
Critical Transitions in Lake Ecosystem State May Be Driven by Coupled Feedback Mechanisms: A Case Study from Lake Erhai, China


Abstract
:1. Introduction
2. Materials and Methods
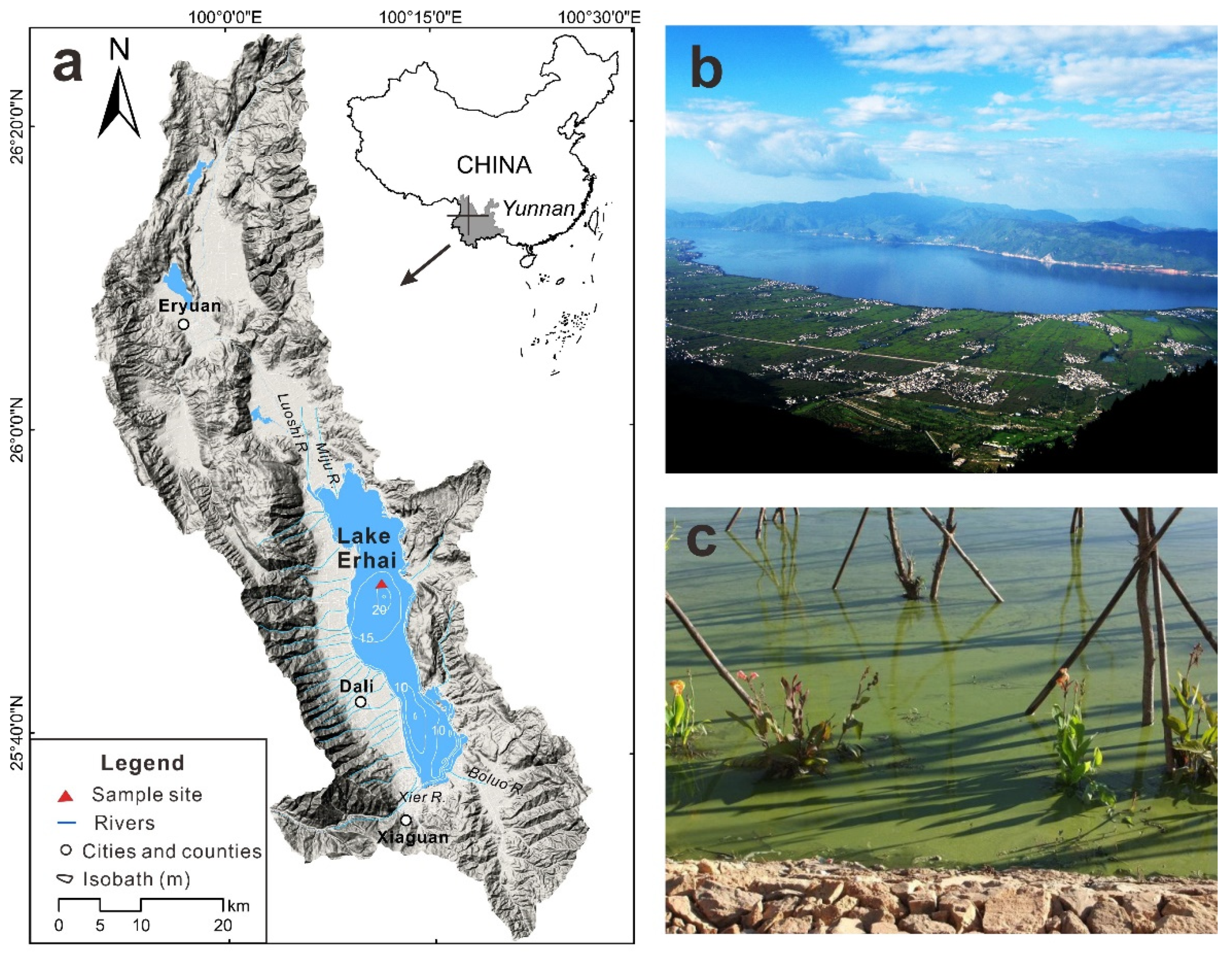
2.1. Study Site
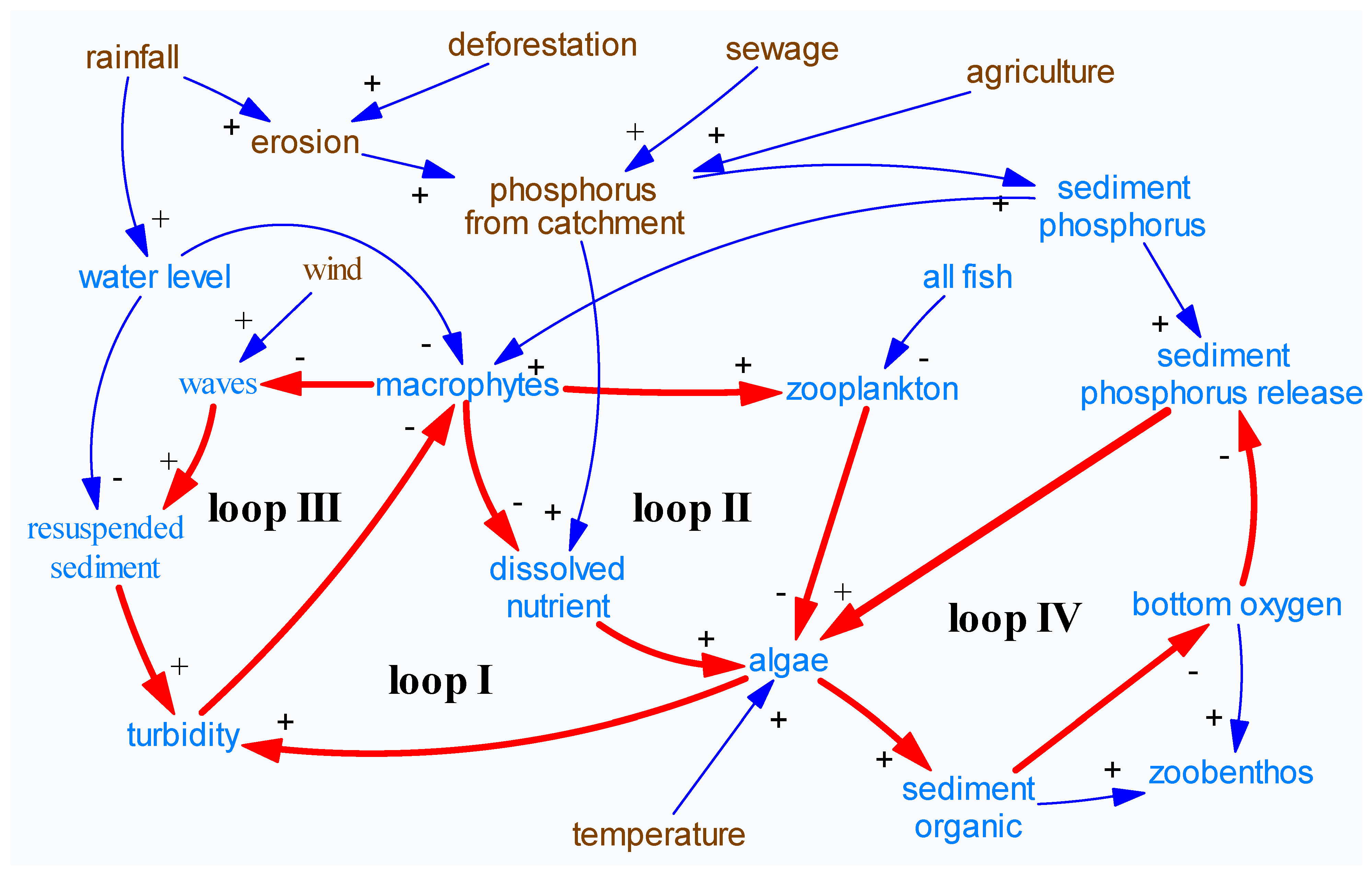
2.2. Causal Loop and Hypotheses
- Loop I Macrophyte-dissolved nutrient-algae-turbidity-macrophytes;
- Loop II Macrophyte-zooplankton-algae-turbidity-macrophytes;
- Loop III Macrophyte-waves-resuspended sediment-turbidity-macrophytes;
- Loop IV Algae-organic material-bottom water anoxia-nutrient release-algae.
2.3. Analyses
3. Results
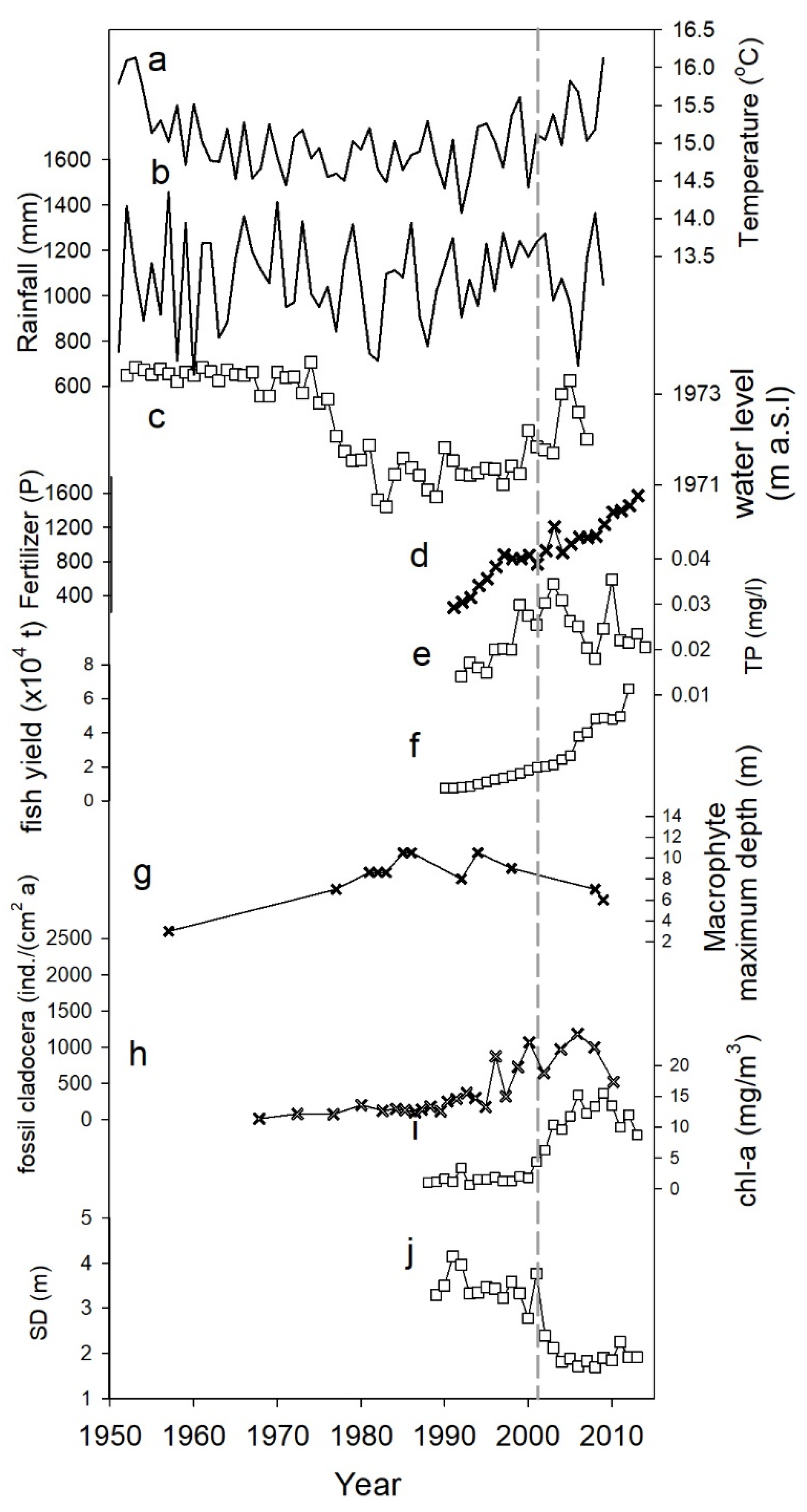
3.1. Long-Term Change (1950s–2010s)
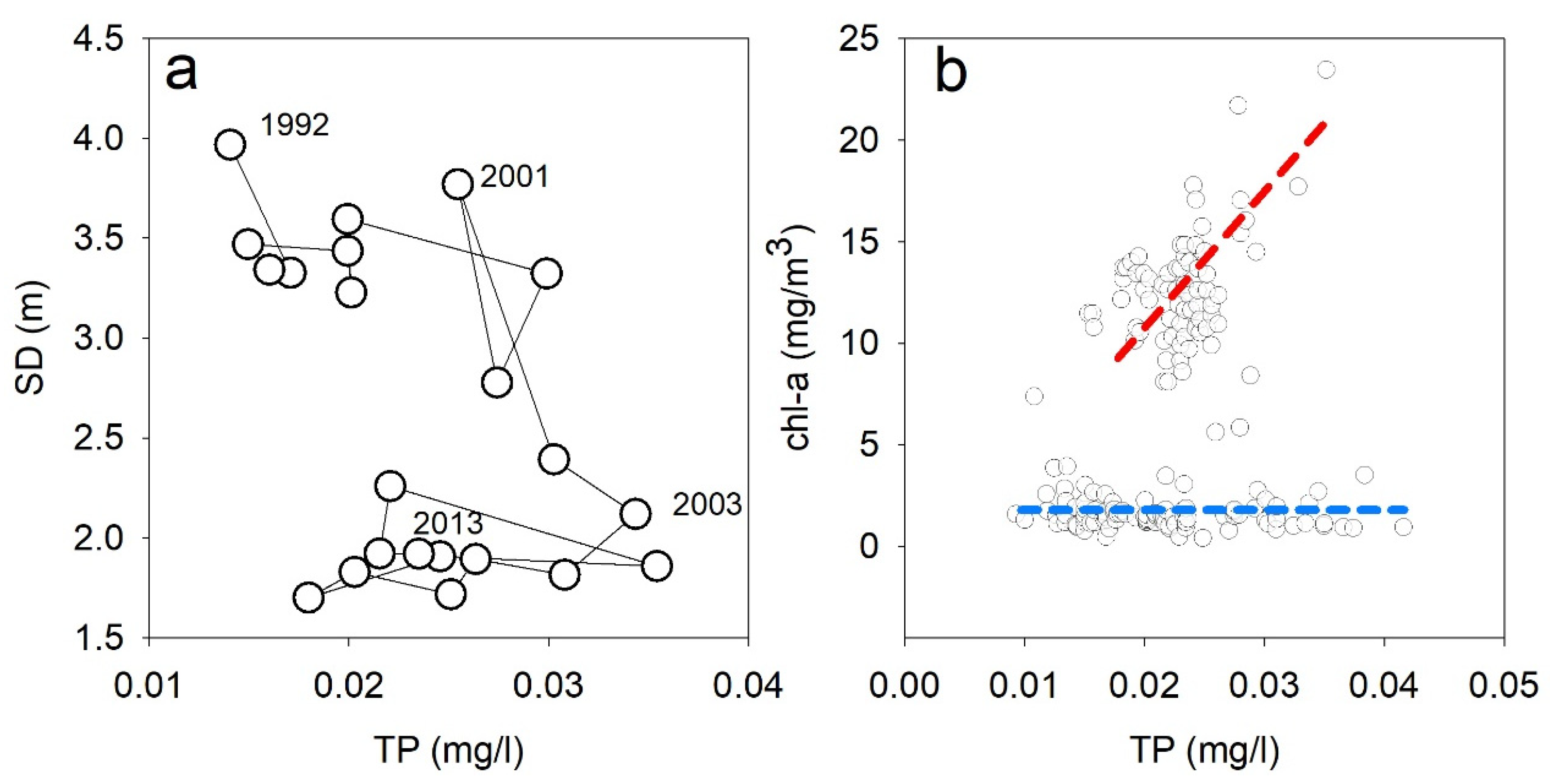
3.2. Critical Transition (2000–2004)
3.3. Testing the Hypothesized Loops—Long-Term Analysis (1950s–2010s)
3.4. Testing the Hypothesized Loops—Short Term Analysis (2000–2004)
4. Discussion
5. Conclusions
- Using a range of paleolimnological, monitored and documentary data sources from Erhai, China, we reconstructed the significant positive feedback loops prior to a state shift from clear to turbid water conditions in 2001 brought about by multiple stresses;
- We identified the growth and decline of two significant and positive feedback loops coupled through phytoplankton: a macrophyte-loop and a P-recycling loop. As the regulating macrophyte-loop became weaker during the process of lake eutrophication, the re-organizing P-recycling-loop became stronger and was ultimately responsible for transitioning the lake state across a tipping point;
- This insight suggests that management aimed at restoring the lake to the earlier clear, stable state needs to focus initially on weakening the P-recycling loop through controls on nutrient loading, water levels and fish farming;
- The presence of a critical transition preceded by changing feedback loops agrees with alternative stable state theory, but we suggest that the lake’s hysteretic path will be asymmetrical with an incremental, rather than abrupt, recovery phase.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scheffer, M. Critical Transitions in Nature and Society; Princeton University Press: Princeton, NJ, USA, 2009. [Google Scholar]
- Barnosky, A.D.; Hadly, E.A.; Bascompte, J.; Berlow, E.L.; Brown, J.H.; Fortelius, M.; Getz, W.M.; Harte, J.; Hastings, A.; Marquet, P.A.; et al. Approaching a state shift in Earth’s biosphere. Nature 2012, 486, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Scheffer, M.; Carpenter, S.R.; Foley, J.A.; Folke, C.; Walker, B. Catastrophic shifts in ecosystems. Nature 2001, 413, 591–596. [Google Scholar] [CrossRef]
- Scheffer, M.; Bascompte, J.; Brock, W.A.; Brovkin, V.; Carpenter, S.R.; Dakos, V.; Held, H.; van Nes, E.H.; Rietkerk, M.; Sugihara, G. Early-warning signals for critical transitions. Nature 2009, 461, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Holling, C.S. Resilience and stability of ecological systems. Annu. Rev. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Folke, C.; Carpenter, S.R.; Walker, B.; Scheffer, M.; Chapin, T.; Rockstrom, J. Resilience Thinking: Integrating Resilience, Adaptability and Transformability. Ecol. Soc. 2010, 15, 20. [Google Scholar] [CrossRef]
- May, R.M. Thresholds and breakpoints in ecosystems with a multiplicity of stable states. Nature 1977, 269, 471–477. [Google Scholar] [CrossRef]
- Dakos, V.; Scheffer, M.; van Nes, E.H.; Brovkin, V.; Petoukhov, V.; Held, H. Slowing down as an early warning signal for abrupt climate change. Proc. Natl. Acad. Sci. USA 2008, 105, 14308–14312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Dearing, J.A.; Langdon, P.G.; Zhang, E.; Yang, X.; Dakos, V.; Scheffer, M. Flickering gives early warning signals of a critical transition to a eutrophic lake state. Nature 2012, 492, 419–422. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Cole, J.J.; Pace, M.L.; Batt, R.; Brock, W.A.; Cline, T.; Coloso, J.; Hodgson, J.R.; Kitchell, J.F.; Seekell, D.A.; et al. Early warnings of regime shifts: A whole-ecosystem experiment. Science 2011, 332, 1079–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veraart, A.J.; Faassen, E.J.; Dakos, V.; van Nes, E.H.; Lürling, M.; Scheffer, M. Recovery rates reflect distance to a tipping point in a living system. Nature 2012, 481, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Vorselen, D.; Korolev, K.S.; Gore, J. Generic indicators for loss of resilience before a tipping point leading to population collapse. Science 2012, 336, 1175–1177. [Google Scholar] [CrossRef] [Green Version]
- Scheffer, M.; Carpenter, S.R.; Lenton, T.M.; Bascompte, J.; Brock, W.; Dakos, V.; van de Koppel, J.; van de Leemput, I.A.; Levin, S.A.; van Nes, E.H. Anticipatimg critical transitons. Science 2012, 338, 344–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, G.S.; Dearing, J.A. Modelling future safe and just operating spaces in regional social-ecological systems. Sci. Total Environ. 2018, 651, 2105–2117. [Google Scholar] [CrossRef]
- Zhang, K.; Dearing, J.A.; Dawson, T.P.; Dong, X.H.; Yang, X.D.; Zhang, W.G. Poverty alleviation strategies in eastern China lead to critical ecological dynamics. Sci. Total Environ. 2015, 506, 164–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Dearing, J.A.; Doncaster, C.P.; Yang, X.D.; Zhang, E.L.; Langdon, P.G.; Yang, H.; Dong, X.H.; Hu, Z.J.; Xu, M.; et al. Network parameters quantify loss of assemblage structure in human-impacted lake ecosystems. Glob. Chang. Biol. 2019, 25, 3871–3882. [Google Scholar] [CrossRef] [PubMed]
- Doncaster, C.P.; Chavez, V.A.; Viguier, C.; Wang, R.; Zhang, E.L.; Dong, X.H.; Dearing, J.A.; Langdon, P.G.; Dyke, J.G. Early warning of critical transitions in biodiversity from compositional disorder. Ecology 2016, 97, 3079–3090. [Google Scholar] [CrossRef] [PubMed]
- Suding, K.N.; Hobbs, R.J. Threshold models in restoration and conservation: A developing framework. Trends Ecol. Evol. 2009, 24, 271–279. [Google Scholar] [CrossRef]
- Zhao, Q.H.; Sun, J.H. The variation features of the surface mixed layer depth in Erhai Lake and Taihu Lake in spring and autumn and their mechanism analyses. Acta Phys. Sin. 2013, 62, 039203. [Google Scholar]
- Yan, C.; Jin, X.; Zhao, J.; Shen, B.; Li, N.; Huang, C.; Xiong, Z. Ecological Protection and Sustainable Utilization of Erhai Lake, Yunnan. Environ. Sci. 2005, 26, 38–43. [Google Scholar]
- Yang, S.; Ni, X. The status of agriculture Non-point pollution in watershed of Erhai lake. Agro-Environ. Dev. 1999, 2, 65–70. [Google Scholar]
- World Bank. China—Second Yunnan Urban Environmental Project (Vol. 5): Erhai Lake Basin Regional Environmental Assessment (English). Washingto, DC: World Bank. 2007. Available online: http://documents.worldbank.org/curated/en/407591468218982740/Erhai-Lake-Basin-regional-environmental-assessment (accessed on 31 December 2021).
- Correl, D.L. The role of phosphorus in the eutrophication of receiving waters: A review. J. Environ. Qual. 1998, 27, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Scheffer, M.; Hosper, S.H.; Meijer, M.L.; Moss, B.; Jeppesen, E. Alternative equilibria in shallow lakes. Trends Ecol. Evol. 1993, 8, 275–279. [Google Scholar] [CrossRef]
- Scheffer, M. Ecology of Shallow Lakes, 1st ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands; Boston, MA, USA, 1998. [Google Scholar]
- Chen, X.H. Study on the Recent 20-Year Eutrophication Process and Development of Nutrient Criteria/Standards of Lake Erhai at Its Initial Stage of Eutrophication. Ph.D. Thesis, East China Normal University, Shanghai, China, 2015. [Google Scholar]
- Wang, X.M. The Spatial-Temporal Analysis of the Water-Environment Effects of Industry Structure in Erhai Basin. Master’s Thesis, Central China Normal University, Wuhan, China, 2016. [Google Scholar]
- Wu, G.G.; Ni, L.Y.; Cao, T.; Xie, P.; Xu, J. Patterns and controls of dynamics of macrophytes and phytoplankton changes in Lake Erhai from 1977 to 2009. Acta Hydrobiol. Sin. 2013, 37, 912–918. [Google Scholar]
- Liu, G.; Liu, Z.; Gu, B.; Mmoak, J.M.; Zhang, Z. How important are trophic state, macrophyte and fish population effects on cladoceran community? A study in Lake Erhai. Hydrobiologia 2014, 736, 189–204. [Google Scholar] [CrossRef]
- Dearing, J.A. Sedimentary indicators of lake-level changes in the humid temperate zone: A critical review. J. Paleolimnol. 1997, 18, 1–14. [Google Scholar] [CrossRef]
- Hupfer, M.; Lewandowski, J. Oxygen Controls the Phosphorus Release from Lake Sediments—A Long-Lasting Paradigm in Limnology. Int. Rev. Hydrobiol. 2008, 93, 415–432. [Google Scholar] [CrossRef]
- Du, B.; Li, Y. Danger Risk to Fish Diversity in Erhai Lake and Proposals to Dispel it. Res. Environ. Sci. 2001, 14, 42–45. [Google Scholar]
- van der Heide, T.; van Nes, E.H.; Geerling, G.W.; Smolder, A.J.P.; Bouma, T.J.; van Katwijk, M.M. Positive feedbacks in seagrass ecosystems: Implications for success in conservation and restoration. Ecosystems 2007, 10, 1311–1322. [Google Scholar] [CrossRef]
- van Nes, E.H.; Hirota, M.; Holmgren, M.; Scheffer, M. Tipping points in tropical tree cover: Linking theory to data. Glob. Chang. Biol. 2014, 20, 1016–1021. [Google Scholar] [CrossRef]
- Kéfi, S.; Holmgren, M.; Scheffer, M. When can positive interactions cause alternative stable states in ecosystems? Funct. Ecol. 2016, 30, 88–97. [Google Scholar] [CrossRef]
- van de Leemput, I.A.; Hughes, T.P.; van Nes, E.H.; Scheffer, M. Multiple feedbacks and the prevalence of alternate stable states on coral reefs. Coral Reefs 2016, 35, 857–865. [Google Scholar] [CrossRef]
- Yan, X.; Xu, X.; Wang, M.; Wang, G.; Wu, S.; Li, Z.; Sun, H.; Shi, A.; Yang, Y. Climate warming and cyanobacteria blooms: Loos at their relationships from a new perspective. Water Res. 2017, 125, 449–457. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System Component | Indicator | Date Range | Resolution of Sampling | Data Source |
|---|---|---|---|---|
| turbidity | Secchi disk depth | 1989–2013 | monthly | Chen, 2015 [26] |
| dissolved nutrient | total P in water column | 1992–2014 | monthly | Wang, 2016 [27] |
| bottom oxygen | fossil chironomid head capsules | 1822–2007 | sedimentation rate dependent | Wang et al., 2012 [9] |
| P from catchment | P fertilizer use | 1990–2012 | annual | Chen, 2015 [26] |
| sediment organic | total organic carbon in sediment | 1900–2010 | sedimentation rate dependent | Wang et al., 2012 [9] |
| sediment P | P in sediment | 1900–2010 | sedimentation rate dependent | Wang et al., 2012 [9] |
| sediment P release | the ratio of sediment P and Fe | 1900–2010 | sedimentation rate dependent | Wang et al., 2012 [9] |
| macrophytes | maximum macrophyte depth | 1957–2009 | single survey | Wu et al., 2013 [28] |
| algae | chlorophyll-a concentration | 1988–2013 | monthly | Chen, 2015 [26] |
| zooplankton | fossil Cladocera | 1956–2012 | sedimentation rate dependent | Liu et al., 2014 [29] |
| fish | fish yield | 1990–2012 | annual | Chen, 2015 [26] |
| water level | monitoring lowest water level | 1952–2011 | annual | Wang et al., 2012 [9] |
| temperature | monitoring temperature in Dali station | 1951–2010 | daily | National Meteorological Information Center (http://data.cma.cn, accessed on 31 December 2021) |
| rainfall | monitoring rainfall in Dali station | 1951–2009 | daily | National Meteorological Information Center (http://data.cma.cn, accessed on 31 December 2021) |
| wind | monitoring wind in Dali station | 1951–2010 | daily | National Meteorological Information Center (http://data.cma.cn, accessed on 31 December 2021) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; Dearing, J.A.; Langdon, P.G. Critical Transitions in Lake Ecosystem State May Be Driven by Coupled Feedback Mechanisms: A Case Study from Lake Erhai, China. Water 2022, 14, 85. https://doi.org/10.3390/w14010085
Wang R, Dearing JA, Langdon PG. Critical Transitions in Lake Ecosystem State May Be Driven by Coupled Feedback Mechanisms: A Case Study from Lake Erhai, China. Water. 2022; 14(1):85. https://doi.org/10.3390/w14010085
Chicago/Turabian StyleWang, Rong, John A. Dearing, and Peter G. Langdon. 2022. "Critical Transitions in Lake Ecosystem State May Be Driven by Coupled Feedback Mechanisms: A Case Study from Lake Erhai, China" Water 14, no. 1: 85. https://doi.org/10.3390/w14010085
APA StyleWang, R., Dearing, J. A., & Langdon, P. G. (2022). Critical Transitions in Lake Ecosystem State May Be Driven by Coupled Feedback Mechanisms: A Case Study from Lake Erhai, China. Water, 14(1), 85. https://doi.org/10.3390/w14010085