
Sediment as a Refuge Spot for Planktonic Crustaceans



 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
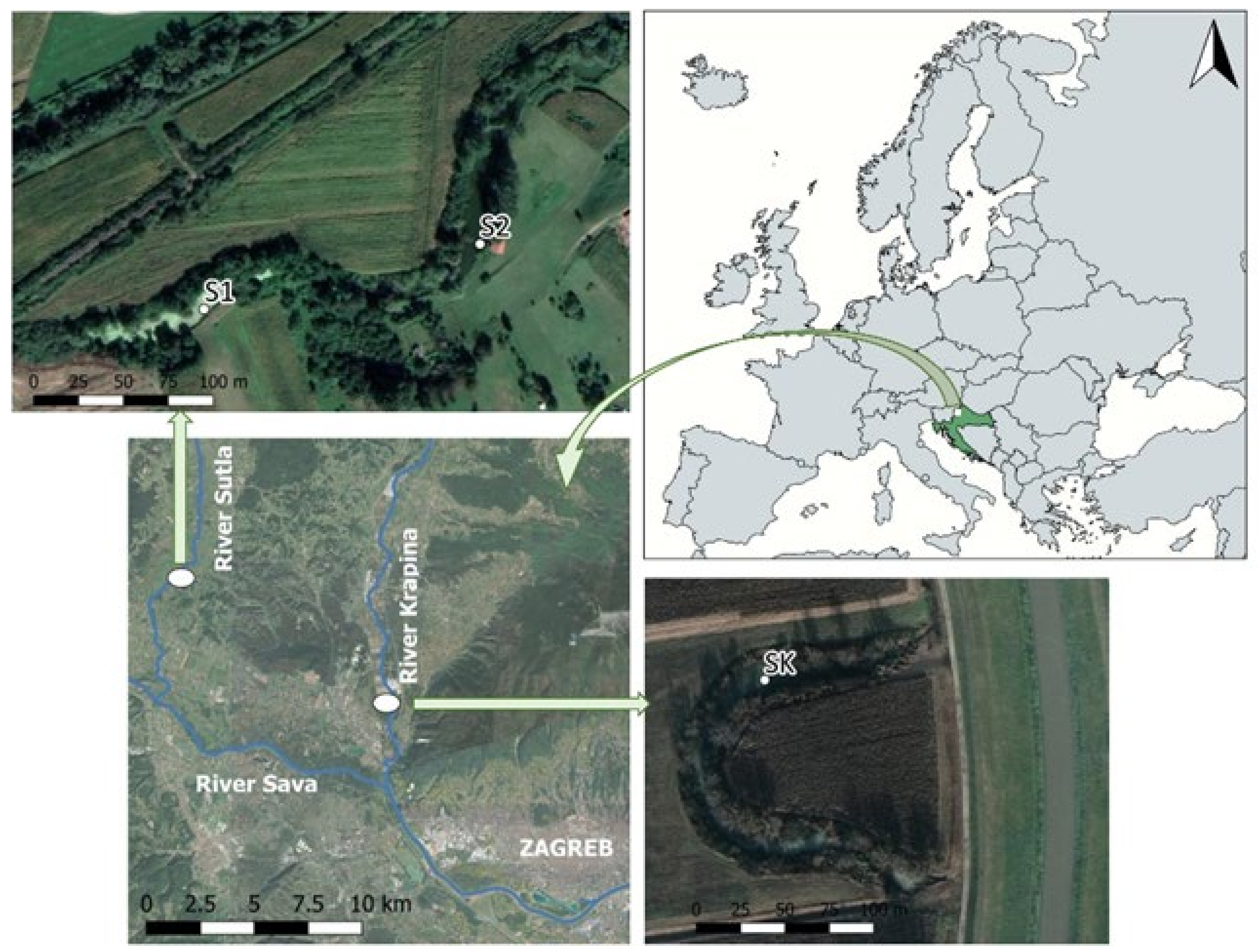
2.1. Study Sites
2.2. Collection and Analysis of Biocoenotic Components
2.3. Determination of Limnological Factors and Sediment Analysis
2.4. Statistical Analysis
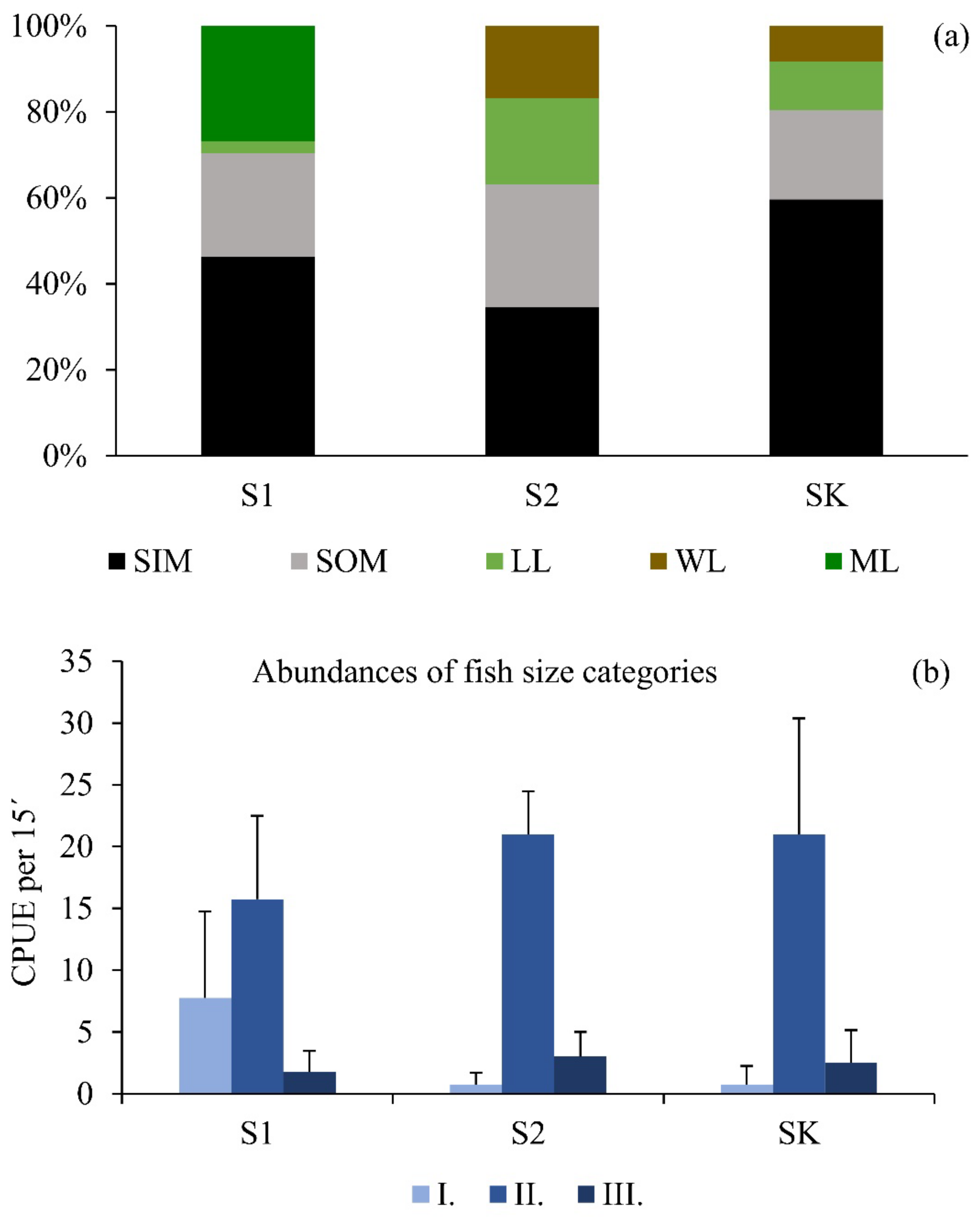
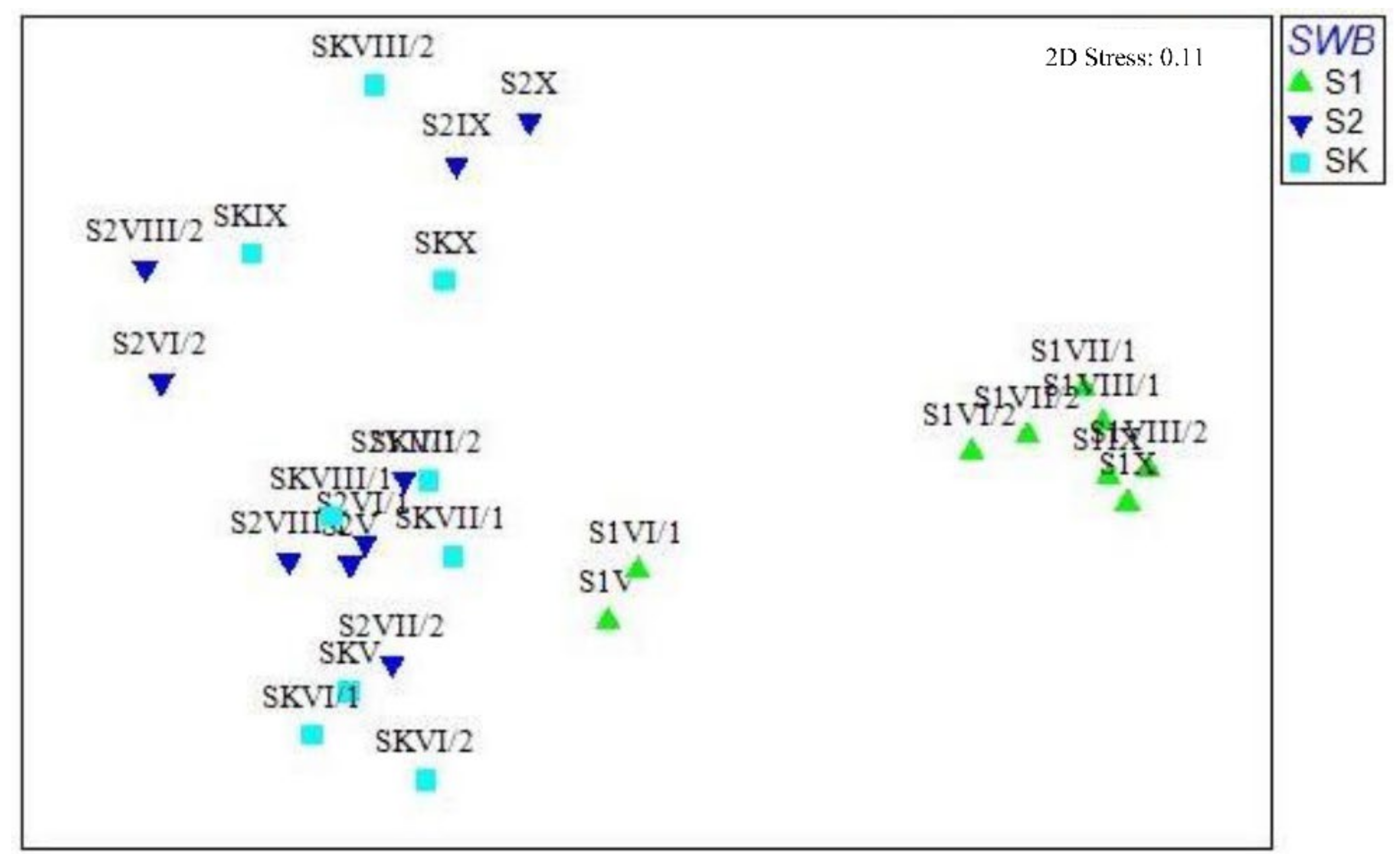
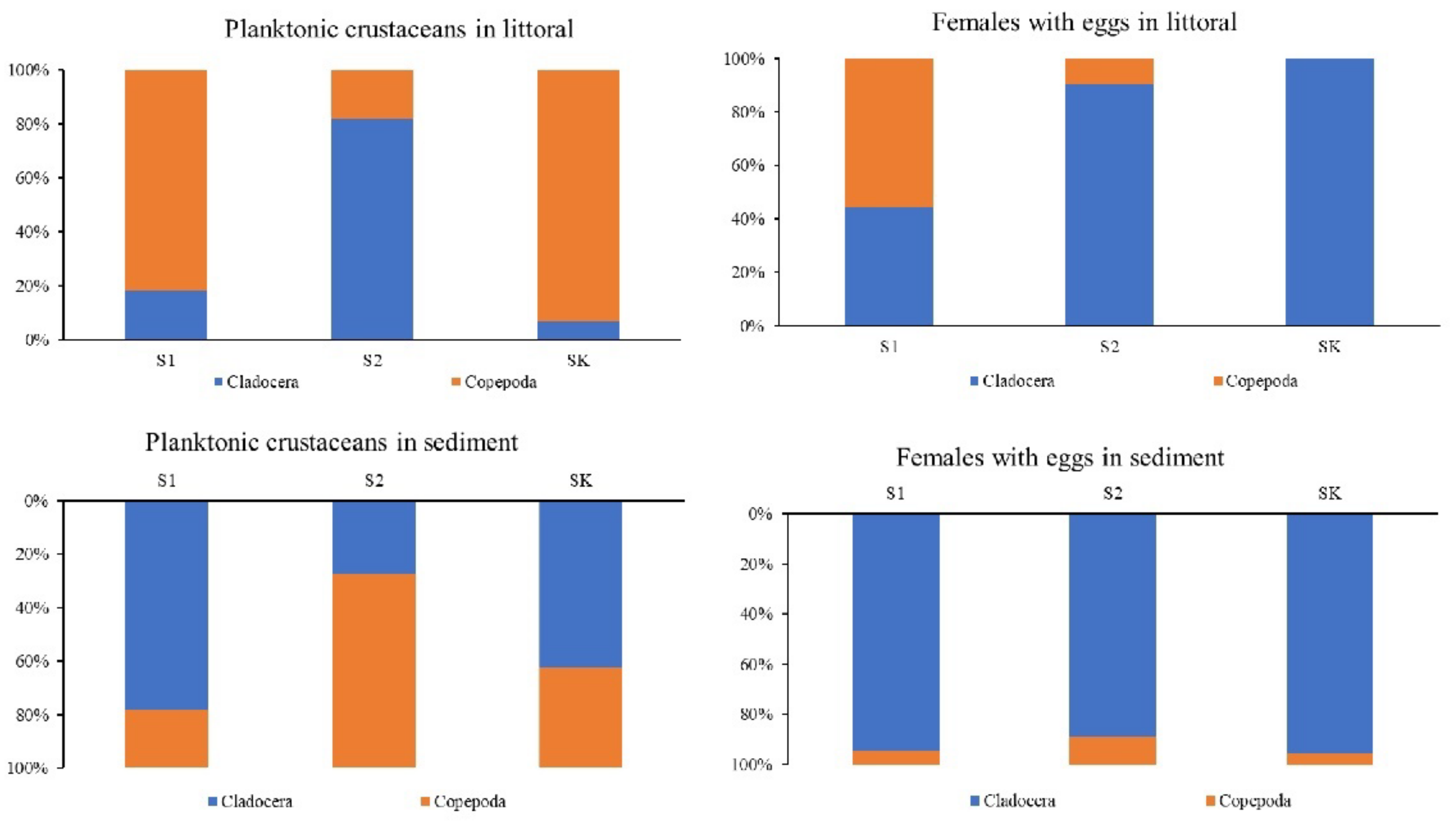
3. Results
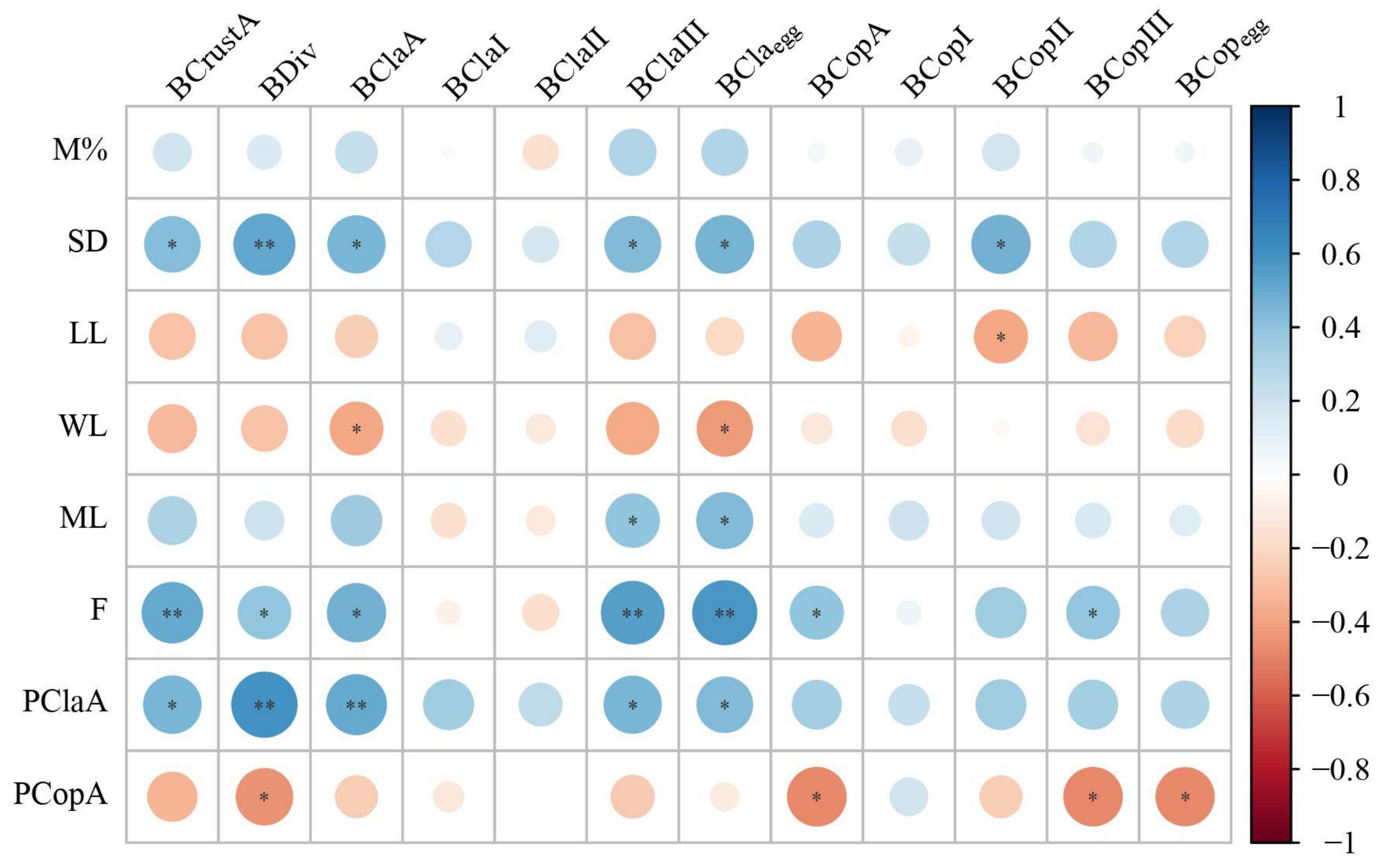
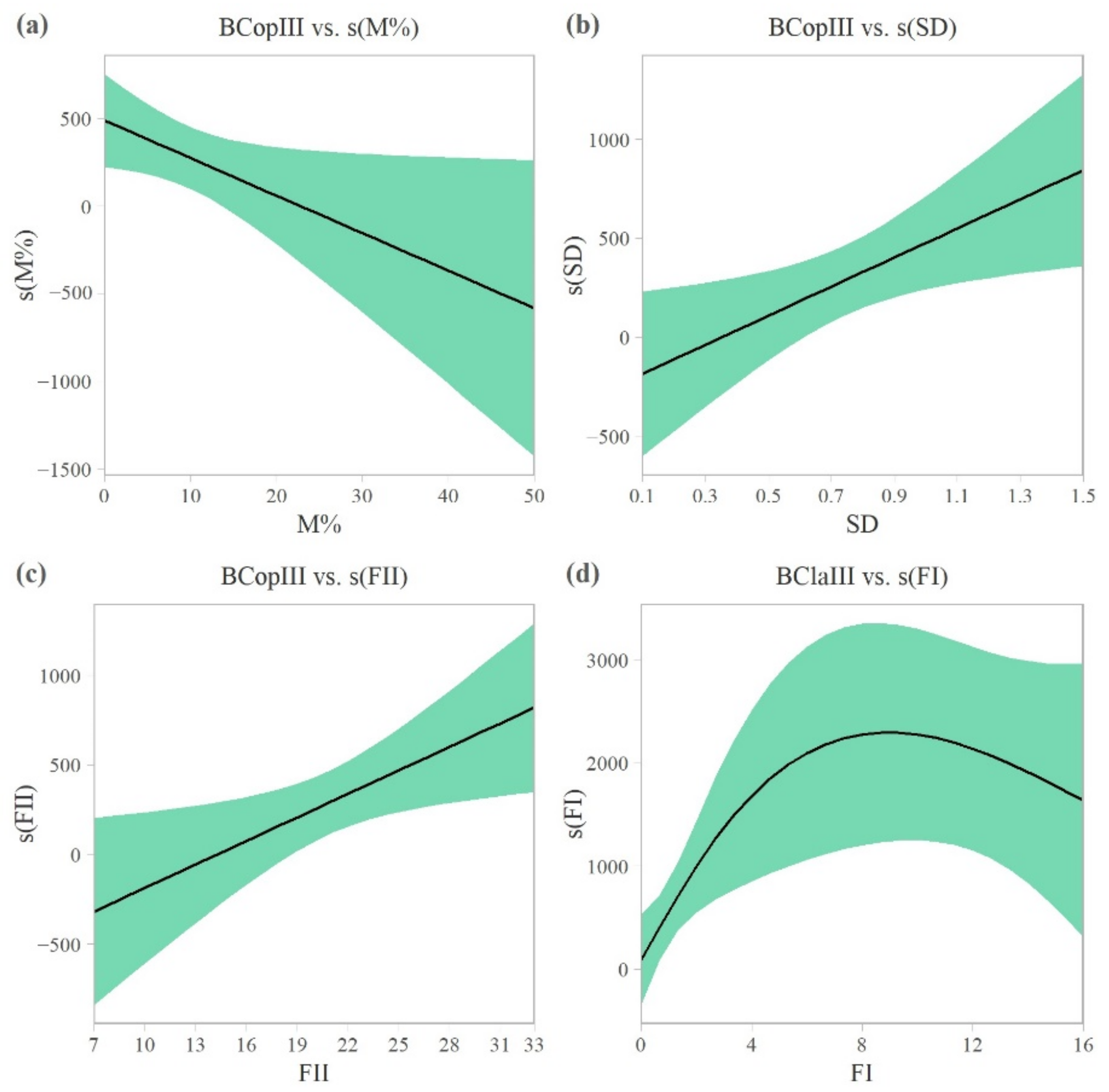
3.1. Abiotic and Biotic Drivers in the Zooplankton Shelter Selection
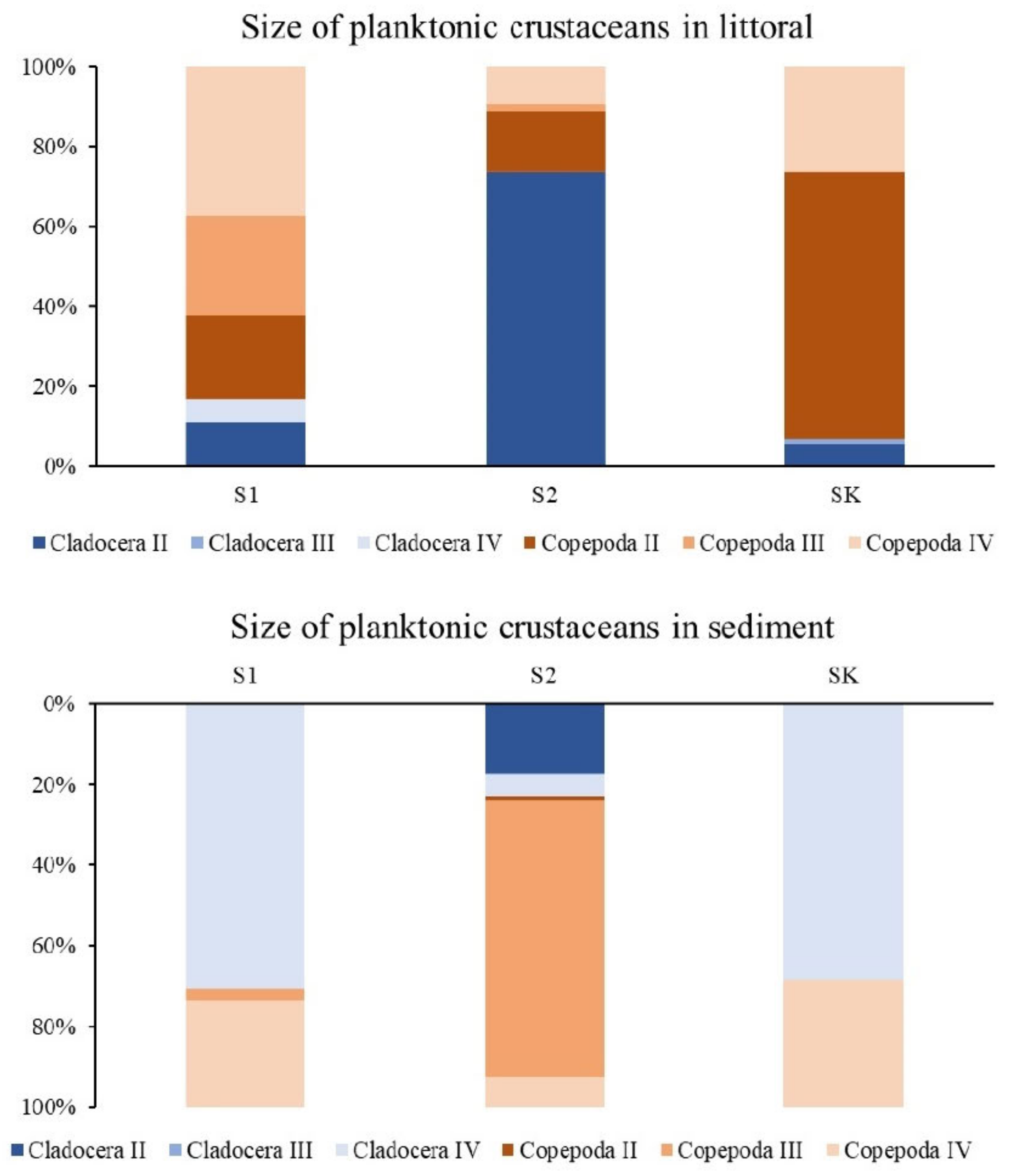
3.2. Planktonic Crustaceans in the Littoral Sediment and Connection with the Littoral Plankton
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sodré, E.D.O.; Bozelli, R.L. How Planktonic Microcrustaceans Respond to Environment and Affect Ecosystem: A Functional Trait Perspective. Int. Aquat. Res. 2019, 11, 207–223. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, E.; Mehner, T.; Winfield, I.J.; Kangur, K.; Sarvala, J.; Gerdeaux, D.; Meerhoff, M. Impacts of climate warming on the long-term dynamics of key fish species in 24 European lakes. Hydrobiologia 2012, 694, 1–39. [Google Scholar] [CrossRef] [Green Version]
- Czerniawski, R.; Krepski, T. Zooplankton Size as a Factor Determining the Food Selectivity of Roach (Rutilus Rutilus) in Water Basin Outlets. Water 2019, 11, 1281. [Google Scholar] [CrossRef] [Green Version]
- Manca, M.; Vijverberg, J.; Polishchuk, L.V.; Voronov, D.A. Daphnia Body Size and Population Dynamics under Predation by Invertebrate and Fish Predators in Lago Maggiore: An Approach Based on Contribution Analysis. J. Limnol. 2008, 67, 15. [Google Scholar] [CrossRef]
- Belmonte, G. The Suspected Contradictory Role of Parental Care in the Adaption of Planktonic Calanoida to Temporary Freshwater. Water 2021, 13, 100. [Google Scholar] [CrossRef]
- Bartholmeé, S.; Samchyshyna, L.; Santer, B.; Lampert, W. Subitaneous eggs of freshwater copepods pass through fish guts: Survival, hatchability, and potential ecological implications. Limnol. Oceanogr. 2005, 50, 923–929. [Google Scholar] [CrossRef]
- Burks, R.; Lodge, D.; Jeppesen, E.; Lauridsen, T. Diel Horizontal migration of zooplankton: Costs and benefits of inhabiting the littoral. Freshw. Biol. 2002, 47, 343–365. [Google Scholar] [CrossRef]
- Meerhoff, M.; Iglesias, C.; De Mello, F.T.; Clemente, J.M.; Jensen, E.; Lauridsen, T.L.; Jeppesen, E. Effects of habitat complexity on community structure and predator avoidance behaviour of littoral zooplankton in temperate versus subtropical shallow lakes. Freshw. Biol. 2007, 52, 1009–1021. [Google Scholar] [CrossRef]
- Tavşanoğlu, Ü.N.; Idil Çakiroğlu, A.; Erdoğan, Ş.; Meerhoff, M.; Jeppesen, E.; Beklioglu, M. Sediments, not plants, offer the preferred refuge for Daphnia against fish predation in Mediterranean shallow lakes: An Experimental Demonstration: Daphnia Refugia in Warm Lakes. Freshw. Biol. 2012, 57, 795–802. [Google Scholar] [CrossRef]
- Ternjej, I.; Plenković Moraj, A.; Mihaljević, Z.; Kerovec, M. Spatial and temporal variation of plankton in a Mediterranean karstic lake. Ekol. Bratisl. 2010, 29, 65–86. [Google Scholar] [CrossRef]
- Cáceres, C.E.; Rogers, D.C. Class Branchiopoda. In Thorp and Covich’s Freshwater Invertebrates-Ecology and General Biology; Thorp, J.H., Rogers, D.C., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2015; pp. 687–708. [Google Scholar]
- Castro, B.B.; Marques, S.M.; Gonçalves, F. Habitat selection and diel distribution of the crustacean zooplankton from a shallow Mediterranean lake during the turbid and clear water phases. Freshw. Biol. 2007, 52, 421–433. [Google Scholar] [CrossRef]
- Špoljar, M.; Tomljanović, T.; Dražina, T.; Lajtner, J.; Štulec, H.; Matulić, D.; Fressl, J. Zooplankton structure in two interconnected ponds: Similarities and differences. Croat. J. Fish. 2016, 74, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Son, S.-H.; Kwon, S.-J.; Im, J.-H.; Kim, S.-K.; Kong, D.; Choi, J.-Y. Aquatic Macrophytes Determine the Spatial Distribution of Invertebrates in a Shallow Reservoir. Water 2021, 13, 1455. [Google Scholar] [CrossRef]
- Jeppesen, E.; Jensen, J.P.; Søndergaard, M.; Lauridsen, T.; Pedersen, L.J.; Jensen, L. Top-down control in freshwater lakes: The role of nutrient state, submerged macrophytes and water depth. Hydrobiologia 1997, 342/343, 151–164. [Google Scholar] [CrossRef]
- Špoljar, M.; Dražina, T.; Lajtner, J.; Duić Sertić, M.; Radanović, I.; Wallace, R.L.; Matulić, D.; Tomljanović, T. Zooplankton assemblage in four temperate shallow waterbodies in association with habitat heterogeneity and alternative states. Limnologica 2018, 71, 51–61. [Google Scholar] [CrossRef]
- Špoljar, M.; Lajtner, J.; Dražina, T.; Malekzadeh Viayeh, R.; Radanović, I.; Zrinščak, I.; Fressl, J.; Matijašec, D. Disentangling food webs interactions in the littoral of temperate shallow lakes. Nat. Croat. 2017, 26, 145–166. [Google Scholar] [CrossRef] [Green Version]
- González Sagrario, M.A.; de losÁNGELES, M.; Balseiro, E.; Ituarte, R.; Spivak, E. Macrophytes as refuge or risky area for zooplankton: A balance set by littoral predacious macroinvertebrates. Freshw. Biol. 2009, 54, 1042–1053. [Google Scholar] [CrossRef]
- Špoljar, M.; Dražina, T.; Šargač, J.; Kralj Borojević, K.; Žutinić, P. Submerged macrophytes as a habitat for zooplankton development in two reservoirs of a flow-through system (Papuk nature park, Croatia). Ann. Limnol. Int. J. Limnol. 2012, 48, 161–175. [Google Scholar] [CrossRef] [Green Version]
- Meerhoff, M.; Clemente, J.M.; de Mello, F.T.; Iglesias, C.; Pedersen, A.R.; Jeppesen, E. Can warm climate-related structure of littoral predator assemblies weaken the clear water state in shallow lakes? Glob. Chang. Biol. 2007, 13, 1888–1897. [Google Scholar] [CrossRef]
- Kuczyńska-Kippen, N.; Špoljar, M.; Mirosław, M.; Zhang, C. Elodeids, but not helophytes, increase community diversity and reduce trophic state: Case study with rotifer indices in field ponds. Ecol. Indic. 2021, 128, 107829. [Google Scholar] [CrossRef]
- Kuczyńska-Kippen, N.; Joniak, T. Zooplankton diversity and macrophyte biometry in shallow water bodies of various trophic state. Hydrobiologia 2016, 774, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Kolarova, N.; Napiórkowski, P. How do specific environmental conditions in canals affect the structure and variability of the zooplankton community? Water 2022, 14, 979. [Google Scholar] [CrossRef]
- Compte, J.; Montenegro, M.; Ruhí, A.; Gascón, S.; Sala, J.; Boix, D. Microhabitat selection and diel patterns of zooplankton in a Mediterranean temporary pond. Hydrobiologia 2015, 766, 201–213. [Google Scholar] [CrossRef]
- Fressl, J. Impact of Submersed Macrophytes and Fish Predation on the Structure of Zooplankton in Shallow Eutrophic Lakes. Ph.D. Dissertation, University of Zagreb, Zagreb, Croatia, 21 September 2020. [Google Scholar]
- Špoljar, M.; Perić, M.S.; Wang, H.; Zhang, C.; Kuczyńska-Kippen, N.; Fressl, J.; Ercegovac, Z. Does the size structure of the littoral community reflect water level fluctuations in shallow waterbodies? Ecol. Indic. 2021, 132, 108330. [Google Scholar] [CrossRef]
- Riley, W.D.; Potter, E.C.E.; Biggs, J.; Collins, A.L.; Jarvie, H.P.; Jones, J.I.; Kelly-Quinn, M.; Ormerod, S.J.; Sear, D.A.; Wilby, R.L.; et al. Small Water Bodies in Great Britain and Ireland: Ecosystem function, human-generated degradation, and options for restorative action. Sci. Total Environ. 2018, 645, 1598–1616. [Google Scholar] [CrossRef] [PubMed]
- Amoros, C. Crustaces Cladoceres. Bull. Mens. Soc. Linn. Lyon. 1984, 3–4, 1–63. [Google Scholar]
- Einsle, U. Crustacea: Copepoda: Calanoida und Cyclopoida. In Süsswasserfauna von Mitteleuropa; Schwoerbel, J., Zwick, P., Eds.; Gustav Fischer Verlag: Stuttgart, Germany, 1993; Volume 8/4–1. [Google Scholar]
- Kottelat, M.; Freyhof, J. Handbook of European Freshwater Fishes; Kottelat: Cornol, Switzerland; Freyhof: Berlin, Germany, 2007. [Google Scholar]
- APHA Standard Methods for the Examination of Water and Wastewater, 20th ed.; American Public Health Association: Washington, DC, USA, 1998.
- Nusch, E.A. Comparison of different methods for chlorophyll and phaeopigment determination. Arch. Für Hydrobiol. 1980, 14, 14–36. [Google Scholar]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation; PRIMER-E: Plymouth, UK, 2001. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. Primer v6: User Manual/Tutorial (Plymouth Routines in Multivariate Ecological Research); PRIMER-E: Plymouth, UK, 2006. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org (accessed on 15 February 2022).
- Sakamoto, Y.; Ishiguro, M.; Kitagawa, G. Akaike Information Criterion Statistics, 1st ed.; Springer: Dordrecht, The Netherlands, 1986. [Google Scholar]
- Jin, B.S.; Winemiller, K.O.; Shao, B.; Si, J.K.; Jin, J.F.; Ge, G. Fish assemblage structure in relation to seasonal environmental variation in sub-lakes of the Poyang Lake floodplain, China. Fish. Manag. Ecol. 2019, 26, 131–140. [Google Scholar] [CrossRef]
- Mooij, W.M.; Janse, J.H.; De Senerpont Domis, L.N.; Hülsmann, S.; Ibelings, B.W. Predicting the effect of climate change on temperate shallow lakes with the ecosystem model PCLake. Hydrobiologia 2007, 584, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Molinos, J.G.; Heino, J.; Zhang, H.; Zhang, P.; Xu, J. Eutrophication causes invertebrate biodiversity loss and decreases cross-taxon congruence across anthropogenically-disturbed lakes. Environ. Int. 2021, 153, 106494. [Google Scholar] [CrossRef]
- Pestić, A. Biotic Impact on Zooplankton Distribution in the Eutrophic Lake. Master’s Thesis, University of Zagreb, Zagreb, Croatia, 26 February 2015. (In Croatian). [Google Scholar]
- Iglesias, C.; Goyenola, G.; Mazzeo, N.; Meerhoff, M.; Rodo, E.; Jeppesen, E. Horizontal dynamics of Zooplankton in subtropical Lake Bianca (Uruguay) hosting multiple Zooplankton predators and aquatic plant refuges. In Shallow Lakes in a Changing World; Gulati, R.D., Lammens, E., DePauw, N., Van Donk, E., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 179–189. [Google Scholar]
- Kuczyńska-Kippen, N.; Pronin, M. Diversity and zooplankton species associated with certain hydroperiods and fish state in field ponds. Ecol. Ind. 2018, 90, 171–178. [Google Scholar] [CrossRef]
- Cáceres, C.E.; Hairston, N.G., Jr. Benthic-pelagic coupling in planktonic crustaceans: The role of the benthos. Arch. Hydrobiol. 1998, 52, 163–174. [Google Scholar]
- Iglesias, C.; Mazzeo, N.; Meerhoff, M.; Lacerot, G.; Clemente, J.M.; Scasso, F.; Jeppesen, E. High predation is of key importance for dominance of small-bodied zooplankton in warm shallow lakes: Evidence from lakes, fish exclosures and surface sediments. Hydrobiologia 2011, 667, 133–147. [Google Scholar] [CrossRef]
- Olmo, C.; Antón-Pardo, M.; Ortells, R.; Armengol, X. Influence of restoration age on egg bank richness and composition: An ex situ experiment. J. Plankton Res. 2020, 42, 553–563. [Google Scholar] [CrossRef]
- Basińska, A.M.; Świdnicki, K.; Kuczyńska-Kippen, N. Effect of surrounding trees and dry rush presence on spring zooplankton community in an urban pond complex. Ann. Limnol. Int. J. Lim. 2014, 50, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Slusarczyk, M.; Pinel-Alloul, B.; Pietrzak, B. Mechanisms Facilitating Dispersal of Dormant Eggs in a Planktonic Crustacean. In Dormancy in Aquatic Organisms. Theory, Human Use and Modeling; Alekseev, V., Pinel-Alloul, B., Eds.; Monographiae Biologicae; Springer: Cham, Switzerland, 2019; Volume 92. [Google Scholar] [CrossRef]
- Ringelberg, J.; Van Gool, E. On the combined analysis of proximate and ultimate aspects in diel vertical migration (DVM) research. Hydrobiologia 2003, 491, 85–90. [Google Scholar] [CrossRef]
- Battauz, Y.S.; de Paggi, S.B.J.; Paggi, J.C. Macrophytes as dispersal vectors of zooplankton resting stages in a subtropical riverine floodplain. Aquat. Ecol. 2017, 51, 191–201. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H | p | Post-Hoc Test | |
|---|---|---|---|
| Sediment type | |||
| LL (g m−2) | 10.88 | 0.0043 | S1 < S2, SK |
| ML (g m−2) | 17.68 | 0.0001 | S1 > S2, SK |
| Fish | |||
| Total fish abundance | 8.856 | 0.0119 | SK > S1, S2 |
| Fish I | 13.59 | 0.0011 | S1 > S2, SK |
| Average fish length (cm) | 10.66 | 0.0048 | S1 < S2, SK |
| Littoral Sediment | |||
| No. of taxa | 6.73 | 0.0346 | SK < S1 |
| Total Crustacea (Ind. m−2) | 7.89 | 0.0193 | SK < S2 |
| Cladocera females with eggs (Ind. m−2) | 8.24 | 0.0163 | SK < S2 |
| Taxa/Station | S1 | S2 | SK | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Cladocera | |||||||||
| Bosmina longirostris (O.F.Müller, 1776) | 512 | ± | 540 | ||||||
| B. longirostris with eggs | 339 | ± | 358 | ||||||
| Ceriodaphnia pulchella | 13 | ± | 13 | ||||||
| Daphnia curvirostris Eylmann, 1887 | 38 | ± | 40 | ||||||
| D. curvirostris with eggs | 19 | ± | 20 | ||||||
| Ilyocryptus agilis Kurz, 1878 | 1 | ± | 1 | 1 | ± | 1 | |||
| Pleuroxus sp. | 1 | ± | 1 | ||||||
| Simocephalus vetulus (O.F. Müller, 1776) | 773 | ± | 279 | 256 | ± | 105 | 149 | ± | 146 |
| S. vetulus with eggs | 223 | ± | 108 | 68 | ± | 36 | 9 | ± | 8 |
| Simocephalus exspinosus (Koch, 1841) | 261 | ± | 79 | 159 | ± | 79 | 100 | ± | 104 |
| S. exspinosus with eggs | 77 | ± | 38 | 39 | ± | 18 | 5 | ± | 5 |
| Simocephalus serrulatus (Koch, 1841) | 12 | ± | 11 | ||||||
| S. serrulatus with eggs | 10 | ± | 10 | ||||||
| Cladocera total | 1031 | ± | 350 | 978 | ± | 594 | 260 | ± | 160 |
| Cladocera total eggs | 262 | ± | 127 | 469 | ± | 368 | 24 | ± | 23 |
| Cyclopoida | |||||||||
| Acanthocyclops robustus (Sars, 1863) | 88 | ± | 93 | ||||||
| A. robustus with eggs | 2 | ± | 3 | ||||||
| Cyclops sp. | 35 | ± | 17 | 61 | ± | 64 | 96 | ± | 66 |
| Cyclops sp. with eggs | 1 | ± | 1 | 7 | ± | 8 | 1 | ± | 1 |
| Ectocyclops sp. | 1 | ± | 1 | ||||||
| Macrocyclops albidus (Jurine, 1820) | 44 | ± | 25 | 51 | ± | 27 | 64 | ± | 60 |
| M. albidus with eggs | 44 | ± | 25 | 30 | ± | 19 | |||
| Macrocyclops fuscus (Jurine, 1820) | 3 | ± | 3 | ||||||
| Mesocyclops leuckarti (Claus, 1857) | 218 | ± | 229 | ||||||
| M. leuckarti with eggs | 38 | ± | 40 | ||||||
| nauplii | 1 | ± | 1 | 26 | ± | 27 | |||
| copepodites | 26 | ± | 13 | 2265 | ± | 2292 | |||
| Cyclopoida total | 285 | ± | 134 | 2621 | ± | 2539 | 154 | ± | 118 |
| Cyclopoida total eggs | 15 | ± | 4 | 61 | ± | 40 | 1 | ± | 1 |
| Calanoida | |||||||||
| Eudiaptomus gracilis (Sars, 1863) | 6 | ± | 7 | 3 | ± | 3 | |||
| Calanoida total | 6 | ± | 7 | 3 | ± | 3 | |||
| Harpacticoida total | 3 | ± | 3 | ||||||
| Ostracoda total | 2 | ± | 2 | 10 | ± | 7 | 9 | ± | 9 |
| Model Terms | Family: Gaussian | Scale est. 5.17 × 105 | n = 27 | |||
|---|---|---|---|---|---|---|
| BClaIII abundance | Parametric coefficients | Estimate | Std. Error | T value | ||
| (Intercept) | 647.1 | 138.4 | 4.678 | <0.001 | ||
| Aproximate significance of smooth terms | s (FI) | s (FII) | s (FIII) | s (M%) | ||
| p< 0.05 | - | - | - | |||
| R2adj | 0.338 | |||||
| Deviance explained (%) | 47.2 | |||||
| Model Terms | Family: Gaussian | Scale est. 2.19 × 105 | n = 27 | |||
| BCopIII abundance | Parametric coefficients | Estimate | Std. Error | T value | ||
| (Intercept) | 283.56 | 90.21 | 3.143 | <0.01 | ||
| Aproximate significance of smooth terms | s (FI) | s (FII) | s (FIII) | s (M%) | s (SD) | |
| - | <0.05 | - | 0.05 | <0.05 | ||
| R2adj | 0.198 | |||||
| Deviance explained (%) | 37.3 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Špoljar, M.; Kuczyńska-Kippen, N.; Dražina, T.; Fressl, J.; Ternjej, I.; Tomljanović, T.; Zhang, C.; Purgar, M.; Čorkalo, M.; Kekelj, L. Sediment as a Refuge Spot for Planktonic Crustaceans. Water 2022, 14, 1680. https://doi.org/10.3390/w14111680
Špoljar M, Kuczyńska-Kippen N, Dražina T, Fressl J, Ternjej I, Tomljanović T, Zhang C, Purgar M, Čorkalo M, Kekelj L. Sediment as a Refuge Spot for Planktonic Crustaceans. Water. 2022; 14(11):1680. https://doi.org/10.3390/w14111680
Chicago/Turabian StyleŠpoljar, Maria, Natalia Kuczyńska-Kippen, Tvrtko Dražina, Jelena Fressl, Ivančica Ternjej, Tea Tomljanović, Chen Zhang, Marija Purgar, Marko Čorkalo, and Lana Kekelj. 2022. "Sediment as a Refuge Spot for Planktonic Crustaceans" Water 14, no. 11: 1680. https://doi.org/10.3390/w14111680
APA StyleŠpoljar, M., Kuczyńska-Kippen, N., Dražina, T., Fressl, J., Ternjej, I., Tomljanović, T., Zhang, C., Purgar, M., Čorkalo, M., & Kekelj, L. (2022). Sediment as a Refuge Spot for Planktonic Crustaceans. Water, 14(11), 1680. https://doi.org/10.3390/w14111680