
Phytoplankton and Bacterial Communities’ Patterns in a Highly Dynamic Ecosystem (Central Mediterranean Sea)

 ,
, 
 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling
2.3. Physical–Chemical Parameters
2.4. Total and Size-Fractionated Chlorophyll a and Primary Production Rates
2.5. Prokaryotic Abundance and Morphotypes
2.6. Larger Phytoplankton Abundance and Species Composition
2.7. Extracellular Enzyme Activity Rates (EEA)
2.8. Data Processing
2.8.1. Statistical Analysis
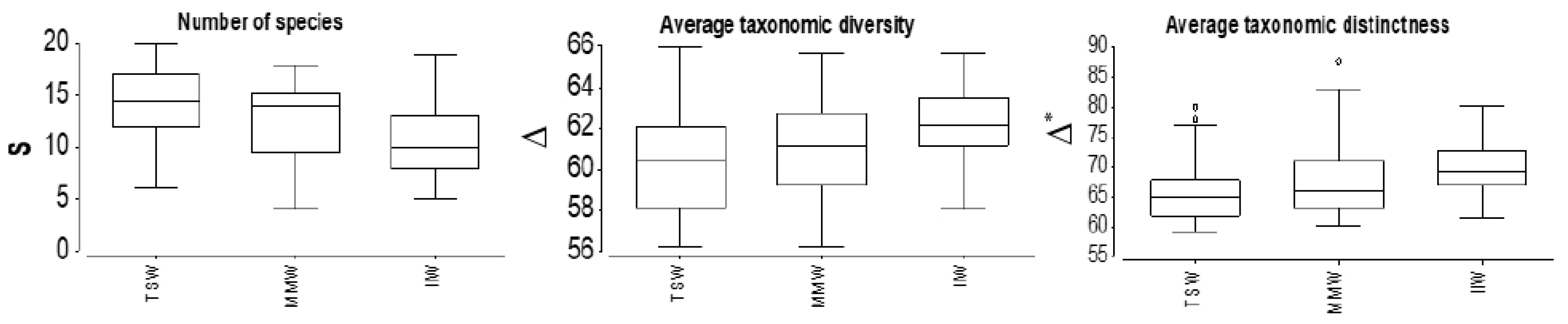
2.8.2. Phytoplankton Community Structure and Biodiversity
3. Results
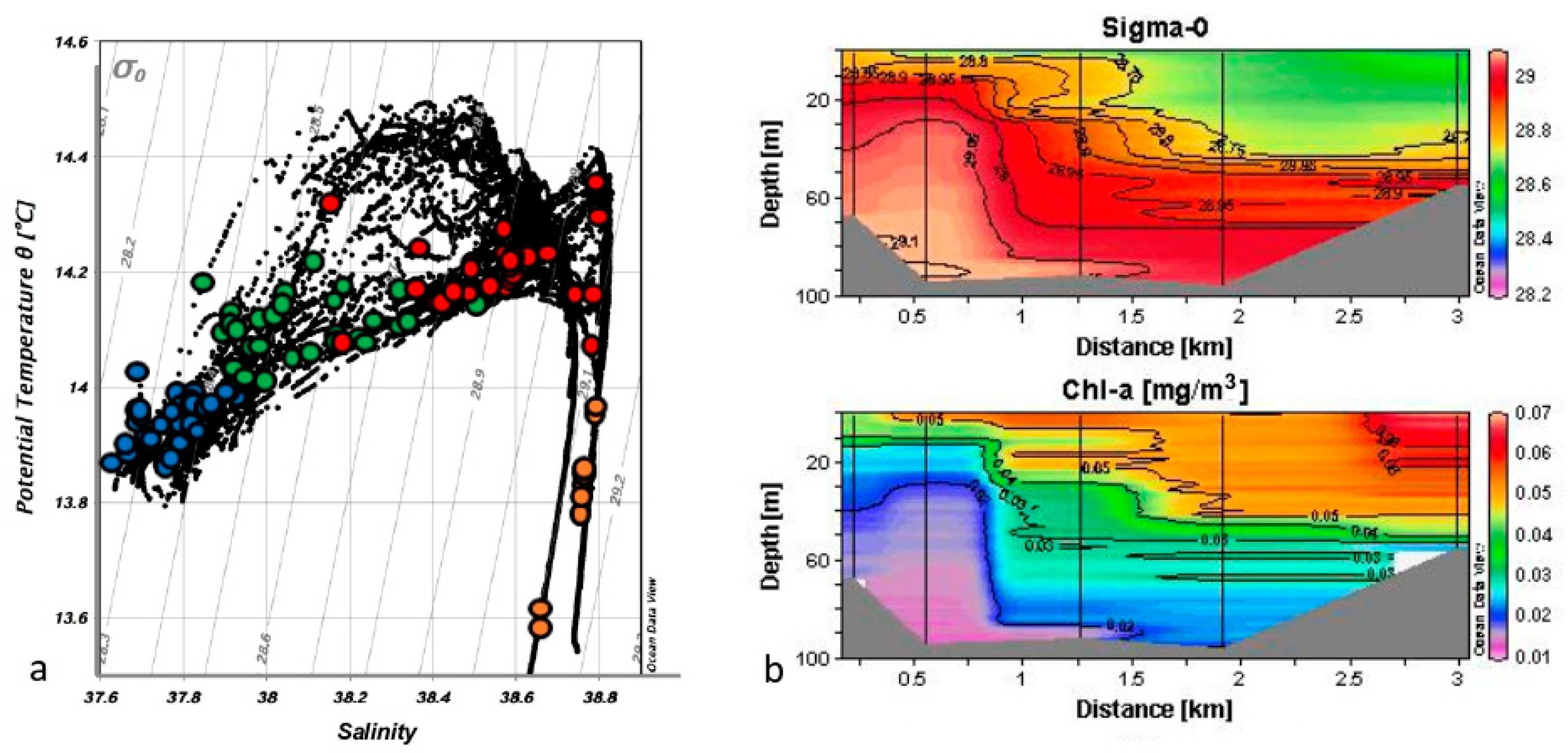
3.1. Physical–Chemical Variables
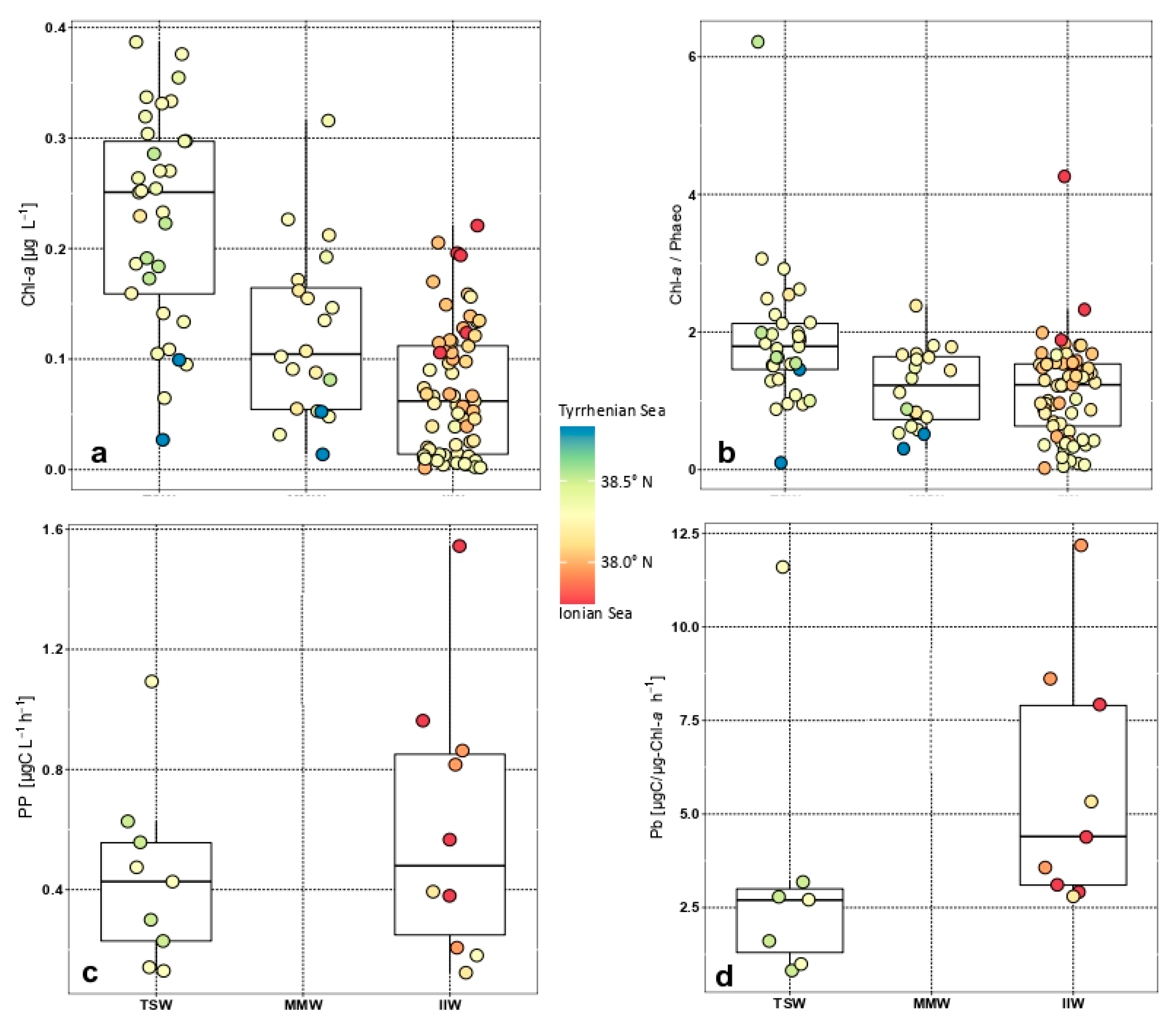
3.2. Distribution of Size-Fractionated Chlorophyll A, Phaeo-Pigments, and Primary Production
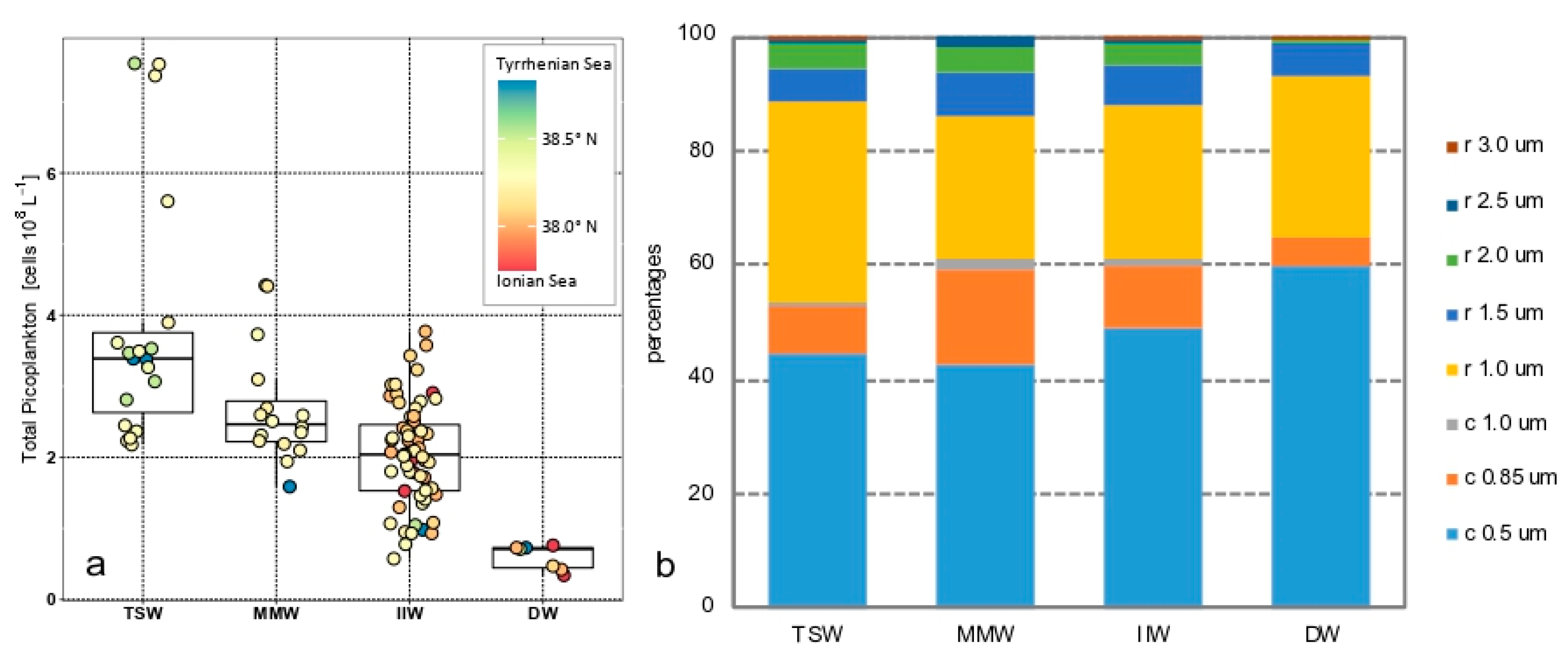
3.3. Total Picoplankton Community
3.4. Phytoplankton Communities
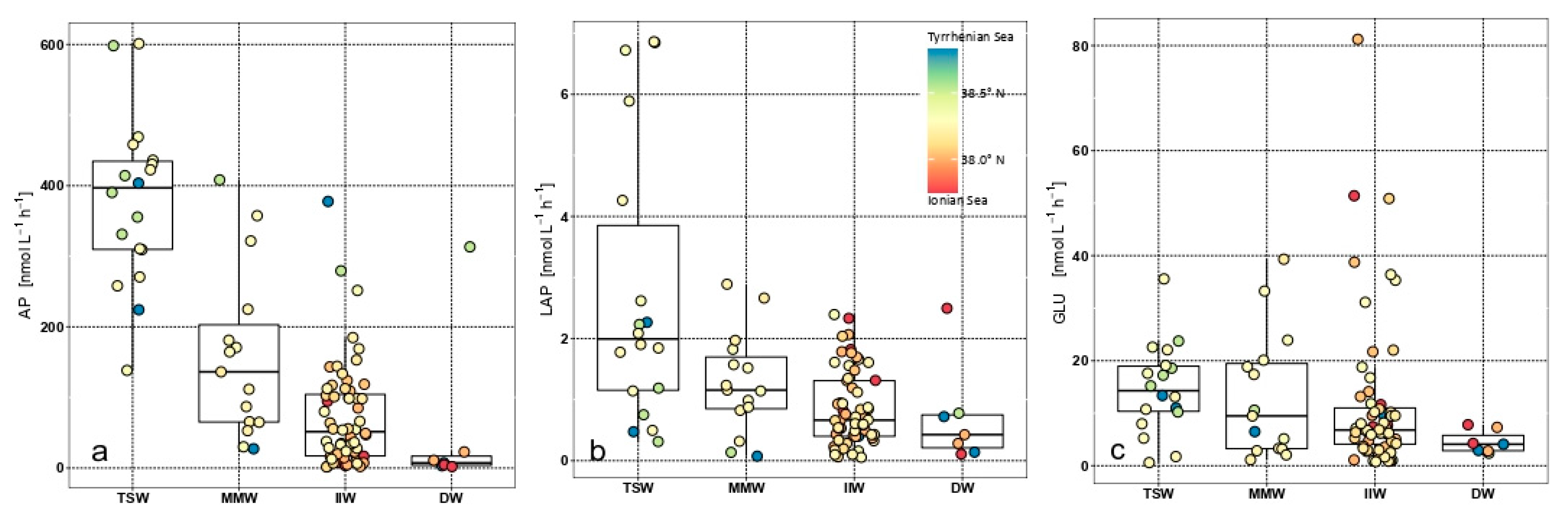
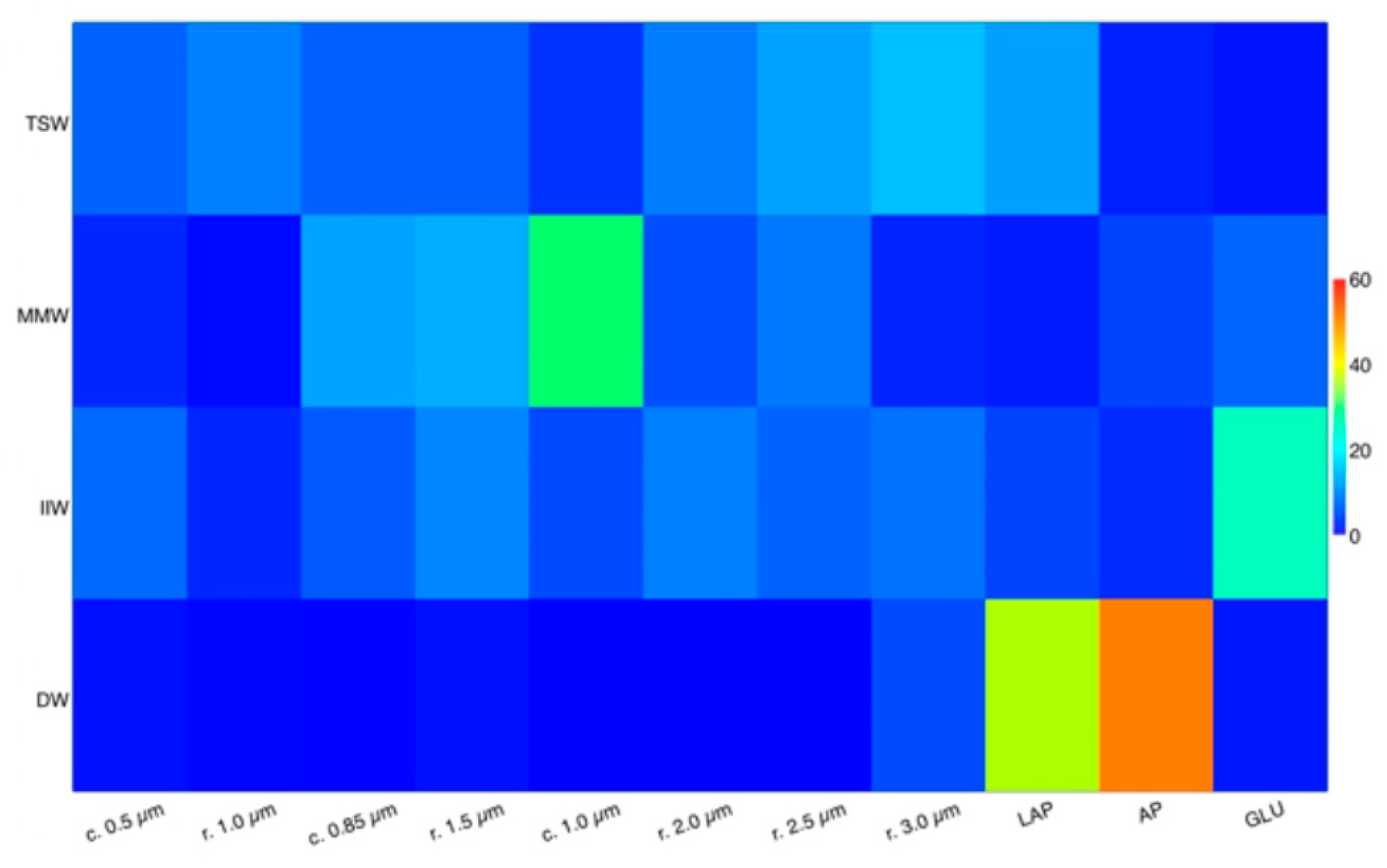
3.5. Functional Patterns
4. Discussion
4.1. Role of the Straits as a Biodiversity Mixer
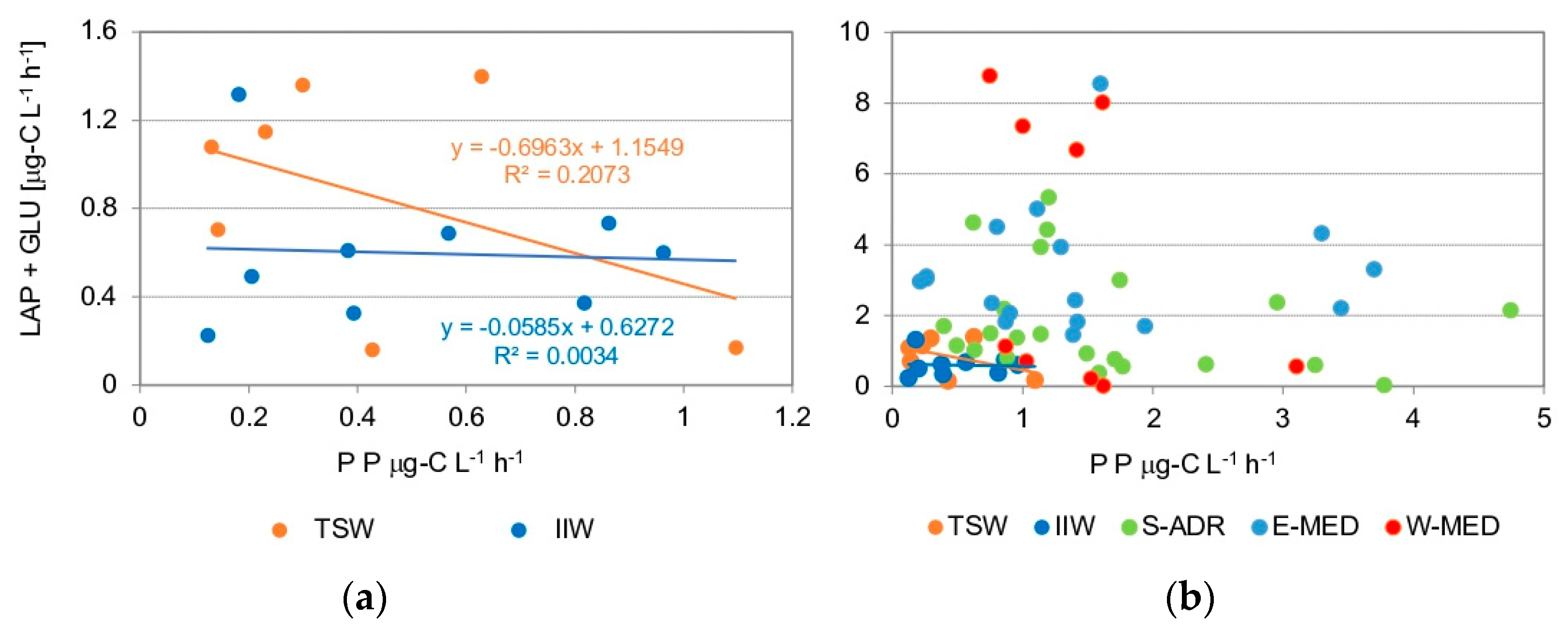
4.2. Organic Matter Photosynthetic Production and Microbial Decomposition Processes
4.3. Links between Autotrophic and Heterotrophic Microbial Processes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Magazzù, M.; Aubert, M.; Decembrini, F. The effect of tidal movements on planktonic transfer through the Straits of Messina. In The Strait of Messina Ecosystem; Guglielmo, L., Manganaro, A., De Domenico, E., Eds.; Università degli Studi: Messina, Italy, 1995; pp. 191–202. [Google Scholar]
- Magazzù, G.; Decembrini, F. Primary production, biomass and abundance of phototrophic picoplankton in the Mediterranean Sea: A review. Aquat. Microb. Ecol. 1995, 9, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Cescon, B.; Azzaro, F.; Creazzo, S.; Decembrini, F.; Magazzù, G. Processes affecting upwelling and primary productivity of the Straits of Messina. Boll. Geofisica Teor. Appl. 1997, 38, 137–147. [Google Scholar]
- Azzaro, F.; Decembrini, F.; Crisafi, E. Continuous survey of upwelling in the Straits of Messina. Rapp. Comm. Int. Mer Médit. 1995, 34, 167. [Google Scholar]
- Decembrini, F.; Azzaro, F.; Paschini, E.; Magazzù, G. Influence of upwelling phenomena on the phytoplankton production and biomass in the Straits of Messina. In Proceedings of the XII Congress of the Italian Society of Oceanology and Limnology, Isola di Vulcano, 18–21 September 1996, Italy; Picazzo, M., Ed.; Italian Society of Oceanology and Limnology: Genova, Italy, 1998; Volume II, pp. 159–171. Available online: http://www.aiol.info/wp-content/uploads/ATTI_12.2_CONGRESSO_AIOL_1_2.pdf (accessed on 26 March 2022).
- Decembrini, F.; Azzaro, F.; Leonardi, M. Short- and long-term variability of the hydrological characteristics in the coastal waters of a system under high hydrodynamism (Straits of Messina). In Proceedings of the XIII Congress of the Italian Society of Oceanology and Limnology, Portonovo, 28–30 September 1998, Ancona, Italy; Picazzo, M., Ed.; Italian Society of Oceanology and Limnology: Genova, Italy, 1999; Volume 3, pp. 151–158. Available online: http://www.aiol.info/wp-content/uploads/ATTI_13.1_CONGRESSO_AIOL.pdf (accessed on 26 March 2022).
- Azzaro, F.; Decembrini, F.; Raffa, F.; Crisafi, E. Seasonal variability of phytoplankton fluorescence in relation to the Straits of Messina (Sicily) tidal upwelling. Ocean Sci. 2007, 3, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Caruso, G.; Azzaro, F.; La Ferla, R.; De Pasquale, F.; Raffa, F.; Decembrini, F. Microbial enzymatic activities and prokaryotic abundance in the upwelling system of the Straits of Messina (Sicily): Distribution, dynamics and biogeochemical considerations. Adv. Oceanogr. Limnol. 2013, 4, 43–69. [Google Scholar] [CrossRef]
- Azam, F. Microbial control of oceanic carbon flux: The plot thickens. Science 1998, 280, 694–696. [Google Scholar] [CrossRef] [Green Version]
- Cotner, J.; Biddanda, B. Small Players, Large Role: Microbial Influence on Biogeochemical Processes in Pelagic Aquatic Ecosystems. Ecosystems 2002, 5, 105–121. [Google Scholar] [CrossRef]
- Liu, J.; Meng, Z.; Liu, X.; Zhang, X.-H. Microbial assembly, interaction, functioning, activity and diversification: A review derived from community compositional data. Mar. Life Sci. Technol. 2019, 1, 112–128. [Google Scholar] [CrossRef] [Green Version]
- Mapelli, F.; Varela, M.M.; Barbato, M.; Alvariño, R.; Fusi, M.; Álvarez, M.; Merlino, G.; Daffonchio, D.; Borin, S. Biogeography of planktonic bacterial communities across the whole Mediterranean Sea. Ocean Sci. 2013, 9, 585–595. [Google Scholar] [CrossRef] [Green Version]
- Caroppo, C.; Buttino, I.; Camatti, E.; Caruso, G.; De Angelis, R.; Facca, C.; Giovanardi, F.; Lazzara, L.; Mangoni, O.; Magaletti, E. State of the art and perspectives on the use of planktonic communities as indicators of environmental status in relation to the EU Marine Strategy Framework Directive. Biol. Mar. Mediterr. 2013, 20, 65–73. [Google Scholar] [CrossRef]
- Caruso, G.; La Ferla, R.; Azzaro, M.; Zoppini, A.; Marino, G.; Petochi, T.; Corinaldesi, C.; Leonardi, M.; Zaccone, R.; Fonda-Umani, S.; et al. Microbial assemblages for environmental quality assessment: Knowledge, gaps and usefulness in the European Marine Strategy Framework Directive. Crit. Rev. Microbiol. 2016, 42, 883–904. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Azzaro, M.; Caroppo, C.; Decembrini, F.; Monticelli, L.S.; Leonardi, M.; Maimone, G.; Zaccone, R.; La Ferla, R. Microbial community and its potential as descriptor of environmental status. ICES J. Mar. Sci. 2016, 73, 2174–2177. [Google Scholar] [CrossRef] [Green Version]
- Decembrini, F.; Caroppo, C.; Caruso, G.; Bergamasco, A. Linking Microbial Functioning and Trophic Pathways to Ecological Status in a Coastal Mediterranean Ecosystem. Water 2021, 13, 1325. [Google Scholar] [CrossRef]
- Hoppe, H.G.; Arnosti, C.; Herndl, G. Ecological significance of bacterial enzymes in the marine environment. In Enzymes in the Environment, Activity, Ecology and Applications; Burns, R.G., Dick, R.P., Eds.; Marcel Dekker: New York, NY, USA, 2002; pp. 73–107. [Google Scholar]
- Cunha, A.; Almeida, A.; Coelho, F.J.R.C.; Gomes, N.C.M.; Oliveira, V.; Santos, A.L. Bacterial extracellular enzymatic activity in globally changing aquatic ecosystems. In Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology; Microbiology Book Series No. 2; Méndez-Vila, A., Ed.; Formatex Research Center: Badajoz, Spain, 2010; pp. 124–135. [Google Scholar]
- Caruso, G.; Giacobbe, M.G.; Azzaro, F.; Decembrini, F.; Leonardi, M.; Miserocchi, S.; Cao, X.; Song, C.; Zhou, Y. All-In-One: Microbial Response to Natural and Anthropogenic Forcings in a Coastal Mediterranean Ecosystem, the Syracuse Bay (Ionian Sea, Italy). J. Mar. Sci. Eng. 2022, 10, 19. [Google Scholar] [CrossRef]
- Young, K.D. The selective value of bacterial shape. Microbiol. Mol. Biol. Rev. 2006, 70, 660–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cottrel, M.T.; Kirchman, D.L. Single-cell analysis of bacterial growth, cell size, and community structure in the Delaware estuary. Aquat. Microb. Ecol. 2004, 34, 139–149. [Google Scholar] [CrossRef] [Green Version]
- Pernthaler, J.; Amann, R. Fate of heterotrophic microbes in pelagic habitats: Focus on populations. Microbiol. Mol. Biol. Rev. 2005, 6, 440–461. [Google Scholar] [CrossRef] [Green Version]
- Racy, F.; Godinho, M.J.L.; Regali-Seleghim, M.H.; Bossolan, N.R.S.; Ferrari, A.C.; Lucca, J.V. Assessment of the applicability of morphological and size diversity indices to bacterial populations of reservoir in different trophic states. Acta Limnol. Bras. 2005, 17, 395–408. [Google Scholar]
- La Ferla, R.; Maimone, G.; Caruso, G.; Azzaro, F.; Azzaro, M.; Decembrini, F.; Cosenza, A.; Leonardi, M.; Paranhos, R. Are prokaryotic cell shape and size suitable to ecosystem characterization? Hydrobiologia 2014, 726, 65–80. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Katz, M.E.; Knoll, A.H.; Quigg, A.; Raven, J.A.; Schofield, O.; Taylor, F.J.R. The evolution of modern eukaryotic phytoplankton. Science 2004, 305, 354–360. [Google Scholar] [CrossRef] [Green Version]
- Finkel, Z.V.; Beardall, J.; Flynn, K.J.; Quigg, A.; Rees, T.A.V.; Raven, J.A. Phytoplankton in a changing world: Cell size and elemental stoichiometry. J. Plankton Res. 2010, 32, 119–137. [Google Scholar] [CrossRef] [Green Version]
- Decembrini, F.; Bergamasco, A.; Mangoni, O. Seasonal characteristics of size-fractionated phytoplankton community and fate of photosynthesized carbon in a sub-Antarctic area (Straits of Magellan). J. Mar. Syst. 2014, 136, 31–41. [Google Scholar] [CrossRef]
- de Vargas, C.; Audic, S.; Henry, N.; Decelle, J.; Mahé, F.; Logares, R.; Lara, E.; Berney, C.; Le Bescot, N.; Probert, I.; et al. Eukaryotic plankton diversity in the sunlit ocean. Science 2015, 348, 1261605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worm, B.; Barbier, E.B.; Beaumont, N.; Duffy, J.E.; Folke, C.; Halpern, B.S.; Jackson, J.B.C.; Lotze, H.K.; Micheli, F.; Palumbi, S.R.; et al. Impacts of biodiversity loss on ocean ecosystem services. Science 2006, 314, 787–790. [Google Scholar] [CrossRef] [Green Version]
- Naeem, S. Ecological consequences of declining biodiversity: A biodiversity-ecosystem function (BEF) framework for marine systems. In Marine Biodiversity and Ecosystem Functioning: Frameworks, Methodologies, and Integration; Solan, M., Aspden, R.J., Paterson, D.M., Eds.; Oxford University Press: Oxford, UK, 2012; pp. 34–51. [Google Scholar]
- Otero, J.; Álvarez-Salgado, X.A.; Bode, A. Phytoplankton Diversity Effect on Ecosystem Functioning in a Coastal Upwelling System. Front. Mar. Sci. 2020, 7, 592255. [Google Scholar] [CrossRef]
- Latasa, M.; Scharek, R.; Vidal, M.; Vila-Reixach, G.; Gutiérrez-Rodríguez, A.; Emelianov, M.; Gasol, J.M. Preferences of phytoplankton groups for waters of different trophic status in the northwestern Mediterranean Sea. Mar. Ecol. Progr. Ser. 2010, 407, 27–42. [Google Scholar] [CrossRef]
- Domingues, R.B.; Barbosa, A.; Galvao, H. Constraints on the Use of Phytoplankton as a Biological Quality Element within the Water Framework Directive in Portoguese Waters. Mar. Pollut. Bull. 2008, 56, 1389–1395. [Google Scholar] [CrossRef]
- Pachés, M.; Romero, I.; Hermosilla, Z.; Martinez-Guijarro, R. PHYMED: An Ecological Classification System for the Water Framework Directive Based on Phytoplankton Community Composition. Ecol. Indic. 2012, 19, 15–23. [Google Scholar] [CrossRef]
- Margalef, R. Turbulence and marine life. Sci. Mar. 1997, 61, 109–123. [Google Scholar]
- Largier, J.L. Upwelling bays: How coastal upwelling controls circulation, habitat, and productivity in bays. Ann. Rev. Mar. Sci. 2020, 12, 20.1–20.33. [Google Scholar] [CrossRef]
- Mayali, X. Editorial: Metabolic Interactions between Bacteria and Phytoplankton. Front. Microbiol. 2018, 9, 727. [Google Scholar] [CrossRef] [PubMed]
- Azam, F.; Malfatti, F. Microbial structuring of marine ecosystems. Nat. Rev. Microbiol. 2007, 5, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, T.S. Recent observations on the intermediate and deep-water circulation in the Southern Tyrrhenian Sea. Oceanol. Acta 1988, 9, 41–50. [Google Scholar]
- Bergamasco, A.; Crisafi, E.; Decembrini, F.; Messina, E.; Tamiro, E.; Giunta, G. A Laboratory for the Observation of a Highly-Energetic Coastal Marine System: The Straits of Messina. In Coastal and Marine Spatial Planning. Marine Research at CNR; Brugnoli, E., Cavarretta, G., Mazzola, S., Trincardi, F., Ravaioli, M., Santoleri, R., Eds.; National Research Council: Rome, Italy, 2011; Volume DTA/06-2011, pp. 2185–2202. Available online: https://dta.cnr.it/dta-06-2011-marine-research-at-cnr/ (accessed on 26 March 2022).
- Cucco, A.; Quattrocchi, G.; Olita, A.; Fazioli, L.; Ribotti, A.; Sinerchia, M.; Tedesco, C.; Sorgente, R. Hydrodynamic modelling of coastal seas: The role of tidal dynamics in the Messina Strait, Western Mediterranean Sea. Nat. Hazards Earth Syst. Sci. 2016, 16, 1553–1569. [Google Scholar] [CrossRef] [Green Version]
- Decembrini, F.; Hopkins, T.S.; Azzaro, F. Variability and sustenance of the deep-chlorophyll maximum over a narrow shelf, Augusta gulf (Sicily). Chem. Ecol. 2004, 20, S231–S247. [Google Scholar] [CrossRef]
- Battaglia, P.; Pagano, L.; Consoli, P.; Esposito, V.; Granata, A.; Guglielmo, L.; Pedà, C.; Romeo, T.; Zagami, G.; Vicchio, T.M.; et al. Consumption of mesopelagic prey in the strait of Messina, an upwelling area of the Central Mediterranean Sea: Feeding behaviour of the blue jack mackerel Trachurus picturatus (Bowdich, 1825). Deep-Sea Res. I Oceanogr. Res. Pap. 2020, 155, 103158. [Google Scholar] [CrossRef]
- Azzaro, F.; Cavalli, R.M.; Decembrini, F.; Pignatti, S.; Santella, C. Biochemical and dynamical characteristics of the Messina Straits water by means of hyperspectral data. Hyperspectral Remote Sens. Ocean. 2001, 4154, 240–249. [Google Scholar] [CrossRef]
- Aminot, A.; Chaussepied, M. Manuel des Analyses Chimiques en Milieu Marin; Centre National pour l’Exploitation des Océans: Brest, France, 1983; pp. 1–395. [Google Scholar]
- Strickland, J.D.H.; Parsons, T.R. A Practical Handbook of Seawater Analysis, 2nd ed.; Fisheries Research Board of Canada Bulletin 157; Minister of Supply and Services Canada: Ottawa, ON, Canada, 1972; pp. 1–310. [Google Scholar]
- Steeman-Nielsen, E. The use of radioactive carbon (14C) for measuring organic production in the sea. J. Cons. Int. Explor. Mer 1952, 18, 117–140. [Google Scholar] [CrossRef]
- Porter, K.G.; Feig, Y.S. The use of DAPI for identifying and counting aquatic microflora. Limnol. Oceanogr. 1980, 25, 943–948. [Google Scholar] [CrossRef]
- Edler, L.; Elbrächter, M. The Utermöhl method for quantitative phytoplankton analysis. In Microscopic and Molecular Methods for Quantitative Phytoplankton Analysis; Karlson, B., Cusack, C., Bresnan, E., Eds.; UNESCO IOC Manuals and Guides n. 55; UNESCO: Paris, France, 2010; pp. 13–20. [Google Scholar]
- Andersen, P.; Throndsen, J. Estimating cell numbers. In Manual on Harmful Marine Microalgae; Hallegraeff, G.M., Anderson, D.M., Cembella, A.D., Eds.; UNESCO: Paris, France, 2004; pp. 99–129. [Google Scholar]
- Zingone, A.; Totti, C.; Sarno, D.; Cabrini, M.; Caroppo, C.; Giacobbe, M.G.; Lugliè, A.; Nuccio, C.; Socal, G. Fitoplancton: Metodiche di analisi quali-quantitativa. In Metodologie di Studio del Plancton Marino; Socal, G., Buttino, I., Cabrini, M., Mangoni, O., Penna, A., Totti, C., Eds.; Manuali e Linee Guida ISPRA SIBM: Rome, Italy, 2010; pp. 213–237. ISBN 978-88-448-0427-5. [Google Scholar]
- Hoppe, H.-G. Use of fluorogenic model substrates for extracellular enzyme activity (EEA) measurement of bacteria. In Handbook of Methods in Aquatic Microbial Ecology; Kemp, P.F., Sherr, B.F., Sherr, E.B., Cole, J.J., Eds.; Lewis Publisher: Boca Raton, FL, USA, 1993; pp. 423–432. [Google Scholar]
- Hoppe, H.-G. Phosphatase activity in the sea. Hydrobiologia 2003, 493, 187–200. [Google Scholar] [CrossRef]
- Monticelli, L.S.; Caruso, G.; Decembrini, F.; Caroppo, C.; Fiesoletti, F. Role of prokaryotic biomasses and activities in Carbon and Phosphorus cycles at a coastal, thermohaline front and in offshore waters (Gulf of Manfredonia, Southern Adriatic Sea). Microb. Ecol. 2014, 67, 501–519. [Google Scholar] [CrossRef] [PubMed]
- Schlitzer, R. Ocean Data View. 2001. Available online: https://odv.awi.de/software/download/ (accessed on 26 March 2022).
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software package for education and data analysis. Paleontol. Electronica 2001, 4, 1–9. [Google Scholar]
- Bier, R.L.; Bernhardt, E.S.; Boot, C.M.; Graham, E.B.; Hall, E.K.; Lennon, J.T.; Nemergut, D.R.; Osborne, B.B.; Ruiz-González, C.; Schimel, J.P.; et al. Linking microbial community structure and microbial processes: An empirical and conceptual overview. FEMS Microbiol. Ecol. 2015, 91, fiv113. [Google Scholar] [CrossRef] [PubMed]
- Clarke, K.R.; Gorley, R.N.; Somerfield, P.J.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 3rd ed.; PRIMER-E Ltd.: Plymouth, UK, 2014. [Google Scholar]
- Margalef, R. Ecología; Omega: Barcelona, Spain, 1977; pp. 1–951. [Google Scholar]
- Ludwig, J.A.; Reynolds, J.F. Statistical Ecology: A Primer on Methods and Computing; Wiley: New York, NY, USA, 1988; pp. 85–106. [Google Scholar]
- Pielou, E.C. Ecological Diversity; John Wiley & Sons: New York, NY, USA, 1975. [Google Scholar]
- Clarke, K.R.; Warwick, R.M. A taxonomic distinctness index and its statistical properties. J. Appl. Ecol. 1998, 35, 523–531. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. The taxonomic distinctness measure of biodiversity: Weighting of step lengths between hierarchical levels. Mar. Ecol. Prog. Ser. 1999, 184, 21–29. [Google Scholar] [CrossRef]
- Siokou-Frangou, I.; Christaki, U.; Mazzocchi, M.G.; Montresor, M.; Ribera d’Alcalá, M.; Vaqué, D.; Zingone, A. Plankton in the open Mediterranean Sea: A review. Biogeosciences 2010, 7, 1543–1586. [Google Scholar] [CrossRef] [Green Version]
- Caroppo, C.; Turicchia, S.; Margheri, M.C. Phytoplankton assemblages in coastal waters of the Northern Ionian Sea (eastern Mediterranean), with special reference to cyanobacteria. J. Mar. Biol. Assoc. UK 2006, 86, 927–937. [Google Scholar] [CrossRef]
- Caroppo, C.; Musco, L.; Stabili, L. Planktonic Assemblages in a Coastal Mediterranean Area Subjected to Anthropogenic Pressure. J. Geogr. Nat. Disasters 2014, 4, 121. [Google Scholar] [CrossRef] [Green Version]
- Mena, C.; Reglero, P.; Hidalgo, M.; Sintes, E.; Santiago, R.; Martín, M.; Moyà, G.; Balbín, R. Phytoplankton Community Structure Is Driven by Stratification in the Oligotrophic Mediterranean Sea. Front. Microbiol. 2019, 10, 1698. [Google Scholar] [CrossRef] [Green Version]
- Vidussi, F.; Marty, J.C.; Chiavérini, J. Phytoplankton pigment variations during the transition from spring bloom to oligotrophy in the northwestern Mediterranean sea. Deep-Sea Res. I Oceanogr. Res. Pap. 2000, 47, 423–445. [Google Scholar] [CrossRef]
- Marty, J.C.; Chiavérini, J.; Pizay, M.D.; Avril, B. Seasonal and interannual dynamics of nutrients and phytoplankton pigments in the western Mediterranean Sea at the DYFAMED time-series station (1991–1999). Deep-Sea Res. II Top. Stud. Oceanogr. 2002, 49, 1965–1985. [Google Scholar] [CrossRef]
- Brunet, C.; Casotti, R.; Vantrepotte, V.; Corato, F.; Conversano, F. Picophytoplankton diversity and photoacclimation in the Strait of Sicily (Mediterranean Sea) in summer. I. Mesoscale variations. Aquat. Microb. Ecol. 2006, 44, 127–141. [Google Scholar] [CrossRef]
- Casotti, R.; Landolfi, A.; Brunet, C.; D’Ortenzio, F.; Mangoni, O.; Ribera d’Alcalà, M. Composition and dynamics of the phytoplankton of the Ionian Sea (Eastern Mediterranean). J. Geophy. Res. 2003, 108, 1–17. [Google Scholar] [CrossRef]
- Decembrini, F.; Caroppo, C.; Azzaro, M. Size structure and production of phytoplankton community and carbon pathways channelling in the southern Tyrrhenian sea (Western Mediterranean). Deep-Sea Res. II Top. Stud. Oceanogr. 2009, 56, 687–699. [Google Scholar] [CrossRef]
- Karuza, A.; Caroppo, C.; Camatti, E.; Di Poi, E.; Monti, M.; Stabili, L.; Auriemma, R.; Pansera, M.; Cibic, T.; Del Negro, P. ‘End to end’ planktonic trophic web and its implications for the mussel farms in the Mar Piccolo of Taranto (Ionian Sea, Italy). Environ. Sci. Pollut. Res. 2016, 23, 12707–12724. [Google Scholar] [CrossRef] [PubMed]
- Varkitzi, I.; Psarra, S.; Assimakopoulou, G.; Pavlidou, A.; Krasakopoulou, E.; Velaoras, D.; Papathanassiou, E.; Pagou, K. Phytoplankton dynamics and bloom formation in the oligotrophic Eastern Mediterranean: Field studies in the Aegean, Levantine and Ionian seas. Deep-Sea Res. II Top. Stud. Oceanogr. 2020, 171, 104662. [Google Scholar] [CrossRef]
- Bonomo, S.; Placenti, F.; Quinci, E.M.; Cuttitta, A.; Genovese, S.; Mazzola, S.; Bonanno, A. Living coccolithophores community from Southern Tyrrhenian Sea (Central Mediterranean—Summer 2009). Mar. Micropaleontol. 2017, 131, 10–24. [Google Scholar] [CrossRef]
- Marañón, E. Cell size as a key determinant of phytoplankton metabolism and community structure. Ann. Rev. Mar. Sci. 2015, 7, 241–264. [Google Scholar] [CrossRef] [Green Version]
- Bode, A.; Álvarez-Ossorio, M.T.; Barquero, S.; Lorenzo, J.; Louro, A.; Varela, M. Seasonal variations in upwelling and in the grazing impact of copepods on phytoplankton off A Coruña (Galicia, NW Spain). J. Exp. Mar. Biol. Ecol. 2003, 297, 85–105. [Google Scholar] [CrossRef]
- Smayda, T.S. Turbulence, watermass stratification and harmful algal blooms: An alternative view and frontal zones as “pelagic seed banks”. Harmful Algae 2002, 1, 95–112. [Google Scholar] [CrossRef]
- Martellucci, R.; Salon, S.; Cossarini, G.; Piermattei, V.; Marcelli, M. Coastal phytoplankton bloom dynamics in the Tyrrhenian Sea: Advantage of integrating in situ observations, large-scale analysis and forecast systems. J. Mar. Syst. 2021, 218, 103528. [Google Scholar] [CrossRef]
- Zingone, A.; D’Alelio, D.; Mazzocchi, M.G.; Montresor, M.; Sarno, D.; LTER-MC team. Time series and beyond: Multifaceted plankton research at a marine Mediterranean LTER site. Nat. Conserv. 2019, 34, 273–310. [Google Scholar] [CrossRef] [Green Version]
- Margalef, R. The food web in the pelagic environment. Helgol. Wiss. Meer. 1967, 15, 548–559. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, C.N.; Morri, C. Marine biodiversity of the Mediterranean Sea: Situation, problems and prospects for future research. Mar. Pollut. Bull. 2000, 40, 367–376. [Google Scholar] [CrossRef]
- Antacli, J.C.; Silva, R.I.; Jaureguizar, A.J.; Hernández, D.R.; Mendiolar, M.; Sabatini, M.E.; Akselman, R. Phytoplankton and protozooplankton on the southern Patagonian shelf (Argentina, 47°–55° S) in late summer: Potentially toxic species and community assemblage structure linked to environmental features. J. Sea Res. 2018, 140, 63–80. [Google Scholar] [CrossRef]
- Robarts, R.D.; Zohary, T.; Waiser, M.J.; Yacobi, Y.Z. Bacterial abundance, biomass, and production in relation to phytoplankton biomass in the Levantine Basin of the southeastern Mediterranean Sea. Mar. Ecol. Prog. Ser. 1996, 137, 273–281. [Google Scholar] [CrossRef]
- Acinas, S.G.; Antón, J.; Rodgríguez-Valera, F. Diversity of free-living and attached bacteria in offshore western Mediterranean waters as depicted by analysis of genes encoding 16S rRNA. Appl. Environ. Microbiol. 1999, 65, 514–522. [Google Scholar] [CrossRef] [Green Version]
- La Ferla, R.; Lo Giudice, A.; Maimone, G. Morphology and LPS content for the estimation of marine bacterioplankton biomass in the Ionian Sea. Sci. Mar. 2004, 68, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Jochem, F.J. Morphology and DNA content of bacterioplankton in the northern Gulf of Mexico: Analysis by epifluorescence microscopy and flow cytometry. Aquat. Microb. Ecol. 2001, 25, 179–194. [Google Scholar] [CrossRef]
- Coelho-Souza, S.A.; Araújo, F.V.; Cury, J.C.; Jesus, H.E.; Pereira, G.C.; Guimarães, J.R.D.; Peixoto, R.S.; Dávila, A.M.R.; Rosado, A.S. Bacterial and Archaeal Communities Variability Associated with Upwelling and Anthropogenic Pressures in the Protection Area of Arraial do Cabo (Cabo Frio region-RJ). An. Acad. Bras. Ciênc. 2015, 87, 1737–1750. [Google Scholar] [CrossRef]
- La Ferla, R.; Leonardi, M. Ecological implications of biomass and morphotype variations of bacterioplankton: An example in a coastal zone of the Northern Adriatic Sea (Mediterranean). Mar. Ecol. 2005, 26, 82–88. [Google Scholar] [CrossRef]
- Gasol, J.M.; Del Giorgio, P.A.; Massana, R.; Duarte, C.M. Active versus inactive bacteria: Size-dependence in a coastal marine plankton community. Mar. Ecol. Prog. Ser. 1995, 128, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Tamburini, C.; Garel, M.; Ali, B.A.; Mérigot, B.; Kriwy, P. Distribution and activity of Bacteria and Archea in the different water masses of the Tyrrhenian Sea. Deep-Sea Res. II Top. Stud. Oceanogr. 2009, 56, 700–712. [Google Scholar] [CrossRef]
- La Ferla, R.; Azzaro, M.; Caruso, G.; Monticelli, L.S.; Maimone, G.; Zaccone, R.; Packard, T.T. Prokaryotic abundance and heterotrophic metabolism in the deep Mediterranean Sea. Adv. Oceanogr. Limnol. 2010, 1, 143–166. [Google Scholar] [CrossRef]
- Placenti, F.; Azzaro, M.; Artale, V.; La Ferla, R.; Caruso, G.; Santinelli, C.; Maimone, G.; Monticelli, L.; Quinci, E.M.; Sprovieri, M. Biogeochemical patterns and microbial processes in the Eastern Mediterranean Deep Water of Ionian Sea. Hydrobiologia 2018, 815, 97–112. [Google Scholar] [CrossRef]
- Zaccone, R.; Caruso, G.; Azzaro, M.; Azzaro, F.; Crisafi, E.; Decembrini, F.; De Domenico, E.; De Domenico, M.; La Ferla, R.; Leonardi, M.; et al. Prokaryotic activities and abundance in pelagic areas of the Ionian Sea. Chem. Ecol. 2010, 26 (Suppl. S1), 169–197. [Google Scholar] [CrossRef]
- Azzaro, M.; La Ferla, R.; Maimone, G.; Monticelli, L.S.; Zaccone, R.; Civitarese, G. Prokaryotic dynamics and heterotrophic metabolism in a deep convection site of Eastern Mediterranean Sea (the Southern Adriatic Pit). Cont. Shelf Res. 2012, 44, 106–118. [Google Scholar] [CrossRef]
- Zaccone, R.; Caruso, G. Microbial enzymes in the Mediterranean Sea: Relationship with climate changes. AIMS Microbiol. 2019, 5, 251–271. [Google Scholar] [CrossRef]
- Luna, G.M.; Bianchelli, S.; Decembrini, F.; De Domenico, E.; Danovaro, R.; Dell’Anno, A. The dark portion of the Mediterranean Sea is a bioreactor of organic matter cycling. Glob. Biogeochem. Cycles 2012, 26, GB2017. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TSW | MMW | IIW | DW | |||||||||||
| Parameter * | Fraction/Species | Unit | Avg | SD | n | Avg | SD | n | Avg | SD | n | Avg | SD | n |
| T | °C | 13.90 | 0.05 | 35 | 14.09 | 0.06 | 23 | 14.25 | 0.11 | 113 | 13.72 | 0.19 | 10 | |
| S | 37.78 | 0.08 | 35 | 38.06 | 0.15 | 23 | 38.61 | 0.16 | 113 | 38.70 | 0.10 | 10 | ||
| O2 | mL L−1 | 5.50 | 0.05 | 35 | 5.24 | 0.11 | 23 | 4.85 | 0.26 | 113 | 4.36 | 0.10 | 10 | |
| Fluorescence | µg L−1 | 0.075 | 0.017 | 35 | 0.050 | 0.020 | 23 | 0.030 | 0.016 | 113 | 0.013 | 0.001 | 10 | |
| PO4 | Concentration | µM | 0.206 | 0.071 | 34 | 0.263 | 0.101 | 23 | 0.255 | 0.114 | 104 | 0.294 | 0.060 | 10 |
| NH4 | 0.907 | 0.648 | 34 | 0.938 | 0.535 | 23 | 1.030 | 0.590 | 104 | 1.027 | 0.570 | 10 | ||
| NO2 + NO3 | 1.160 | 0.440 | 34 | 1.600 | 0.420 | 23 | 2.244 | 1.116 | 104 | 8.408 | 3.475 | 10 | ||
| Si(OH)4 | 2.077 | 1.470 | 34 | 2.518 | 1.536 | 23 | 2.856 | 1.139 | 104 | 6.314 | 1.622 | 10 | ||
| chla | Total | µg L−1 | 0.228 | 0.095 | 33 | 0.122 | 0.076 | 20 | 0.065 | 0.059 | 76 | |||
| >10 µm | 0.013 | 0.006 | 12 | 0.007 | 0.002 | 6 | 0.010 | 0.012 | 10 | |||||
| 2–10 µm | 0.034 | 0.017 | 12 | 0.020 | 0.013 | 6 | 0.028 | 0.019 | 10 | |||||
| 0.5–2 µm | 0.098 | 0.054 | 12 | 0.056 | 0.046 | 6 | 0.071 | 0.055 | 10 | |||||
| phaeo | Total | µg L−1 | 0.137 | 0.065 | 33 | 0.102 | 0.049 | 20 | 0.052 | 0.029 | 76 | |||
| >10 µm | 0.031 | 0.014 | 12 | 0.052 | 0.057 | 6 | 0.028 | 0.017 | 10 | |||||
| 2–10 µm | 0.040 | 0.036 | 12 | 0.019 | 0.014 | 6 | 0.013 | 0.008 | 10 | |||||
| 0.5–2 µm | 0.035 | 0.022 | 12 | 0.030 | 0.010 | 6 | 0.030 | 0.010 | 10 | |||||
| PP | Total | µgC L−1 h−1 | 0.442 | 0.302 | 9 | 1.069 | 1 | 0.603 | 0.446 | 10 | ||||
| >10 µm | 0.028 | 0.019 | 9 | 0.054 | 1 | 0.026 | 0.012 | 10 | ||||||
| 2–10 µm | 0.073 | 0.094 | 9 | 0.302 | 1 | 0.134 | 0.134 | 10 | ||||||
| 0.5–2 µm | 0.341 | 0.206 | 9 | 0.713 | 1 | 0.443 | 0.308 | 10 | ||||||
| LAP | nmol L−1 h−1 | 2.759 | 2.305 | 18 | 1.279 | 0.828 | 15 | 0.847 | 0.619 | 60 | 0.707 | 0.831 | 7 | |
| AP | 378.99 | 119.36 | 18 | 160.26 | 120.62 | 15 | 72.357 | 73.227 | 60 | 52.029 | 115.42 | 7 | ||
| GLU | 14.760 | 8.533 | 18 | 13.134 | 11.957 | 15 | 11.726 | 14.620 | 60 | 4.507 | 2.197 | 7 | ||
| PPA | Abundance | cells × 106 L−1 | 19.516 | 3.619 | 18 | 13.213 | 5.943 | 12 | 7.812 | 3.913 | 44 | |||
| PPT | cells × 108 L−1 | 3.867 | 1.788 | 19 | 2.699 | 0.824 | 16 | 2.051 | 0.710 | 64 | 0.594 | 0.178 | 7 | |
| Cocci 0.5 µm | 1.806 | 0.895 | 10 | 1.417 | 0.445 | 6 | 0.994 | 0.452 | 29 | 0.343 | 0.117 | 4 | ||
| Cocci 0.85 µm | 0.324 | 0.308 | 10 | 0.543 | 0.306 | 6 | 0.223 | 0.143 | 29 | 0.026 | 0.023 | 4 | ||
| Cocci 1.0 µm | 0.023 | 0.044 | 10 | 0.071 | 0.090 | 6 | 0.015 | 0.023 | 29 | 0.000 | 0.000 | 4 | ||
| PPT | Rods 1.0 µm | cells × 108 L−1 | 1.411 | 0.877 | 10 | 0.830 | 0.251 | 6 | 0.545 | 0.236 | 29 | 0.163 | 0.070 | 4 |
| Rods 1.5 µm | 0.245 | 0.194 | 10 | 0.243 | 0.198 | 6 | 0.143 | 0.103 | 29 | 0.031 | 0.024 | 4 | ||
| Rods 2.0 µm | 0.165 | 0.160 | 10 | 0.160 | 0.098 | 6 | 0.081 | 0.075 | 29 | 0.004 | 0.007 | 4 | ||
| Rods 2.5 µm | 0.048 | 0.107 | 10 | 0.049 | 0.071 | 6 | 0.017 | 0.039 | 29 | 0.000 | 0.000 | 4 | ||
| Rods 3.0 µm | 0.013 | 0.042 | 10 | 0.008 | 0.015 | 6 | 0.006 | 0.015 | 29 | 0.004 | 0.007 | 4 | ||
| Larger Phytoplankton | Total | 69.102 | 40.590 | 18 | 39.580 | 18.002 | 13 | 44.783 | 21.547 | 42 | ||||
| Diatoms | 6.585 | 5.414 | 18 | 4.825 | 2.982 | 13 | 3.595 | 2.770 | 42 | |||||
| Dinoflagellates | cells × 103 L−1 | 3.424 | 1.826 | 18 | 1.992 | 1.425 | 13 | 2.214 | 1.798 | 42 | ||||
| Coccolithophorids | 4.599 | 3.683 | 18 | 2.160 | 2.108 | 13 | 3.568 | 4.756 | 42 | |||||
| Others | 54.495 | 35.758 | 18 | 30.603 | 15.792 | 13 | 35.406 | 16.480 | 42 | |||||
| TSW | MMW | IIW | |
|---|---|---|---|
| S | 13.89 ± 3.92 [6, 20] | 12.23 ± 4.07 [4, 18] | 8.83 ± 1.98 [6, 12] |
| d | 2.73 ± 0.71 [1.26, 3.78] | 2.44 ± 0.77 [0.82, 3.49] | 2.17 ± 0.61 [1.05, 3.65] |
| J’ | 0.98 ± 0.01 [0.95, 0.98] | 0.97 ± 0.02 [0.93, 0.98] | 0.98 ± 0.01 [0.96, 0.99] |
| H’ | 2.53 ± 0.35 [1.71, 2.95] | 2.38 ± 0.43 [1.29, 2.84] | 2.27 ± 0.31 [1.55, 2.90] |
| Δ | 60.54 ± 2.67 [56.21, 65.95] | 60.80 ± 2.77 [56.25, 65.65] | 62.16 ± 1.88 [58.09, 65.69] |
| Δ* | 66.18 ± 5.65 [59.12, 80.01] | 68.03 ± 7.71 [60.23, 87.72] | 69.59 ± 4.49 [61.43, 80.17] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caroppo, C.; Azzaro, F.; Bergamasco, A.; Caruso, G.; Decembrini, F. Phytoplankton and Bacterial Communities’ Patterns in a Highly Dynamic Ecosystem (Central Mediterranean Sea). Water 2022, 14, 2057. https://doi.org/10.3390/w14132057
Caroppo C, Azzaro F, Bergamasco A, Caruso G, Decembrini F. Phytoplankton and Bacterial Communities’ Patterns in a Highly Dynamic Ecosystem (Central Mediterranean Sea). Water. 2022; 14(13):2057. https://doi.org/10.3390/w14132057
Chicago/Turabian StyleCaroppo, Carmela, Filippo Azzaro, Alessandro Bergamasco, Gabriella Caruso, and Franco Decembrini. 2022. "Phytoplankton and Bacterial Communities’ Patterns in a Highly Dynamic Ecosystem (Central Mediterranean Sea)" Water 14, no. 13: 2057. https://doi.org/10.3390/w14132057
APA StyleCaroppo, C., Azzaro, F., Bergamasco, A., Caruso, G., & Decembrini, F. (2022). Phytoplankton and Bacterial Communities’ Patterns in a Highly Dynamic Ecosystem (Central Mediterranean Sea). Water, 14(13), 2057. https://doi.org/10.3390/w14132057