
Northward Expansion of a Warm-Water Doliolid Dolioletta gegenbauri (Uljanin, 1884) into a Temperate Bay, China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Environmental Data
2.2. Sampling
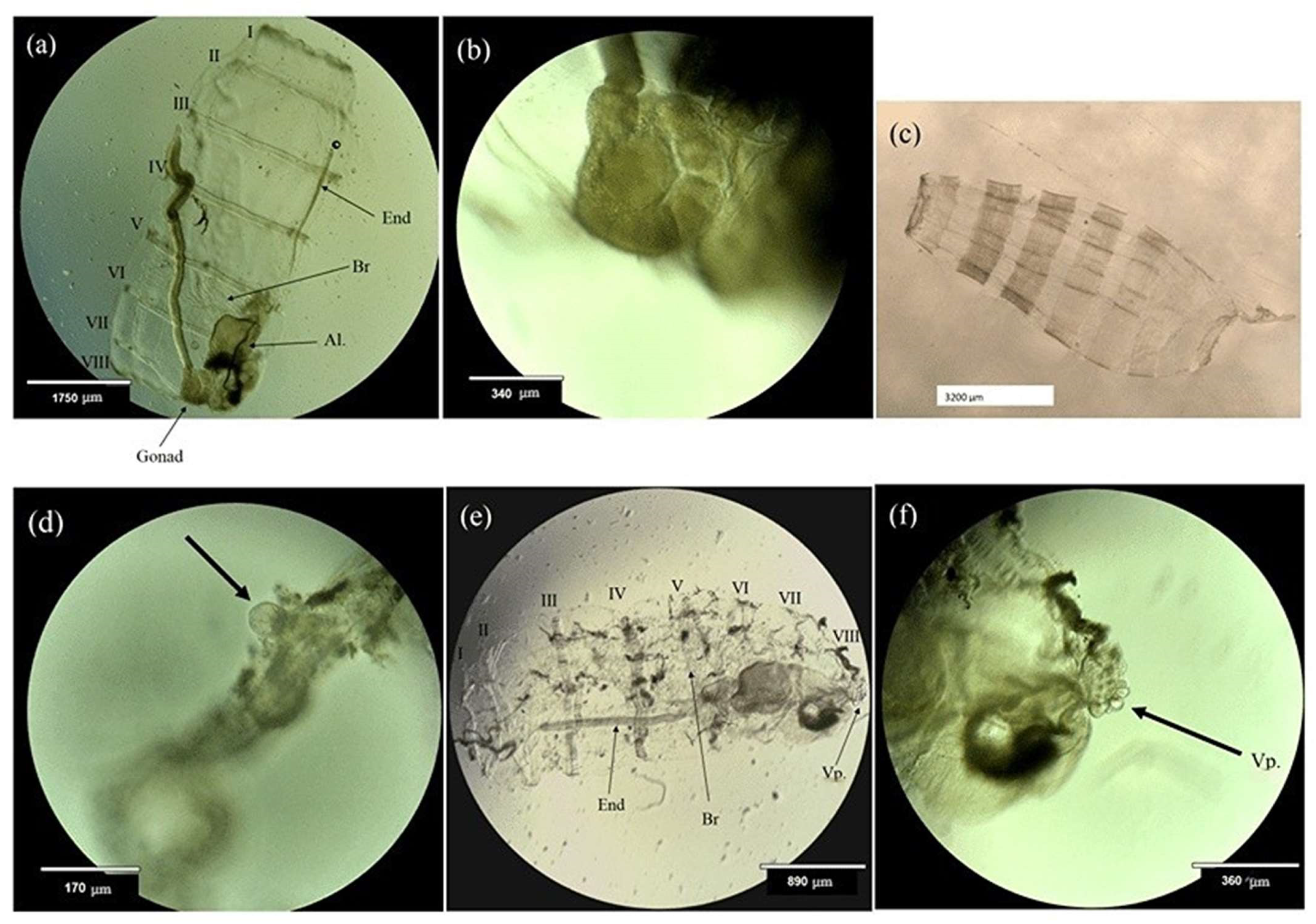
2.3. Body Size, Life History Stage, and Abundance
2.4. Role of Environmental Factors
3. Results
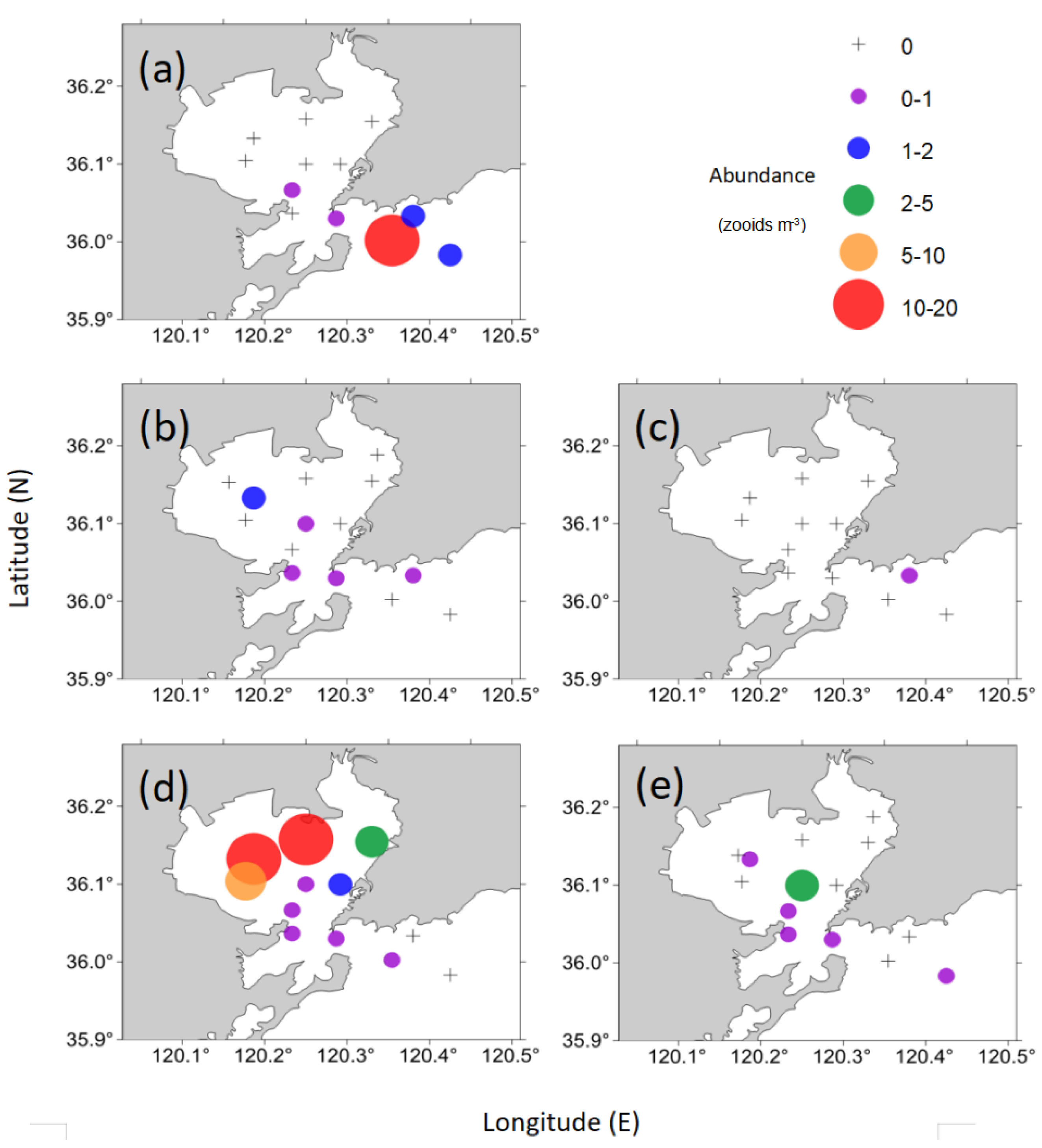
3.1. Abundance and Distribution
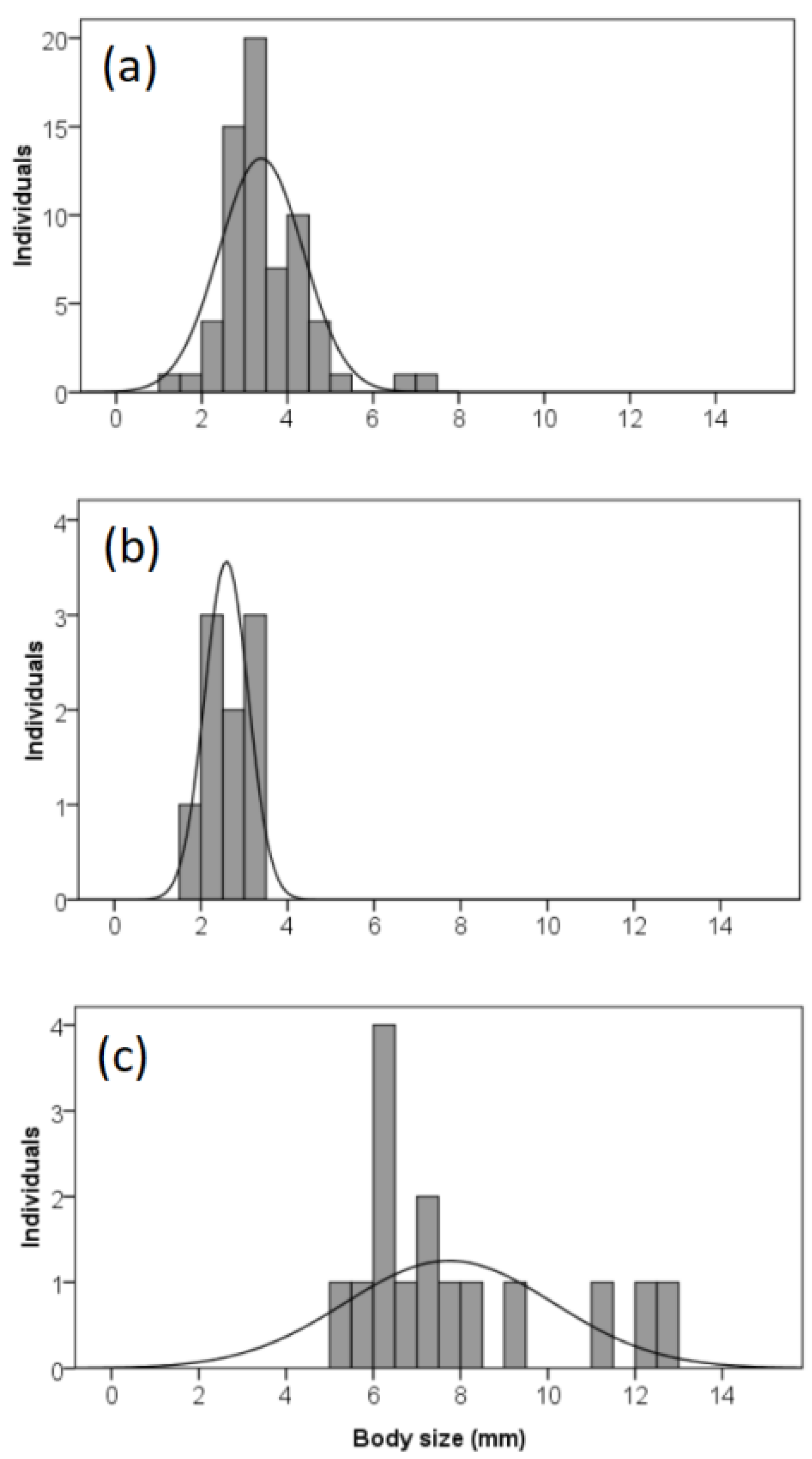
3.2. Life History Stage and Body Size Compositions
3.3. Temperature and Salinity Long-Term Change
3.4. Relationship with Temperature, Salinity and Food
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deibel, D.; Lowen, D. A review of the life cycles and life-history adaptations of pelagic tunicates to environmental conditions. ICES J. Mar. Sci. 2012, 69, 358–369. [Google Scholar] [CrossRef]
- Paffenhöfer, G.-A.; Köster, M. From one to many: On the life cycle of Dolioletta gegenbauri Uljanin (Tunicata, Thaliacea). J. Plankton Res. 2011, 33, 1139–1145. [Google Scholar] [CrossRef] [Green Version]
- Lucas, C.H.; Dawson, M.N. What Are Jellyfishes and Thaliaceans and Why Do They Bloom? In Jellyfish Blooms; Pitt, K., Lucas, C., Eds.; Springer: Dordrecht, The Netherland, 2014. [Google Scholar]
- Takahashi, K.; Ichikawa, T.; Fukugama, C.; Yamane, M.; Kakehi, S.; Okazaki, Y.; Kubota, H.; Furuya, K. In situ observations of a doliolid bloom in a warm water filament using a video plankton recorder: Bloom development, fate, and effect on biogeochemical cycles and planktonic food webs. Limnol. Oceanogr. 2015, 60, 1763–1780. [Google Scholar] [CrossRef] [Green Version]
- Paffenhofer, G.-A.; Gibson, D.M. Determination of generation time and asexual fecundity of doliolids (Tunicata, Thaliacea). J. Plankton Res. 1999, 21, 1183–1189. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Yin, J.; Huang, L.; Zhang, J.; Lian, S.; Liu, C. Distribution and abundance of thaliaceans in the northwest continental shelf of South China Sea, with response to environmental factors driven by monsoon. Cont. Shelf Res. 2011, 31, 979–989. [Google Scholar] [CrossRef]
- He, D.; Yang, G.; Fang, S.; Shen, W.; Liu, H.; Gao, A.; Huang, S. Study of zooplankton ecology in Zhejiang coastal upwelling system-zooplankton biomass and abundance of major groups. Acta Oceanol. Sinica 1988, 7, 607–620. [Google Scholar]
- Xu, Z.; Lin, M.; Zhang, J. Relationship of water environment and abundance distribution of thaliacea in the East China Sea. Ocean. Limnol. Sinica 2007, 38, 549–554. [Google Scholar]
- Xu, Z.; Sun, J.; Lin, M. Statistical analysis to ecological group of Thaliacea in the East China Sea. Acta Ecol. Sinica 2008, 28, 5698–5705. [Google Scholar]
- Liu, L.R. Checklist of Marine Biota of China Seas; China Science Press: Beijing, China, 2008; p. 870. [Google Scholar]
- Sun, S.; Huo, Y.; Yang, B. Zooplankton functional groups on the continental shelf of the yellow sea. Deep Sea Res. II Top. Stud. Oceanogr. 2010, 57, 1006–1016. [Google Scholar] [CrossRef]
- Franco, P.; Chen, H.; Liu, G. Distribution and abundance of pelagic tunicates in the North Yellow Sea. J. Ocean Univ. China 2014, 13, 782–790. [Google Scholar] [CrossRef]
- Wang, X. Studies on Zooplankton Community and the Effects of Environmental Factors on Its Distribution in the Southern Yellow Sea. Ph.D. Thesis, University of China, Qingdao, China, 2012. [Google Scholar]
- Sun, S.; Sun, X.; Zhang, G.; Tang, H.; Liu, Q.; Li, G. Long-term changes in major meteorological and hydrological factors in the Jiaozhou Bay. Oceanol. Limnol. Sin. 2011, 42, 632–638. [Google Scholar]
- Sun, S.; Li, C.; Zhang, G.; Sun, X.; Yang, B. Long-term changes in the zooplankton community in the Jiaozhou Bay. Oceanol. Limnol. Sin. 2011, 42, 625–671. [Google Scholar]
- Ma, X.; Sun, S.; Gao, S. Ecology of jellyfishes in Jiaozhou Bay-Seasonal and annual changes in community structure. Stud. Mar. Sin. 2000, 42, 91–99. [Google Scholar]
- Wang, S.; Zhang, G.; Zhou, K.; Sun, S. Long-term population variability and reproductive strategy of a northward expanded ctenophore Pleurobrachia globosa Moser, 1903 in a temperate bay, China. J. Exp. Mar. Biol. Ecol. 2020, 533, 151457. [Google Scholar] [CrossRef]
- Adam, N.A.; Ishak, N. Taxonomic revision of the family Doliolidae Bronn, 1862 (Chordata, Tunicata, Thaliacea, Doliolida) from east coast of Peninsular Malaysia), with an updated worldwide distribution. J. Sustain. Sci. Manag. 2018, 13, 147–178. [Google Scholar]
- Deibel, D.; Paffenhöfer, G.-A. Predictability of patches of neritic salps and doliolids (Tunicata, Thaliacea). J. Plankton Res. 2009, 31, 1571–1579. [Google Scholar] [CrossRef] [Green Version]
- Su, J. Circulation dynamics of the China Seas North of 18 N. In The Sea; Robinson, A.R., Brink, K.H., Eds.; Wiley: New York, NY, USA, 1998; Volume 11, pp. 483–505. [Google Scholar]
- Liu, Z.; Gan, J.; Hu, J.; Wu, H.; Cai, Z.; Deng, Y. Progress on circulation dynamics in the East China Sea and southern Yellow Sea: Origination, pathways, and destinations of shelf currents. Progr. Oceanogr. 2021, 193, 102553. [Google Scholar] [CrossRef]
- Tang, Y.; Zhou, E.; Lie, H. On the origin and path of the Huanghai Warm Current during winter and early spring. Acta Oceanol. Sin. 2001, 23, 1–12. [Google Scholar]
- Wang, R.; Zuo, T. The Yellow Sea Warm Current and the Yellow Sea Cold Bottom Water, their impact on the distribution of zooplankton in the southern Yellow Sea. J. Soc. Oceanogr. Korean 2004, 39, 1–13. [Google Scholar]
- Zuo, T.; Wang, R.; Chen, Y.; Gao, S.; Wang, K. Autumn net copepod abundance and assemblages in relation to water masses on the continental shelf of the Yellow Sea and East China Sea. J. Mar. Syst. 2006, 59, 159–172. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Z.; Chen, D.; Qiu, B.; Wang, W. Strengthening of the Kuroshio current by intensifying tropical cyclones. Science 2020, 368, 988–993. [Google Scholar] [CrossRef]
- Poloczanska, E.S.; Brown, C.J.; Sydeman, W.J.; Kiessling, W.; Schoeman, D.S.; Moore, P.J.; Brander, K.; Bruno, J.F.; Buckley, L.B.; Burrows, M.T.; et al. Global imprint of climate change on marine life. Nature Clim. Change 2013, 3, 919–925. [Google Scholar] [CrossRef]
- Lin, C.; Ning, X.; Su, J.; Lin, Y.; Xu, B. Environmental changes and the responses of the ecosystems of the Yellow Sea during 1976–2000. J. Mar. Syst. 2005, 55, 223–234. [Google Scholar] [CrossRef]
- Jung, H.K.; Rahman, S.M.; Kang, C.K.; Park, S.Y.; Lee, S.H.; Park, H.J.; Kim, H.W.; Lee, C.I. The influence of climate regime shifts on the marine environment and ecosystems in the East Asian Marginal Seas and their mechanisms. Deep Sea Res. Part II 2017, 143, 110–120. [Google Scholar] [CrossRef]
- Cheng, L.; Abraham, J.; Zhu, J.; Trenberth, K.E.; Fasullo, J.; Boyer, T.; Locarnini, R.; Zhang, B.; Yu, F.; Wan, L.; et al. Record-Setting Ocean Warmth Continued in 2019. Adv. Atmos. Sci. 2019, 37, 137–142. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.; Abraham, J.; Trenberth, K.E.; Fasullo, J.; Boyer, T.; Locarnini, R.; Zhang, B.; Yu, F.; Wan, L.; Chen, X.; et al. Upper Ocean Temperatures Hit Record High in 2020. Adv. Atmos. Sci. 2019, 38, 523–530. [Google Scholar] [CrossRef]
- Paffenhöfer, G.-A.; Atkinson, L.P.; Lee, T.N.; Verity, P.G.; Bulluck, L.R. Distribution and abundance of thaliaceans and copepods off the southeastern U.S.A. during winter, 1995. Cont. Shelf Res. 1995, 15, 225–280. [Google Scholar] [CrossRef]
- Li, S.; Zhang, G. Role of intraspecific competition in intrinsic growth rate regulation in an Oikopleura dioica (Tunicata) population. J. Ocean Limnol. 2021, 39, 609–622. [Google Scholar] [CrossRef]
- Harbison, G.R.; Bone, Q. The parasites and predators of Thaliacea. In The Biology of Pelagic Tunicates; Oxford University Press: Oxford, UK, 1998; pp. 187–214. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Life Stages | 2019 | 2020 | |||
|---|---|---|---|---|---|
| Oct | Nov | Dec | Oct | Nov | |
| Nurse | 19.8 ± 7.1 | 16.2 ± 10.0 | 100.0 | 21.8 ± 8.3 | 3.0 ± 2.6 |
| Gonozooid | 70.7 ± 14.2 | 83.8 ± 10.0 | NA | 62.0 ± 18.0 | 87.8 ± 9.9 |
| Phorozooid | 9.5 ± 7.6 | NA | NA | 13.5 ± 9.3 | 8.4 ± 9.7 |
| Larva | NA | NA | NA | 2.0 ± 3.8 | NA |
| Correlation Coefficient | p | n | |
|---|---|---|---|
| A vs. T | 0.35 | 0.005 * | 64 |
| A vs. S | −0.09 | 0.496 | 64 |
| A vs. Chl a | −0.17 | 0.179 | 64 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Wan, A.; Zhang, G.; Sun, S. Northward Expansion of a Warm-Water Doliolid Dolioletta gegenbauri (Uljanin, 1884) into a Temperate Bay, China. Water 2022, 14, 1685. https://doi.org/10.3390/w14111685
Wang S, Wan A, Zhang G, Sun S. Northward Expansion of a Warm-Water Doliolid Dolioletta gegenbauri (Uljanin, 1884) into a Temperate Bay, China. Water. 2022; 14(11):1685. https://doi.org/10.3390/w14111685
Chicago/Turabian StyleWang, Shiwei, Aiyong Wan, Guangtao Zhang, and Song Sun. 2022. "Northward Expansion of a Warm-Water Doliolid Dolioletta gegenbauri (Uljanin, 1884) into a Temperate Bay, China" Water 14, no. 11: 1685. https://doi.org/10.3390/w14111685
APA StyleWang, S., Wan, A., Zhang, G., & Sun, S. (2022). Northward Expansion of a Warm-Water Doliolid Dolioletta gegenbauri (Uljanin, 1884) into a Temperate Bay, China. Water, 14(11), 1685. https://doi.org/10.3390/w14111685