
Food Web Responses to a Cyanobacterial Bloom in a Freshwater Eutrophic Lake


Abstract
:1. Introduction
2. Material and Methods
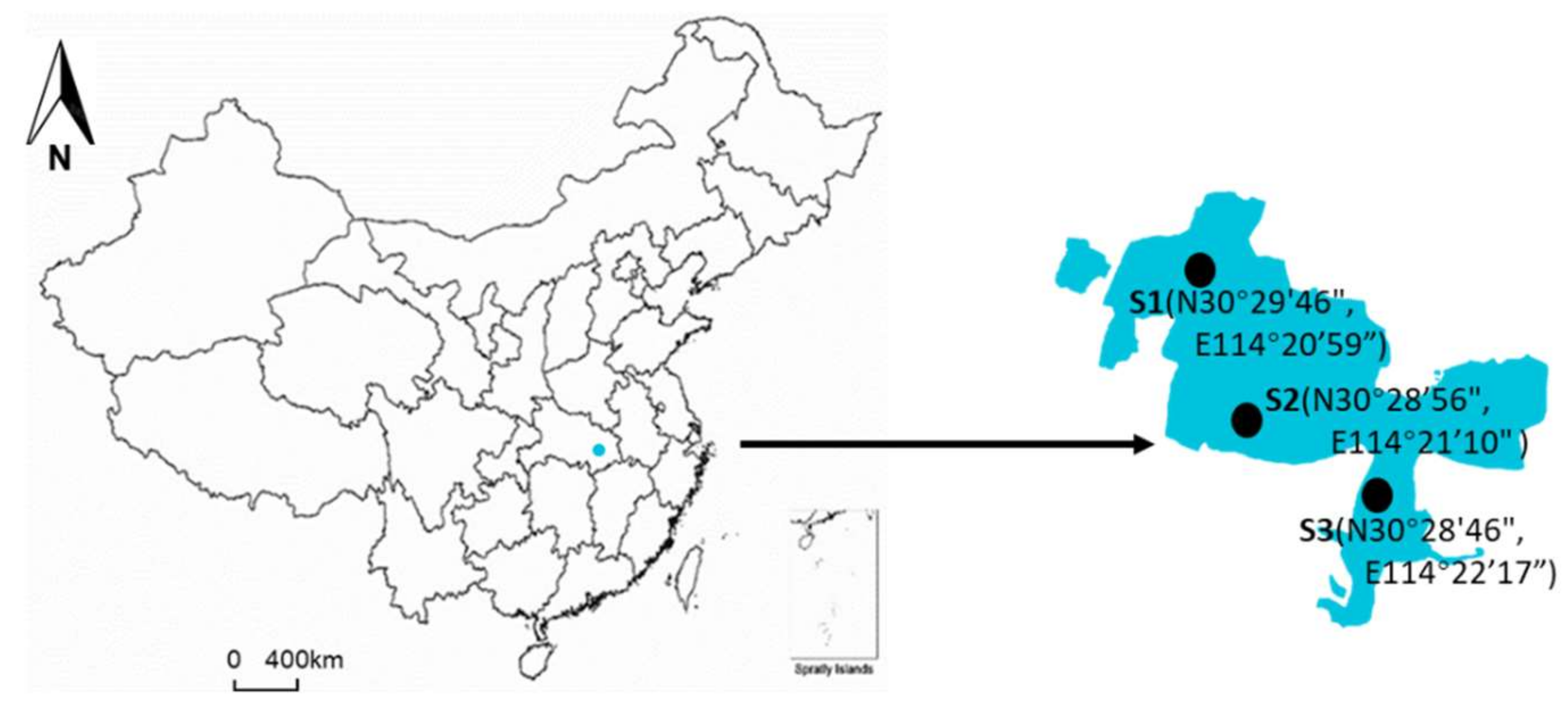
2.1. Field Observations and Sample Treatments
2.2. Analysis of Plankton Community Structure
2.3. Analysis of Bacterial Community Structure
2.4. Microzooplankton Grazing Experiments
2.5. HPLC Pigment Analysis
2.6. Date Analysis
3. Results
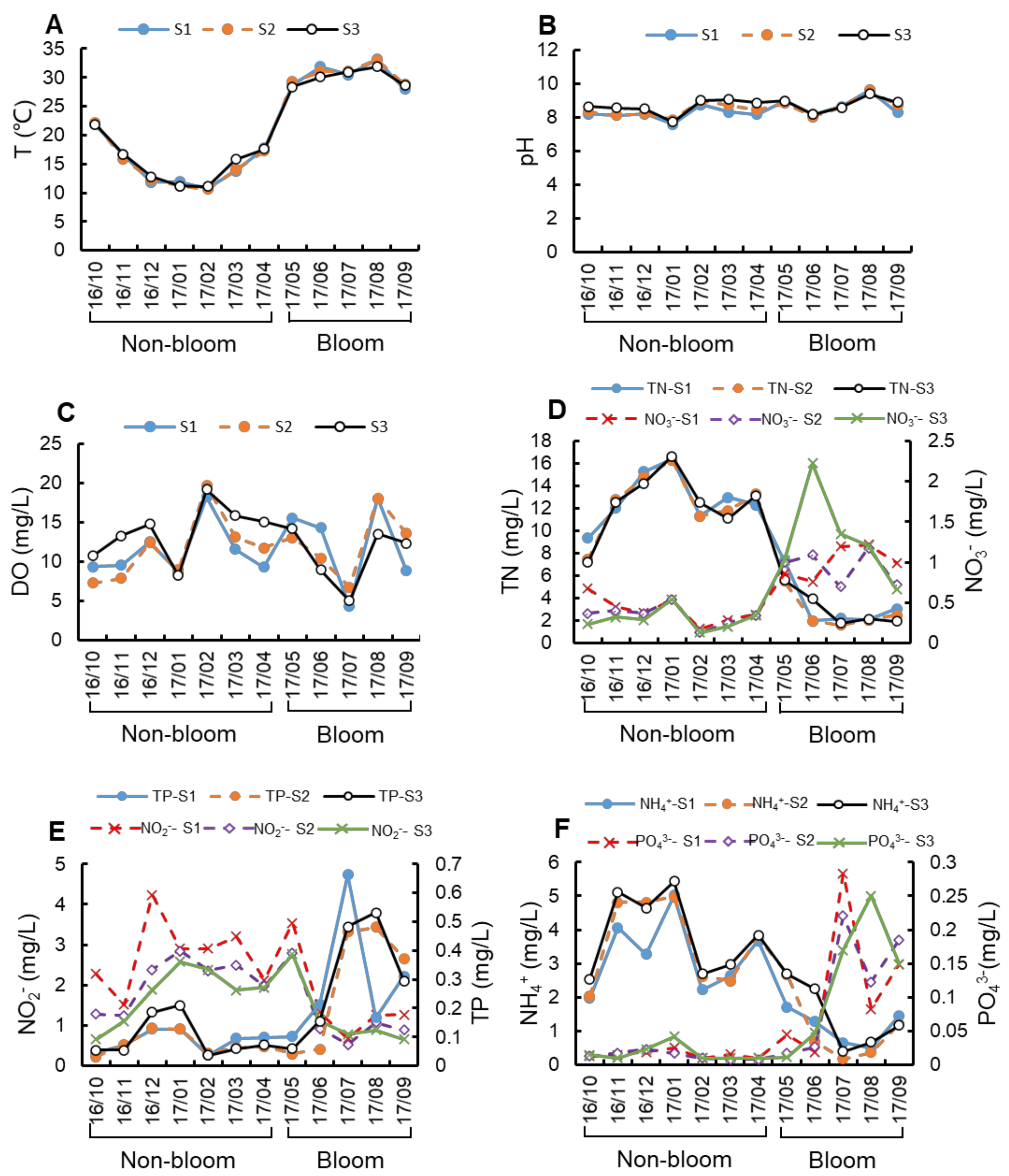
3.1. Environmental Conditions
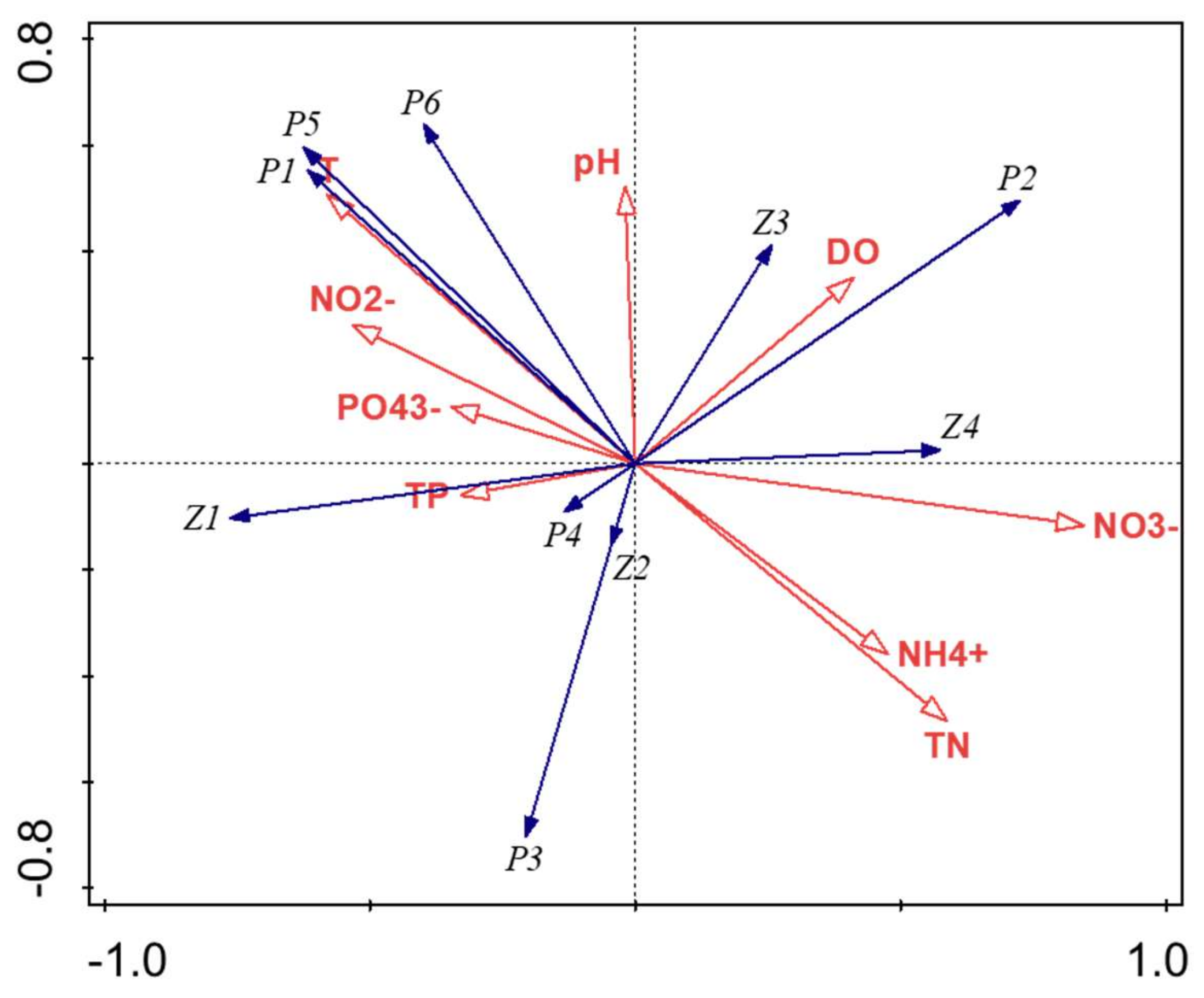
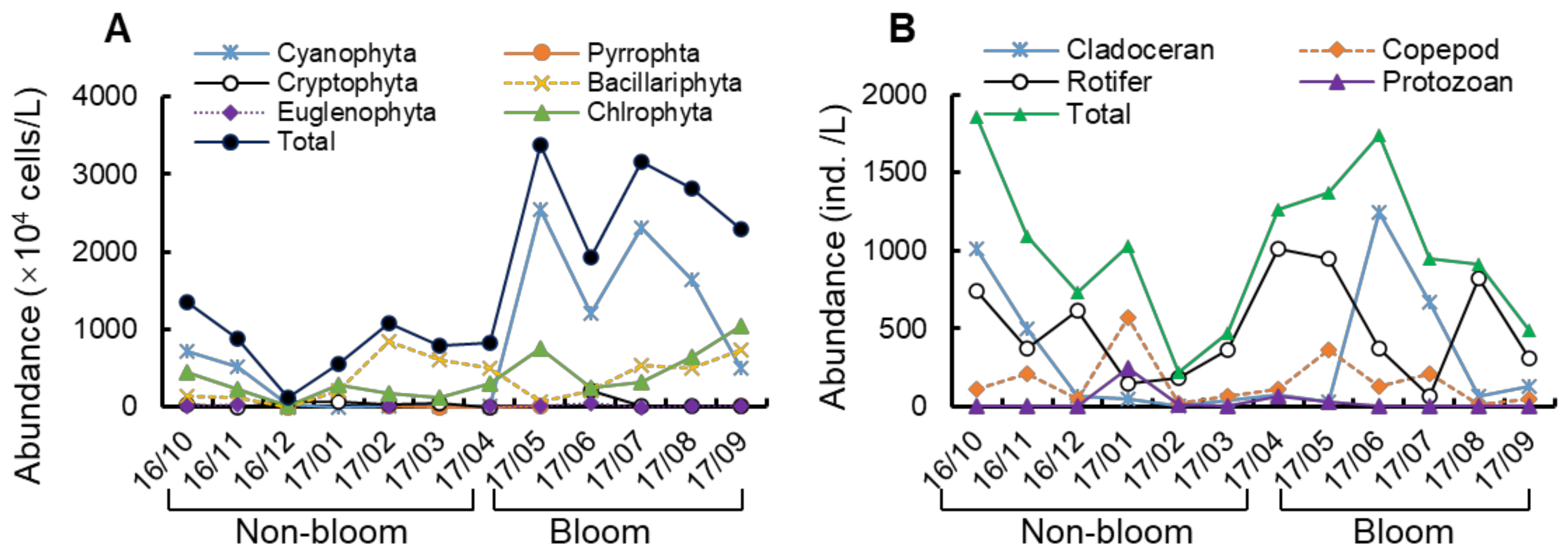
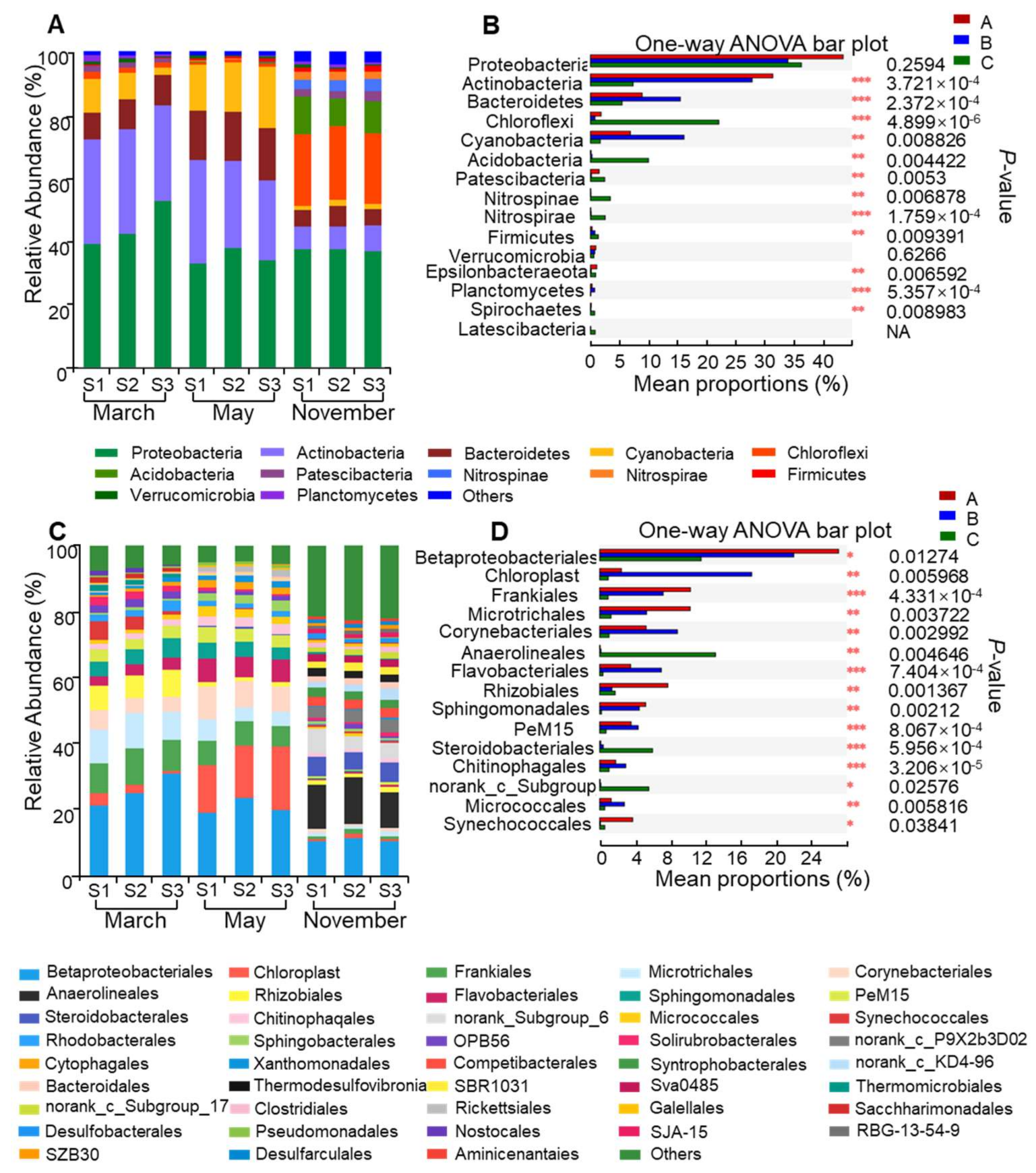
3.2. Variation in Plankton Community Composition
3.3. Variation in Bacterial Community Structure
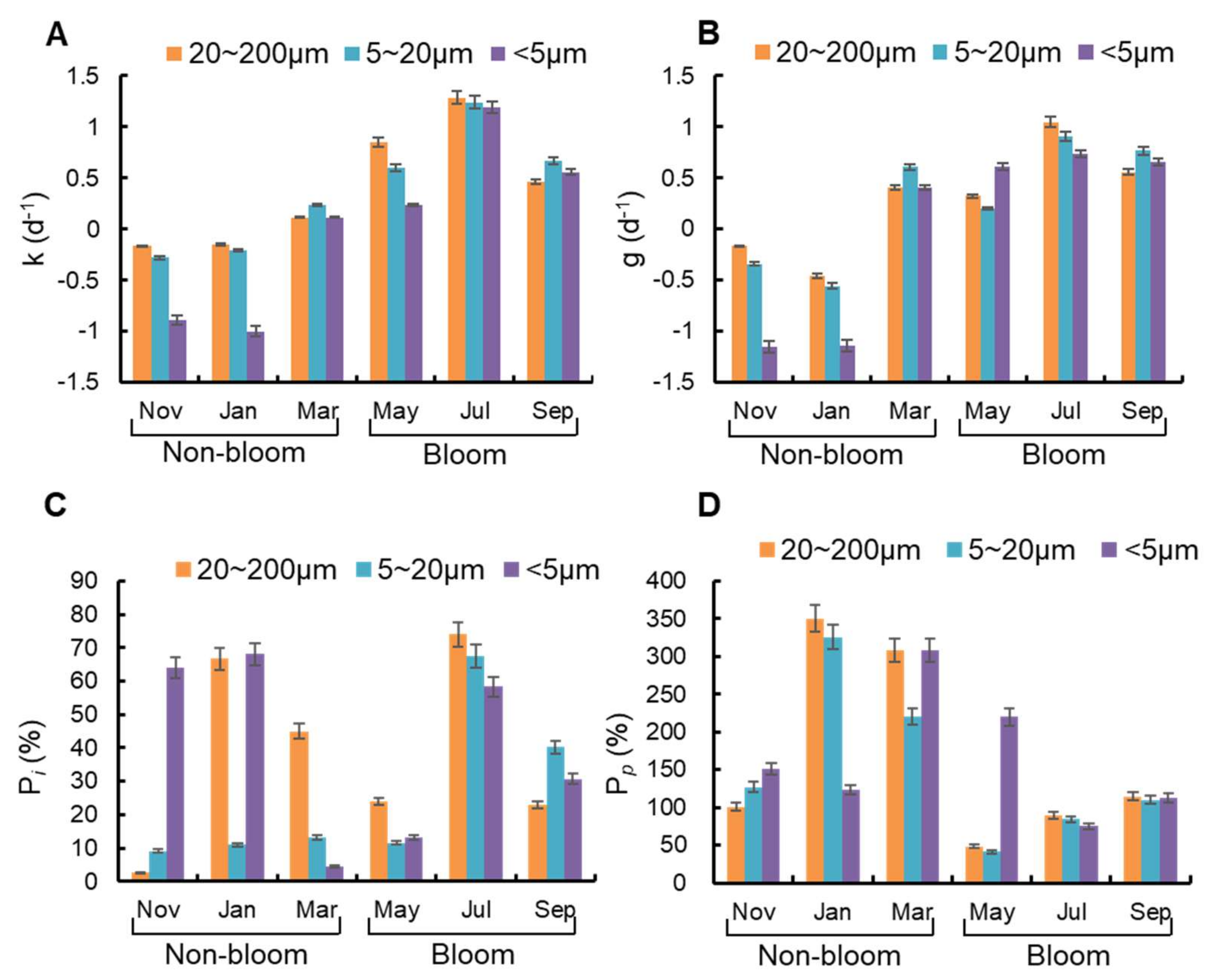
3.4. Microzooplankton Grazing on Size Specific Phytoplankton
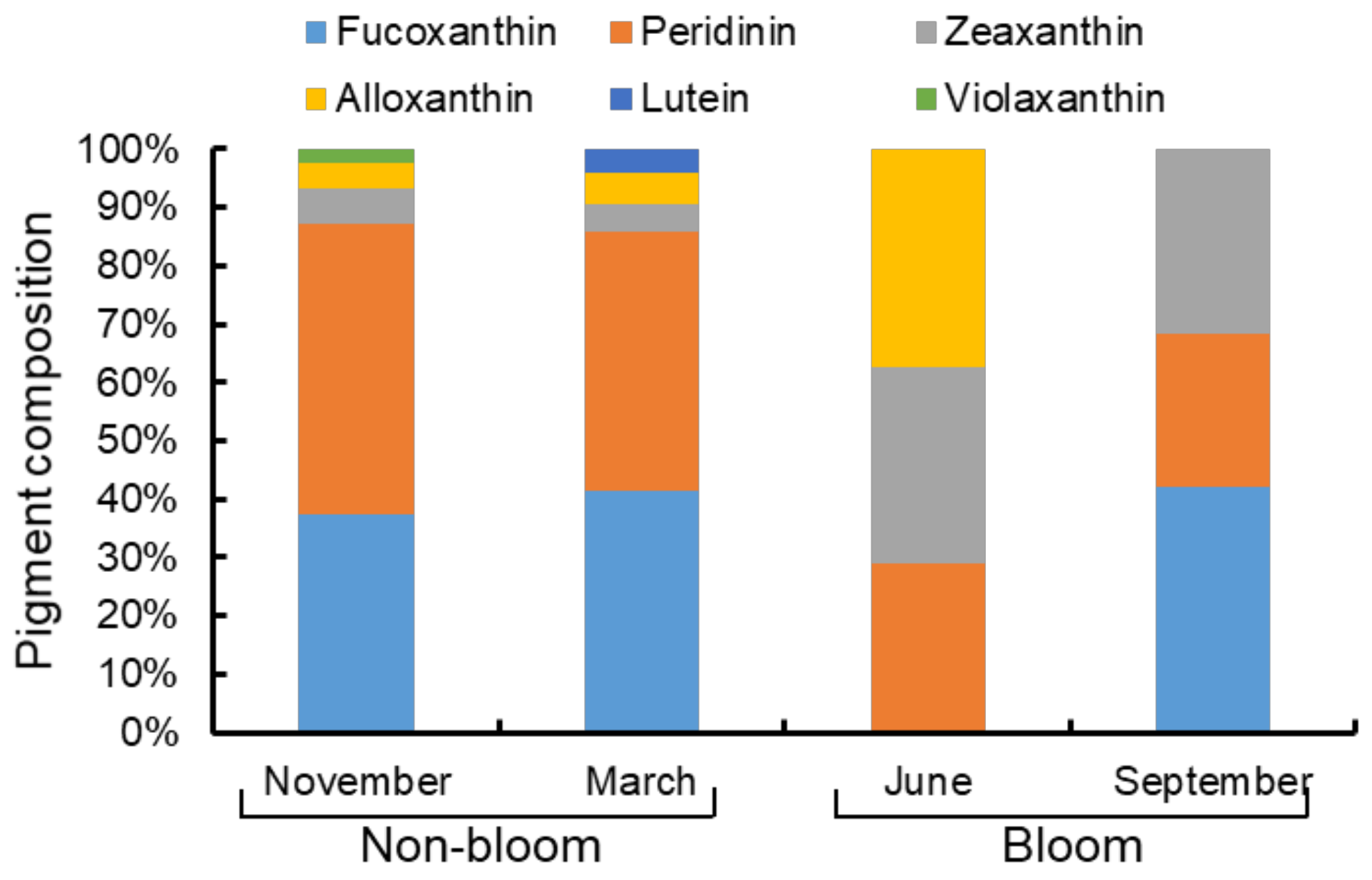
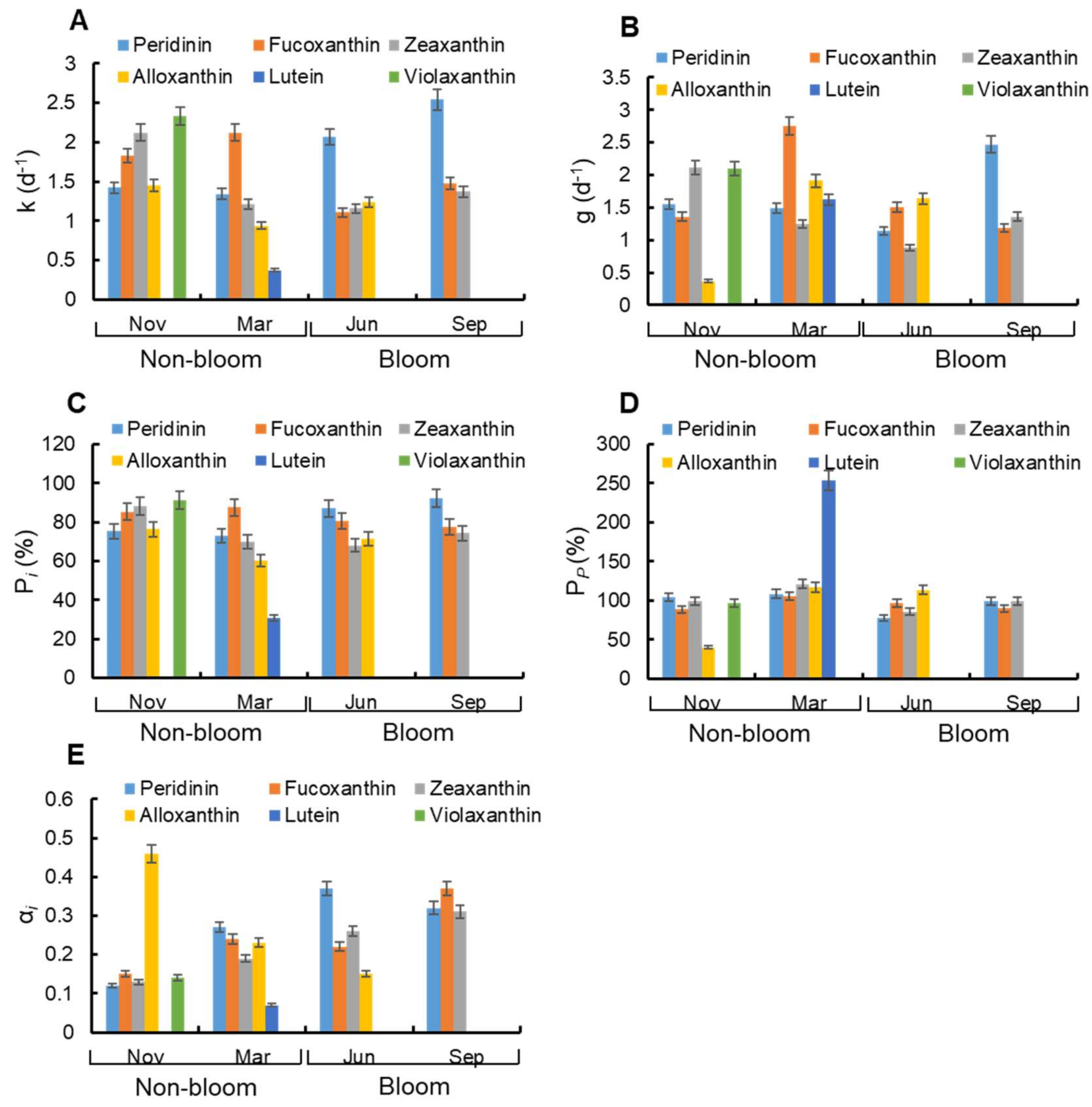
3.5. Microzooplankton Grazing on Pigment Specific Phytoplankton
4. Discussion
4.1. Changes in Phytoplankton Communities between Bloom and Non-Bloom Period
4.2. Changes in Zooplankton Communities between Bloom and Non-Bloom Period
4.3. Changes in Bacteria Communities between Bloom and Non-Bloom Period
4.4. Microzooplankton Grazing on Different Phytoplankton Groups between Bloom and Non-Bloom Period
4.5. Ecological Restoration of Eutrophic Lakes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Azam, F.; Fenchel, T.; Field, J.G.; Gray, J.S. The ecological role of water-column microbes in the sea. Mar. Ecol. Prog. Ser. 1983, 10, 257–263. [Google Scholar] [CrossRef]
- Garrison, D.L.; Gowing, M.M.; Hughes, M.P.; Campbell, L.; Caron, D.A.; Dennett, M.R.; Shalapyonok, A.; Olson, R.J.; Landry, M.R.; Brown, S.L.; et al. Microbial food web structure in the Arabian Sea: A US JGOFS study. Deep-Sea Res. Pt. II 2000, 47, 1387–1422. [Google Scholar] [CrossRef]
- Riemann, B.; Christoffersen, K. Microbial trophodynamics in temperate lakes. Mar. Microb. Food Webs 1993, 7, 69–100. [Google Scholar]
- Fenchel, T. The microbial loop-25 years later. J. Exp. Mar. Biol. Ecol. 2008, 366, 99–103. [Google Scholar] [CrossRef]
- Azam, F.; Malfatti, F. Microbial structuring of marine ecosystems. Nat. Rev. Microbiol. 2007, 5, 782–791. [Google Scholar] [CrossRef]
- Leakey, R.J.G.; Burkill, P.H.; Sleigh, M.A. Planktonic ciliates in Southampton Water: Abundance, biomass, production, and role in pelagic carbon flow. Mar. Biol. 1992, 114, 67–83. [Google Scholar]
- Miki, T.; Jacquet, S. Complex interactions in the microbial world: Underexplored key links between viruses, bacteria and protozoan grazers in aquatic environments. Aquat. Microb. Ecol. 2008, 51, 195–208. [Google Scholar] [CrossRef]
- Dussart, B.H. Les différentes catégories de plancton. Hydrobiologia 1965, 26, 72–74. [Google Scholar] [CrossRef]
- Calbet, A.; Saiz, E. The ciliate-copepod link in marine ecosystems. Aquat. Microb. Ecol. 2005, 38, 157–167. [Google Scholar] [CrossRef]
- Calbet, A.; Landry, M.R. Phytoplankton growth, microzooplankton grazing, and carbon cycling in marine systems. Limnol. Oceanog. 2004, 49, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Kainz, M.; Arts, M.T.; Mazumder, A. Essential fatty acids in the planktonic food web and their ecological role for higher trophic levels. Limnol. Oceanog. 2004, 49, 1784–1793. [Google Scholar] [CrossRef]
- Schmoker, C.; Hernández-León, S.; Calbet, A. Microzooplankton grazing in the oceans: Impacts, data variability, gaps of knowledge, and future directions. J. Plankton Res. 2013, 35, 691–706. [Google Scholar] [CrossRef]
- Liu, X.J.; Deng, C.H.; Wong, C.K. Microzooplankton selective feeding in subtropical coastal waters as revealed by HPLC pigment analysis. J. Sea Res. 2014, 90, 83–94. [Google Scholar] [CrossRef]
- Irigoien, X.; Flynn, K.; Harris, R.P. Phytoplankton blooms: A ‘loophole’ in microzooplankton grazing impact. J. Plankton Res. 2005, 27, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Sherr, E.B.; Sherr, B.F. Heterotrophic dinoilagellates: A significant component of microzooplankton biomass and major grazers of diatoms in the sea. Mar. Ecol. Prog. Ser. 2007, 352, 187–197. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Chen, X.; Wang, K.; Chen, J.; Zheng, B.; Jiang, X. Comparison among the microbial communities in the lake, lake wetland, and estuary sediments of a plain river network. Microbiol. Open 2019, 8, 00644. [Google Scholar] [CrossRef]
- Tandon, K.; Yang, S.H.; Wan, M.T.; Chia-Chin, Y.; Bayanmunkh, B.; Chiu-Yu, C.; Jeng-Wei, T.; Wen-Cheng, L.; Sen-Lin, T. Bacterial Community in Water and Air of Two Sub-Alpine Lakes in Taiwan. Microbes. Environ. 2018, 33, 120–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tranvik, L.J. Bacterioplankton Growth on Fractions of Dissolved Organic-Carbon of Different Molecular-Weights from Humic and Clear Waters. Appl. Environ. Microbiol. 1990, 56, 1672–1677. [Google Scholar] [CrossRef] [Green Version]
- Keshri, J.; Pradeep, R.A.S.; Sime-Ngando, T. Distinctive patterns in the taxonomical resolution of bacterioplankton in the sediment and pore waters of contrasted freshwater lakes. Microb. Ecol. 2017, 75, 662–673. [Google Scholar] [CrossRef]
- Wu, L.; Yu, Y.; Zhang, T.; Feng, W.; Zhang, X.; Li, W.; Wu, L.; Yu, Y.; Zhang, T.; Feng, W. PCR-DGGE Fingerprinting Analysis of Plankton Communities and Its Relationship to Lake Trophic Status. Int. Rev. Hydrobiol. 2010, 94, 528–541. [Google Scholar] [CrossRef]
- Souffreau, C.; van der Gucht, K.; van Gremberghe, I.; Kosten, S.; Lacerot, G.; Lobão, L.M.; de Moraes Huszar, V.L.; Roland, F.; Jeppesen, E.; Vyverman, W.; et al. Environmental rather than spatial factors structure bacterioplankton communities in shallow lakes along a >6000 km latitudinal gradient in South America. Environ. Microbiol. 2015, 17, 2336–2351. [Google Scholar] [CrossRef]
- Kong, Z.Y.; Kou, W.B.; Ma, Y.T.; Yu, H.T.; Ge, G.; Wu, L. Seasonal dynamics of the bacterioplankton community in a large, shallow, highly dynamic freshwater lake. Can. J. Microbiol. 2018, 64, 786–797. [Google Scholar] [CrossRef] [PubMed]
- Sadro, S.; Nelson, C.E.; Melack, J.M. Linking diel patterns in community respiration to bacterioplankton in an oligotrophic high-elevation lake. Limnol. Oceanogr. 2001, 56, 540–550. [Google Scholar] [CrossRef]
- Schweitzer-Natan, O.; Ofek-Lalzar, M.; Sher, D.; Sukenik, A. Particle-Associated Microbial Community in a Subtropical Lake during Thermal Mixing and Phytoplankton Succession. Front. Microbiol. 2019, 10, 2142. [Google Scholar] [CrossRef]
- Paver, S.F.; Hayek, K.R.; Gano, K.A.; Fagen, J.R.; Brown, C.T.; Davis-Richardson, A.; Crabb, D.B.; Rosario-Passapera, R.; Giongo, A.; Triplett, E.; et al. Interactions between specific phytoplankton and bacteria affect lake bacterial community succession. Environ. Microbiol. 2013, 15, 2489–2504. [Google Scholar] [CrossRef]
- Nygaard, K.; Tobiesen, A. Bacterivory in Algae—A Survival Strategy during Nutrient Limitation. Limnol. Oceanogr. 1993, 38, 273–279. [Google Scholar] [CrossRef]
- Tijdens, M.; Waal, D.B.V.D.; Slovackova, H.; Hoogveld, H.L.; Gons, H.J. Estimates of bacterial and phytoplankton mortality caused by viral lysis and microzooplankton grazing in a shallow eutrophic lake. Freshwater Biol. 2008, 53, 1126–1141. [Google Scholar] [CrossRef]
- Suttle, C.A. Viruses in the sea. Nature 2005, 437, 356–361. [Google Scholar] [CrossRef]
- Jankowiak, J.G.; Gobler, C.J. The Composition and Function of Microbiomes Within Microcystis Colonies Are Significantly Different Than Native Bacterial Assemblages in Two North American Lakes. Front. Microbiol. 2020, 11, 1016. [Google Scholar] [CrossRef]
- Yang, L.; Liu, Y.; Cao, X.Y.; Zhou, Z.; Wang, S.; Xiao, J.; Song, C.; Zhou, Y. Community composition specificity and potential role of phosphorus solubilizing bacteria attached on the different bloom-forming cyanobacteria. Microbiol. Res. 2017, 12, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Cai, Y.; Chen, Z.; Zhou, Y.; Li, P.; Kong, F. Diversity and abundance of aerobic anoxygenic phototrophic bacteria in two cyanobacterial bloom-forming lakes in China. Ann. Limnol. Int. J. Lim. 2010, 46, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Lu, D.; Liu, P.; Fan, T.; Peng, H.; Zhang, Z. The investigation of “water bloom” in the downstream of the Han River. Res. Environ. Sci. 2000, 13, 29–31. [Google Scholar]
- Zheng, J.; Zhong, C.; Deng, C. Discussion on definition of algal bloom. Water Resour. Prot. 2006, 22, 45–47. [Google Scholar]
- Hu, H.J.; Wei, Y.X. The freshwater algae of China–Systematics, Taxonomy and Ecology; Sciencep: Beijing, China, 2006. [Google Scholar]
- Kofoid, C.A.; Campbell, A.S. Aconspectus of the Marine and Fresh Water Ciliate Belonging to the Suborder Tintinnoniea, with Descriptions of New Species Principally from the Agssiz Expedition to the Eastern Tropical Pacific; University of California Press: Berkeley, CA, USA, 1929. [Google Scholar]
- Kofoid, C.A.; Campbell, A.S. The Ciliate: The Tintinnoinea; Museum of Comparative Zoology at Harvard College: Cambridge, MA, USA, 1939. [Google Scholar]
- Lee, J.J.; Hunter, S.H.; Bovee, E.C. An Illustrated Guide to the Protozoa; Society of Protozoologists Publishers: Guelph, ON, Canada, 1985; pp. 393–575. [Google Scholar]
- Feng, C.; Jia, J.; Wang, C.; Han, M.; Dong, C.; Huo, B.; Li, D.; Liu, X. Phytoplankton and Bacterial Community Structure in Two Chinese Lakes of Different Trophic Status. Microorganisms 2019, 7, 621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landry, M.R. Estimating rates of growth and grazing of phytoplankton by dilution method. In Handbook of Methods in Aquatic Microbial Ecology; Lewis Publishers: Ann Arbor, MI, USA, 1993; pp. 715–722. [Google Scholar]
- Burkill, P.H. Microzooplankton grazing and selectivity of phytoplankton in coastal waters. Mar. Biol. 1987, 93, 581–590. [Google Scholar] [CrossRef]
- Wong, C.K.; Wong, C.K. HPLC pigment analysis of marine phytoplankton during a red tide occurrence in Tolo Harbour, Hong Kong. Chemosphere 2003, 52, 1633–1640. [Google Scholar] [CrossRef]
- Verity, P.G.; Stoecker, D.K.; Sieracki, M.E.; Nelson, J.R. Grazing, growth and mortality of microzooplankton during the 1989 North Atlantic spring bloom at 47° N, 18° W. Deep Sea Res. I 1993, 40, 1793–1814. [Google Scholar] [CrossRef]
- Chesson, J. The estimation and analysis of preference and its relationship to foraging models. Ecology 1983, 64, 1297–1304. [Google Scholar] [CrossRef]
- Tang, C.Y.; Sun, B.; Yu, K.F.; Shi, J.T.; Liu, M.M.; Jiang, T.; Huo, Y.Z.; He, P.M. Environmental triggers of a Microcystis (Cyanophyceae) bloom in an artificial lagoon of Hangzhou Bay, China. Mar. Pollut. Bull. 2018, 135, 776–782. [Google Scholar] [CrossRef]
- Jan, L.; Perr, S. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University: London, UK, 2013. [Google Scholar]
- Jiang, Y.J.; He, W.; Liu, W.X.; Qin, N.; Ouyang, H.L.; Wang, Q.M.; Kong, X.Z.; He, Q.S.; Yang, C.; Yang, B.; et al. The seasonal and spatial variations of phytoplankton community and their correlation with environmental factors in a large eutrophic Chinese lake (Lake Chaohu). Ecol. Indic. 2014, 40, 58–67. [Google Scholar] [CrossRef]
- Harke, M.J.; Steffen, M.M.; Gobler, C.J.; Otten, T.G.; Wilhelm, S.W.; Wood, S.A.; Paerl, H.W. A review of the global ecology, genomics, and biogeography of the toxic cyanobacterium, Microcystis spp. Harmful Algae 2016, 54, 4–20. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Zhang, Y.; Yang, Z.; Wei, L.; Yang, W.; Chen, C.; Kong, F. Spatial and seasonal shifts in bloom-forming cyanobacteria in Lake Chaohu: Patterns and driving factors. Phycol. Res. 2016, 64, 44–55. [Google Scholar] [CrossRef]
- Watanabe, M.F. Toxic water bloom of blue-green alga: Biological and chemical characterization. Jpn. J. Limnol. 1993, 54, 225–243. [Google Scholar] [CrossRef]
- Berry, M.A.; Davis, T.W.; Cory, R.M.; Duhaime, M.B.; Johengen, T.H.; Kling, G.W.; Marino, J.A.; Den Uyl, P.A.; Gossiaux, D.; Dick, G.J.; et al. Cyanobacterial harmful algal blooms are a biological disturbance to Western Lake Erie bacterial communities. Environ. Microbiol. 2017, 19, 1149–1162. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, W.W. Health effects of toxin-producing cyanobacteria: “The CyanoHABs”. Hum. Ecol. Risk Assess 2001, 7, 1393–1407. [Google Scholar] [CrossRef]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Sarma, S.; Nandini, S. Life history strategies of cladocerans: Comparisons of tropical and temperature taxa. Hydrobiologia 2005, 542, 315–334. [Google Scholar] [CrossRef]
- Yang, Z.; Li, C.; Zhang, S.; Sun, B. Temporal and spatial distribution of chlorophyll-a concentration and the relationships with TN, TP concentrations in Lake Ulansuhai, Inner Mongolia. J. Lake Sci. 2009, 37, 403–408. [Google Scholar]
- Jin, Y.; Yu, R.; Zhang, Z.; Zhang, Q.; Li, M.; Cao, Z.; Wu, L.; Hao, Y. Spatiotemporal variability of phytoplankton functional groups in a shallow eutrophic lake from cold, arid regions. Environ. Monit. Assess. 2020, 192, 371. [Google Scholar] [CrossRef]
- Shen, Y.; Huang, Y.; Hu, J.; Li, P.; Zhang, C.; Lin, L.; Xu, P.; Zhang, J.; Chen, X. The nitrogen reduction in eutrophic water column driven by Microcystis blooms. J. Hazard. Mater. 2019, 385, 121578. [Google Scholar] [CrossRef]
- Sługocki, Ł.; Czerniawski, R. Trophic state (TSISD) and mixing type significantly influence pelagic zooplankton biodiversity in temper-ate lakes (NW Poland). PeerJ 2018, 6, e5731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nandini, S.; Sarma, S.S.S. Population growth of some genera of cladocerans (Cladocera) in relation to algal food (Chlorella vulgaris) levels. Hydrobiologia 2003, 491, 211–219. [Google Scholar] [CrossRef]
- Nandini, S.; Picazo-Paez, E.A.; Sarma, S.S.S. The combined effects of heavy metals (copperand zinc), temperature and food (Chlorell vulgaris) level on the demographic characters of Moina macrocopa (Crustacea: Cladocera). J. Environ. Sci. Health Part A Toxic Hazard. Subst. Environ. Eng. 2007, 42, 1433–1442. [Google Scholar] [CrossRef]
- Kauler, P.; Enesco, H.E. The effect of temperature on life history parameters and cost of reproduction in the rotifer Brachionus calyciflorus. J. Freshw. Ecol. 2011, 26, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Xi, Y.L.; Ge, Y.L.; Chen, F.; Wen, X.L.; Dong, L.L. Life history characteristics of three strains of Brachionus calyciflorus (Rotifera) at different temperatures. J. Freshw. Ecol. 2005, 20, 707–713. [Google Scholar] [CrossRef]
- Wu, J.; Yan, B.; Feng, Z.; Li, Y.; Shen, X. Zooplankton ecology near the tianwan nuclear power station. Acta Ecol. Sinica. 2011, 31, 6902–6911. [Google Scholar]
- Wang, Y.; He, Z.H. Effect of temperature and salinity on intrinsic increasing rate of Moina mongolica Daddy (Cladocera: Moinidae) population. J. Appl. Ecol. 2011, 12, 91–94. [Google Scholar]
- Schalau, K.; Rinke, K.; Straile, D.; Peeters, F. Temperature is the key factor explaining interannual variability of Daphnia development in spring: A modeling study. Oecologia 2008, 157, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Araiza, M.A.; Sarma, S.S.S.; Nandini, S. Combined effects of food concentration and temperature on competition among four species of Brachionus (Rotifera). Hydrobiologia 2005, 546, 519–534. [Google Scholar] [CrossRef]
- Espinosa-Rodríguez, C.A.; Sarma, S.S.S.; Nandini, S. Interactions between the rotifer Euchlanis dilatata and the cladocerans Alona glabra and Macrothrix triserialis in relation to diet type. Limnologica 2012, 42, 50–55. [Google Scholar] [CrossRef]
- Feniova, I.Y.; Budaev, S.V.; Dgebuadze, Y.Y. Simulation of cladoceran survival strategy under conditions of food depletion. Russ. J. Ecol. 2006, 37, 28–34. [Google Scholar] [CrossRef]
- Lampert, W.; Sommer, U. Limnoecology: The Ecology of Lakes and Streams; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Dettmers, J.M.; Wahl, D.H. Evidence for zooplankton compensation and reduced fish growth in response to increased juvenile fish density. Hydrobiologia 1999, 400, 115–121. [Google Scholar] [CrossRef]
- Romare, P.; Bergman, E.; Hansson, L.A. The impact of larval and juvenile fish on zooplankton and algal dynamics. Limnol. Oceanogr. 1999, 44, 1655–1666. [Google Scholar] [CrossRef]
- Gilbert, J.J. Suppression of rotifer populations by Daphnia: A review of the evidence, the mechanisms, and the effects on zooplankton community structure. Limnol. Oceanogr. 1988, 33, 1286–1303. [Google Scholar] [CrossRef]
- Hurtado-Bocanegra, M.D.; Nandini, S.; Sarma, S.S.S. Combined effects of food level and inoculation density on competition between Brachionus patulus (Rotifera) and the cladocerans Ceriodaphnia dubia and Moina macrocopa. Hydrobiologia 2002, 468, 13–22. [Google Scholar] [CrossRef]
- Xi, Y.L.; Hagiwara, A. Competition between the rotifer Brachionus calyciflorus and the Cladoceran Moina macrocopa in relation to algal food concentration and initial rotifer population density. J. Freshw. Ecol. 2007, 22, 421–427. [Google Scholar] [CrossRef]
- Benincà, E.; Dakos, V.; van Nes, E.H.; Huisman, J.; Scheffer, M. Resonance of plankton communities with temperature fluctuations. Am. Nat. 2011, 178, E87–E95. [Google Scholar] [CrossRef] [PubMed]
- Sterner, R.W.; Hessen, D.O. Algal nutrient limitation and the nutrition of aquatic herbivores. Annu. Rev. Ecol. S 1994, 25, 1–29. [Google Scholar] [CrossRef]
- Elser, J.J.; Hayakawa, K.; Urabe, J. Nutrient limitation reduces food quality for zooplankton: Daphnia response to seston phosphorus enrichment. Ecology 2001, 82, 898–903. [Google Scholar] [CrossRef]
- Ferrao-Filho, A.; Arcifa, M.; Fileto, C. Resource limitation and food quality for cladocerans in a tropical Brazilian lake. Hydrobiologia 2003, 491, 201–210. [Google Scholar] [CrossRef]
- Persson, J.; Vrede, T.; Holmgren, S. Responses in zooplankton populations to food quality and quantity changes after whole lake nutrient enrichment of an oligotrophic subalpine reservoir. Aquat. Sci. 2008, 70, 142–155. [Google Scholar] [CrossRef]
- Langeland, A. Biomanipulation development in Norway. Hydrobiologia 1990, 200, 535–540. [Google Scholar] [CrossRef]
- George, D.G.; Edwards, R.W. Population dynamicsand produc-tion of daphnia hyaline in a eutrophic reservoir. Freshw. Biol. 1974, 4, 445–465. [Google Scholar] [CrossRef]
- Jarvis, A.C. Zooplankton community grazing in a hyper-trophic lake (Harbeespoort Dam, South Africa). Plankton Res. 1986, 8, 1065–1078. [Google Scholar] [CrossRef]
- Moore, M.V.; Folt, C.L.; Stemberger, R.S. Consequences of elevated temperatures for zooplankton assemblages in temperate lakes. Archiv. Hydrobiol. 1996, 135, 289–319. [Google Scholar]
- Liang, Y.; Ouyang, K.; Chen, X.; Su, Y.; Yang, J. Life strategy and grazing intensity responses of Brachionus calyciflorus fed on different concentrations of microcystin-producing and microcystin-free Microcystis aeruginosa. Sci. Rep. 2017, 7, 43127. [Google Scholar] [CrossRef] [Green Version]
- Fulton, R.S., III; Paerl, H.W. Effects of colonial morphologyon zooplankton utilization of algal resources during blue-green algal (Microcystis aeruginosa) blooms. Limnol. Oceanog. 1987, 32, 634–644. [Google Scholar] [CrossRef]
- Anders, F.A.; Lasse, R.; Stefan, B. Pyrosequencing reveals contrasting seasonal dynamics of taxa within Baltic Sea bacterioplankton communities. ISME J. 2010, 4, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Riemann, L.; Steward, G.F.; Azam, F. Dynamics of bacterial community composition and activity during a mesocosm diatom bloom. Appl. Environ. Microbiol. 2000, 66, 578–587. [Google Scholar] [CrossRef] [Green Version]
- Niu, Y.; Shen, H.; Chen, J.; Xie, P.; Yang, X.; Tao, M.; Ma, Z.; Qi, M. Phytoplankton community succession shaping bacterioplankton community composition in Lake Taihu, China. Water Res. 2011, 45, 4169–4182. [Google Scholar] [CrossRef] [PubMed]
- Furman, J.A.; Azam, F. Bacterioplankton secondary production estimates for coastal waters of Birth Columbia, Antarctica and Southern California. Appl. Environ. Microbiol. 1989, 39, 1085–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ducklow, H.W. Factors regulating bottom-up control of bacteria biomass in open ocean plankton communities. Areh. Hydrobiol. Beih. 1992, 37, 207–217. [Google Scholar]
- Jennifer, C.; James, A.A.E.; Ellen, T.M. Utilization and turnover of labile dissolved organic matter by bacterial heterotrophs in eastern North Pacific surface waters. Mar. Ecol. Prog. Ser. 1996, 139, 267–279. [Google Scholar]
- Larsson, U.; Hagstrom, A. Phytopldnkton exuddte release as an energy source for the growth of peldgic bacteria. Mar. Biol. 1979, 52, 199–206. [Google Scholar] [CrossRef]
- Morris, D.P.; Lewis, W.M.J. Nutrient limitation of bacterioplankton growth in Lake Dillo, Colorado. Limnol. Oceanog. 1992, 37, 1179–1192. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Miller, T.D.; Priscu, J.C. Bacterioplankton nutrient deficiency in a eutrophic lake. Arch. Hydrobiol. 1992, 125, 423–439. [Google Scholar]
- Toolan, T.; Wehr, I.D.; Findlay, S. Inorganic phosphorus stimulation of bacterioplankton production in a meso-eutrophic lake. Appl. Environ. Microbiol. 1991, 57, 2074–2078. [Google Scholar] [CrossRef] [Green Version]
- Muylaert, K.; van der Gucht, K.; Vloemans, N.; Meester, L.D.; Gillis, M.; Vyverman, W. Relationship between bacterial community composition and bottom-up versus top-down variables in four eutrophic shallow lakes. Appl. Environ. Microbiol. 2000, 66, 4740–4750. [Google Scholar] [CrossRef] [Green Version]
- Caron, D.A.; Lim, E.L.; Sanders, R.W.; Dennett, M.R.; Berninger, U.G. Responses of bacterioplankton and phytoplankton to organic carbon and inorganic nutrient additions in contrasting oceanic ecosystems. Aquat. Microb. Ecol. 2002, 22, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Ji, B.; Qin, H.; Guo, S.; Chen, W.; Zhang, X.; Liang, J. Bacterial communities of four adjacent fresh lakes at different trophic status. Ecotoxicol. Environ. Saf. 2018, 157, 388–394. [Google Scholar] [CrossRef]
- Paerl, H.W. Transfer of N2 and CO2 fixation products from Anabaena oscillarioides to associated bacteria during inorganic carbon sufficiency and deficiency. J. Phycol. 1984, 20, 600–608. [Google Scholar] [CrossRef]
- Rashidan, K.K.; Bird, D.F. Role of predatory bacteria in the termination of a cyanobacterial bloom. Microb. Ecol. 2001, 41, 97–l05. [Google Scholar] [CrossRef]
- Pinhassi, J.; Sala, M.M.; Havskum, H.; Peters, F.; Guadayol, O.; Malits, A.; Marrase, C. Changes in bacterioplankton composition under different phytoplankton regimens. Appl. Environ. Microbiol. 2004, 70, 6753–6766. [Google Scholar] [CrossRef] [Green Version]
- Bezek, K.D.; Nipič, D.; Torkar, K.G.; Oder, M.; Dražić, G.; Abram, A.; Žibert, J.; Raspor, P.; Bohinc, K. Biofouling of stainless steel surfaces by four common pathogens: The effects of glucose concentration, temperature and surface roughness. Biofouling 2019, 35, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Minami, C.; Liu, H.; Saino, T. Temporal and spatial patterns of chemotaxonomic algal pigments in the subarctic Pacific and the Bering Sea during summer 1999. Deep-Sea Res. II 2002, 49, 5685–5704. [Google Scholar] [CrossRef]
- Ger, K.A.; Hansson, L.-A.; Lürling, M. Understanding cyanobacteria-zooplankton interactions in a more eutrophic world. Freshw. Biol. 2014, 59, 1783–1798. [Google Scholar] [CrossRef] [Green Version]
- Chotiyaputta, C.; Hirayama, K. Food selectivity of the rotifer Brachionus plicatilis feeding on phytoplankton. Mar. Biol. 1978, 45, 105–111. [Google Scholar] [CrossRef]
- Löder, M.G.J.; Meunier, C.; Wiltshire, K.H.; Boersma, M.; Aberle, N. The role of ciliates, heterotrophic dinoflagellates and copepods in structuring spring plankton communities at Helgoland Roads, North Sea. Mar. Biol. 2011, 158, 1551–1580. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, J.; Lamarra, V.; Lynch, M. Biomanipulation: An ecosystem approach to lake restoration. In Proceedings of a Symposium on Water Quality Management through Biological Control; Brezomik, P.L., Fox, J.L., Eds.; University of Florid: Gainesville, FL, USA, 1975; pp. 85–89. [Google Scholar]
- Xie, P.; Liu, J. Practical Success of Biomanipulation using Filter-feeding Fish to Control Cyanobacteria Blooms. Sci. World 2001, 1, 337–356. [Google Scholar]
- Benndorf, J.; Kneshke, H.; Kossatz, K.; Penz, E. Manipulation of the pelagic food web by stocking with predacious fishes. Int. Rev. Gesamten Hydrobiol. 1984, 69, 407–428. [Google Scholar] [CrossRef]
- Jurajda, P.; Adámek, Z.; Janáč, M.; Špaček, J. Use of multiple fish-removal methods during biomanipulation of a drinking water reservoir—Evaluation of the first four years. Fish. Res. 2015, 173, 101–108. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Time | Sample ID | Assigned Reads | 97% Similarity | ||||
|---|---|---|---|---|---|---|---|
| OTUs | Shannon | Chao1 | Ace | Coverage | |||
| March | S1 | 38,094 | 584 | 4.7549 | 757.109 | 725.66 | 0.996614 |
| S2 | 32,198 | 678 | 4.7303 | 826.061 | 755.73 | 0.995341 | |
| S3 | 31,849 | 737 | 4.6504 | 673.0171 | 664.86 | 0.996326 | |
| May | S1 | 32,367 | 691 | 4.4499 | 867.581 | 858.64 | 0.994161 |
| S2 | 33,210 | 844 | 4.4581 | 970.068 | 910.42 | 0.993767 | |
| S3 | 32,352 | 939 | 4.5401 | 953.432 | 944.28 | 0.993447 | |
| November | S1 | 32,773 | 1455 | 6.0961 | 1507.468 | 1492.73 | 0.994845 |
| S2 | 29,682 | 1550 | 6.1652 | 1569.113 | 1529.79 | 0.995332 | |
| S3 | 36,060 | 1595 | 6.1025 | 1568.609 | 1535.84 | 0.995563 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, M.; Dong, C.; Ma, S.; Feng, C.; Lei, C.; Gu, Z.; Liu, X. Food Web Responses to a Cyanobacterial Bloom in a Freshwater Eutrophic Lake. Water 2021, 13, 1296. https://doi.org/10.3390/w13091296
Han M, Dong C, Ma S, Feng C, Lei C, Gu Z, Liu X. Food Web Responses to a Cyanobacterial Bloom in a Freshwater Eutrophic Lake. Water. 2021; 13(9):1296. https://doi.org/10.3390/w13091296
Chicago/Turabian StyleHan, Mengqi, Chenchen Dong, Siqi Ma, Cui Feng, Chengqiang Lei, Zemao Gu, and Xiangjiang Liu. 2021. "Food Web Responses to a Cyanobacterial Bloom in a Freshwater Eutrophic Lake" Water 13, no. 9: 1296. https://doi.org/10.3390/w13091296
APA StyleHan, M., Dong, C., Ma, S., Feng, C., Lei, C., Gu, Z., & Liu, X. (2021). Food Web Responses to a Cyanobacterial Bloom in a Freshwater Eutrophic Lake. Water, 13(9), 1296. https://doi.org/10.3390/w13091296