah, A.; Belal, E.; Massoud, A. Biodegradability of famoxadone by various microbial isolates in aquatic systems. Land Contam. Reclam. 2008, 16, 13. [Google Scholar] [CrossRef]
- Derbalah, A.; Ismail, A. Efficiency of different remediation technologies for fenitrothion and dimethoate removal in the aquatic system. Agrochimica 2012, 56, 234–246. [Google Scholar]
- Ismail, A.; Derbalah, A.; Shaheen, S. Monitoring and remediation technologies of organochlorine pesticides in drainage water. Pol. J. Chem. Technol. 2015, 17, 115–122. [Google Scholar] [CrossRef]
- Monkiedje, A.; Spiteller, M. Degradation of metalaxyl and mefenoxam and effects on the microbiological properties of tropical and temperate soils. Int. J. Environ. Res. Public Health 2005, 2, 272–285. [Google Scholar] [CrossRef] [PubMed]
- Tamimi, M.; Qourzal, S.; Assabbane, A.; Chovelon, J.-M.; Ferronato, C.; Ait-Ichou, Y. Photocatalytic degradation of pesticide methomyl: Determination of the reaction pathway and identification of intermediate products. Photochem. Photobiol. Sci. 2006, 5, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Korsrud, G.O.; Grice, H.C.; McLaughlan, J.M. Sensitivity of several serum enzymes in detecting carbon tetrachloride-induced liver damage in rats. Toxicol. Appl. Pharmacol. 1972, 22, 474–483. [Google Scholar] [CrossRef]
- Derbalah, A.; Ismail, A. Remediation technologies of diazinon and malathion residues in aquatic system. Environ. Prot. Eng. 2013, 39, 135–147. [Google Scholar] [CrossRef]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques; Elsevier Health Sciences: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Waber, H.; Dtsch, M. Cholinestrase kinetic colorimetric method. Dtsch. Med. Wschr. 1966, 91, 1927. [Google Scholar]
- Azazi, I.; ALazab, A.; Nassrallah, M. Influence of Photoperiod and Crude Protein Levels on Productive Traits and Economic Efficiency of Muscovy Duckling Males During Summer Season. J. Product. Dev. 2016, 21, 301–321. [Google Scholar] [CrossRef]
- Reitman, A.; Frankel, S.A. GPT (ALT) Glutamic-Pyruvic Transaminase and GOT (AST) Glutamic -Pyruvic Transaminase colorimetric method. Amer. J. Clin. Path. 1957, 28, 56. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Singleton, I.; Mathew, Z. Fungal remediation of soils contaminated with persistent organic pollutants. In Fungi in Bioremediation; British Mycological Society Symposium Series; Cambridge University Press: Cambridge, UK, 2001; pp. 79–96. [Google Scholar]
- Bhalerao, T.S.; Puranik, P.R. Microbial degradation of monocrotophos by Aspergillus oryzae. Int. Biodeterior. Biodegrad. 2009, 63, 503–508. [Google Scholar] [CrossRef]
- Madhuri, J. Degradation of dimethoate by cellulolytic bacteria in cotton soils. Glob. J. Med. Res. Microbiol. Pathol. 2014, 14, 9–12. [Google Scholar]
- Mohamed, M.S. Degradation of methomyl by the novel bacterial strain Stenotrophomonas maltophilia M1. Electron. J. Biotechnol. 2009, 12, 6–7. [Google Scholar] [CrossRef]
- Lone, M.A.; Wani, M.R. Degradation of dimethoate and pyrethroid by using fungal strains isolated from the rhizosphere of Juglansregia L. in the northern region of Jammu and Kashmir, India. Int. J. Pharma Bio Sci. 2012, 3, 716–723. [Google Scholar]
- Yin, X.; Lian, B. Dimethoate degradation and calcium phosphate formation induced by Aspergillus niger. Afr. J. Microbiol. Res. 2012, 6, 7603–7609. [Google Scholar] [CrossRef]
- Pandey, P.; Khillare, P.; Kumar, K. Assessment of organochlorine pesticide residues in the surface sediments of River Yamuna in Delhi, India. J. Environ. Prot. 2011, 2, 511. [Google Scholar] [CrossRef]
- Hernández, A.F.; Gil, F.; Lacasaña, M.; Rodríguez-Barranco, M.; Gómez-Martin, A.; Lozano, D.; Pla, A. Modulation of the endogenous antioxidants paraoxonase-1 and urate by pesticide exposure and genetic variants of xenobiotic-metabolizing enzymes. Food Chem. Toxicol. 2013, 61, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Lei, A.-P.; Wong, Y.-S.; Tam, N.F.-Y. Pyrene-induced changes of glutathione-S-transferase activities in different microalgal species. Chemosphere 2003, 50, 293–301. [Google Scholar] [CrossRef]
- Pflugmacher, S.; Wiegand, C.; Oberemm, A.; Beattie, K.A.; Krause, E.; Codd, G.A.; Steinberg, C.E. Identification of an enzymatically formed glutathione conjugate of the cyanobacterial hepatotoxin microcystin-LR: The first step of detoxication. Biochim. Biophys. Acta BBA Gen. Subj. 1998, 1425, 527–533. [Google Scholar] [CrossRef]
- Banaee, M.; Sureda, A.; Mirvaghefi, A.; Ahmadi, K. Effects of diazinon on biochemical parameters of blood in rainbow trout (Oncorhynchus mykiss). Pestic. Biochem. Physiol. 2011, 99, 1–6. [Google Scholar] [CrossRef]
- Yousef, M.I.; El-Demerdash, F.; Kamel, K.; Al-Salhen, K. Changes in some hematological and biochemical indices of rabbits induced by isoflavones and cypermethrin. Toxicology 2003, 189, 223–234. [Google Scholar] [CrossRef]
- Bernet, D.; Schmidt, H.; Meier, W.; Burkhardt-Holm, P.; Wahli, T. Histopathology in fish: Proposal for a protocol to assess aquatic pollution. J. Fish Dis. 1999, 22, 25–34. [Google Scholar] [CrossRef]
- Dutta, H.; Adhikari, S.; Singh, N.; Roy, P.; Munshi, J. Histopathological changes induced by malathion in the liver of a freshwater catfish, Heteropneustes fossilis (Bloch). Bull. Environ. Contam. Toxicol. 1993, 51, 895–900. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, E.D.L.; Fanta, E. Liver histopathology of the fish Brachydanio rerio Hamilton-Buchman after acute exposure to sublethal levels of the organophosphate Dimethoate 500. Rev. Bras. Zool. 1998, 15, 441–450. [Google Scholar] [CrossRef]
- Wafa, T.; Amel, N.; Issam, C.; Imed, C.; Abdelhedi, M.; Mohamed, H. Subacute effects of 2, 4-dichlorophenoxyacetic herbicide on antioxidant defense system and lipid peroxidation in rat erythrocytes. Pestic. Biochem. Physiol. 2011, 99, 256–264. [Google Scholar] [CrossRef]
- El-Demerdash, F.; Dewer, Y.; ElMazoudy, R.H.; Attia, A.A. Kidney antioxidant status, biochemical parameters and histopathological changes induced by methomyl in CD-1 mice. Exp. Toxicol. Pathol. 2013, 65, 897–901. [Google Scholar] [CrossRef]
- Sharma, Y.; Bashir, S.; Irshad, M.; Nag, T.; Dogra, T. Dimethoate-induced effects on antioxidant status of liver and brain of rats following subchronic exposure. Toxicology 2005, 215, 173–181. [Google Scholar] [CrossRef]
- Henics, T.; Wheatley, D.N. Cytoplasmic vacuolation, adaptation and cell death: A view on new perspectives and features. Biol. Cell 1999, 91, 485–498. [Google Scholar] [CrossRef]
- Rahm, M.; Atty, Y.A.; Rahman, M.A.; Sabry, M. Structural changes induced by gibberellic acid in the renal cortex of adult male albino rats. MOJ Anat. Physiol. 2017, 3, 21–27. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Degradation Rate Constant (Day −1) | Half-Life (t1/2) (Days) | R2 |
|---|---|---|---|
| Dimethoate | |||
| X. campestris pv. Translucens | 0.0437 a | 15.86 a | 0.91 |
| A. fumigatus | 0.0420 a | 16.50 a | 0.84 |
| Methomyl | |||
| X. campestris pv. Translucens | 0.0351 a | 19.74 a | 0.93 |
| A. fumigatus | 0.0272 b | 25.48 b | 0.93 |
| Treatment | AChE (U/mL) | GPT (U/mL) | GOT (U/mL) | GST (U/mL) |
|---|---|---|---|---|
| Dimethoate | ||||
| D/X.c.pv. Translucens | 0.0091 ± 0.001 a | 19.03 ± 0.73 a | 20.28 ± 0.16 a | 0.0765 ± 0.001 a |
| X.c.pv. Translucens | 0.0091 ± 0.001 a | 19.04 ± 1.22 a | 20.22 ± 0.61 a | 0.0771 ± 0.001 a |
| Methomyl | ||||
| M/X. campestris pv. Translucens | 0.092 ± 0.0013 a | 19.03 ± 0.74 a | 19.93 ± 0.74 a | 0.0775 ± 0.001 a |
| X. campestris pv. Translucens | 0.091 ± 0.0011 a | 19.05 ± 1.25 a | 19.99 ± 1.25 a | 0.0791 ± 0.001 a |
| Control | 0.0091 ± 0.001 a | 19.04 ± 0.68 a | 20.29 ± 0.61 a | 0.0781 ± 0.001 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Derbalah, A.; Massoud, A.; El-Mehasseb, I.; Allah, M.S.; Ahmed, M.S.; Al-Brakati, A.; Elmahallawy, E.K. Microbial Detoxification of Dimethoate and Methomyl Residues in Aqueous Media. Water 2021, 13, 1117. https://doi.org/10.3390/w13081117
Derbalah A, Massoud A, El-Mehasseb I, Allah MS, Ahmed MS, Al-Brakati A, Elmahallawy EK. Microbial Detoxification of Dimethoate and Methomyl Residues in Aqueous Media. Water. 2021; 13(8):1117. https://doi.org/10.3390/w13081117
Chicago/Turabian StyleDerbalah, Aly, Ahmed Massoud, Ibrahim El-Mehasseb, Moustafa Saad Allah, Mohamed S. Ahmed, Ashraf Al-Brakati, and Ehab Kotb Elmahallawy. 2021. "Microbial Detoxification of Dimethoate and Methomyl Residues in Aqueous Media" Water 13, no. 8: 1117. https://doi.org/10.3390/w13081117
APA StyleDerbalah, A., Massoud, A., El-Mehasseb, I., Allah, M. S., Ahmed, M. S., Al-Brakati, A., & Elmahallawy, E. K. (2021). Microbial Detoxification of Dimethoate and Methomyl Residues in Aqueous Media. Water, 13(8), 1117. https://doi.org/10.3390/w13081117