
Recovery of Macrobenthic Food Web on Rocky Shores Following the Hebei Spirit Oil Spill as Revealed by C and N Stable Isotopes



Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Sample Collection and Processing
2.3. Stable Isotope Analysis
2.4. Data Analysis
3. Results
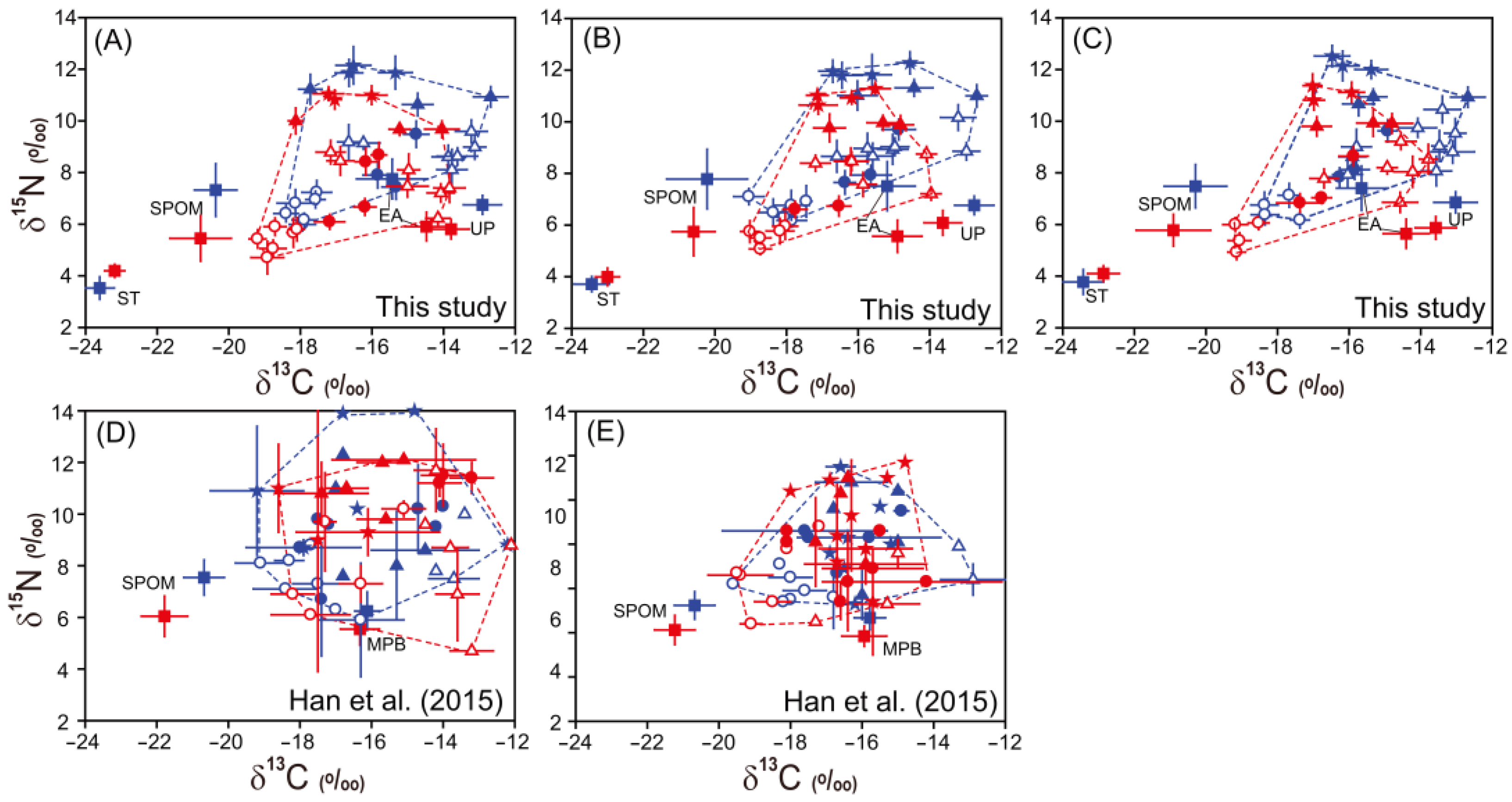
3.1. Stable Isotope Ratios of Potential Food Sources
3.2. Stable Isotope Ratios of Consumer Species
3.3. Isotopic Niche Areas
4. Discussion
4.1. Comparison of Isotopic Signatures of Organic Matter Sources and Microbenthic Consumers between Oil-Impacted and Control Sites
4.2. Isotopic Niche Comparison of Microbenthic Consumers between Oil-Impacted and Control Sites
4.3. Ecological Implication of Trophic Recovery after the HSOS
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kennish, M.J. Environmental threats and environmental future of estuaries. Environ. Conserv. 2002, 29, 78–107. [Google Scholar] [CrossRef]
- Mendelssohn, I.A.; Andersen, G.L.; Baltz, D.M.; Caffey, R.H.; Carman, K.R.; Flegger, J.W.; Joye, S.B.; Lin, Q.; Maltby, E.; Overton, E.B.; et al. Oil impacts on coastal wetlands: Implications for the Mississippi River delta ecosystem after the Deepwater Horizon oil spill. BioScience 2012, 62, 562–574. [Google Scholar] [CrossRef]
- Depellegrin, D.; Blažauskas, N. Integrating ecosystem service values into oil spill impact assessment. J. Coast. Res. 2013, 29, 836–846. [Google Scholar] [CrossRef]
- Suchanek, T.H. Oil impacts on marine invertebrate populations and communities. Am. Zool. 1993, 33, 510–523. [Google Scholar] [CrossRef] [Green Version]
- Peterson, C.H.; Rice, S.D.; Short, J.W.; Esler, D.; Bodkin, J.L.; Ballachey, B.E.; Irons, D.B. Long-term ecosystem response to the Exxon Valdez oil spill. Science 2003, 302, 2082–2086. [Google Scholar] [CrossRef] [Green Version]
- Moreno, R.; Jover, L.; Diez, C.; Sardà, F.; Sanpera, C. Ten years after the Prestige oil spill: Seabird trophic ecology as indicator of long-term effects on the coastal marine ecosystem. PLoS ONE 2013, 8, e77360. [Google Scholar] [CrossRef]
- Carman, K.R.; Fleeger, J.W.; Pomarico, S.M. Response of a benthic food web to hydrocarbon contamination. Limnol. Oceanogr. 1997, 42, 561–571. [Google Scholar] [CrossRef]
- McCann, M.J.; Able, K.W.; Christian, R.R.; Fodrie, F.J.; Jensen, O.P.; Johnson, J.J.; López-Duarte, P.C.; Martin, C.W.; Olin, J.A.; Polito, M.J.; et al. Key taxa in food web responses to stressors: The Deepwater Horizon oil spill. Front. Ecol. Environ. 2017, 15, 142–149. [Google Scholar] [CrossRef]
- Halpern, B.S.; Selkoe, K.A.; Micheli, F.; Kappel, C.V. Evaluating and ranking the vulnerability of global marine ecosystems to anthropogenic threats. Conserv. Biol. 2007, 21, 1301–1315. [Google Scholar] [CrossRef]
- Hawkins, S.J.; Evans, A.J.; Mieszkowska, N.; Adams, L.C.; Bray, S.; Burrow, M.T.; Firth, L.B.; Genner, M.J.; Leung, K.M.Y.; Moore, P.J.; et al. Distinguishing globally-driven changes from regional- and local-scale impacts: The case for long-term and broad-scale studies of recovery from pollution. Mar. Pollut. Bull. 2017, 124, 573–586. [Google Scholar] [CrossRef] [Green Version]
- Yim, U.H.; Hong, S.; Lee, C.; Kim, M.; Jung, J.H.; Ha, S.Y.; An, J.G.; Kwon, B.O.; Kim, T.; Lee, C.H.; et al. Rapid recovery of coastal environment and ecosystem to the Hebei Spirit oil spill’s impact. Environ. Int. 2020, 136, 105438. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Khim, J.S.; Ryu, J.; Kang, S.G.; Shim, W.J.; Yim, U.H. Environmental and ecological effects and recoveries after five years of the Hebei Spirit oil spill, Taean, Korea. Ocean Coast. Manag. 2014, 102, 522–532. [Google Scholar] [CrossRef]
- Kim, M.; Jung, J.H.; Ha, S.Y.; An, J.G.; Shim, W.J.; Yim, U.H. Long-term monitoring of PAH contamination in sediment and recovery after the Hebei Spirit oil spill. Arch. Environ. Con. Tox. 2017, 73, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-H.; Lee, J.H.; Sung, C.G.; Moon, S.D.; Kang, S.K.; Lee, J.H.; Yim, U.H.; Shim, W.J.; Ha, S.Y. Monitoring toxicity of polycyclic aromatic hydrocarbons in intertidal sediments for five years after the Hebei Spirit oil spill in Taean, Republic of Korea. Mar. Pollut. Bull. 2013, 76, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Yu, O.H.; Lee, H.G.; Shim, W.J.; Kim, M.; Park, H.S. Initial impacts of the Hebei Spirit oil spill on the sandy beach macrobenthic community west coast of Korea. Mar. Pollut. Bull. 2013, 70, 189–196. [Google Scholar] [CrossRef]
- Jung, Y.H.; Yoon, K.T.; Shim, W.J.; Park, H.S. Short-term variation of the macrobenthic fauna structure on rocky shores after the Hebei Spirit oil spill, west coast of Korea. J. Coast. Res. 2014, 31, 177–183. [Google Scholar] [CrossRef]
- Han, E.; Park, H.J.; Bergamino, L.; Choi, K.S.; Choy, E.J.; Yu, O.H.; Lee, T.W.; Park, H.S.; Shim, W.J.; Kang, C.K. Stable isotope analysis of a newly established macrofaunal food web 1.5 years after the Hebei Spirit oil spill. Mar. Pollut. Bull. 2015, 90, 167–180. [Google Scholar] [CrossRef]
- Layman, C.A.; Araujo, M.S.; Boucek, R.; Hammerschlag-Peyer, C.M.; Harrison, E.; Jud, Z.R.; Matich, P.; Rosenblatt, A.E.; Vaudo, J.J.; Yeager, L.A.; et al. Applying stable isotopes to examine food-web structure: An overview of analytical tools. Biol. Rev. 2012, 87, 545–562. [Google Scholar] [CrossRef]
- Mancinelli, G.; Vizzini, S. Assessing anthropogenic pressures on coastal marine ecosystems using stable CNS isotopes: State of the art, knowledge gaps, and community scale perspectives. Estuar. Coast. Shelf Sci. 2015, 156, 195–204. [Google Scholar] [CrossRef]
- Fry, B.; Sherr, E. δ13C measurements as indicators of carbon flow in marine and freshwater ecosystems. Contrib. Mar. Sci. 1984, 27, 49–63. [Google Scholar]
- Post, D.M. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- McCutchan, J.H., Jr.; Lewis, W.M., Jr.; Kendall, C.; McGrath, C.C. Variation in trophic shift for stable isotope ratios of carbon, nitrogen, and sulfur. Oikos 2003, 102, 378–390. [Google Scholar] [CrossRef]
- Vizzini, S.; Mazzola, A. The effects of anthropogenic organic matter inputs on stable carbon and nitrogen isotopes in organisms from different trophic levels in a southern Mediterranean coastal area. Sci. Total Environ. 2006, 368, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Quillien, N.; Nordström, M.C.; Schaal, G.; Bonsdorff, E.; Grall, J. Opportunistic basal resource simplifies food web structure and functioning of a highly dynamic marine environment. J. Exp. Mar. Biol. Ecol. 2016, 477, 92–102. [Google Scholar] [CrossRef]
- Park, H.J.; Kwak, J.H.; Lee, Y.J.; Kang, H.Y.; Choy, E.J.; Kang, C.K. Trophic structures of two contrasting estuarine ecosystems with and without a dike on the temperate coast of Korea as determined by stable isotopes. Estuar. Coasts 2020, 43, 560–577. [Google Scholar] [CrossRef]
- Kim, M.; Hong, S.H.; Won, J.; Yim, U.H.; Jung, J.H.; Ha, S.Y.; An, J.G.; Joo, C.; Kim, E.; Han, G.M.; et al. Petroleum hydrocarbon contaminations in the intertidal seawater after the Hebei Spirit oil spill–Effect of tidal cycle on the TPH concentrations and the chromatographic characterization of seawater extracts. Water Res. 2013, 47, 758–768. [Google Scholar] [CrossRef]
- Jung, Y.H.; Park, H.S.; Yoon, K.T.; Kim, H.J.; Shim, W.J. Long-term changes in rocky intertidal macrobenthos during the five years after the Hebei Spirit oil spill, Taean, Korea. Ocean Sci. J. 2017, 52, 103–112. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA + for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Jackson, A.L.; Inger, R.; Parnell, A.C.; Bearhop, S. Comparing isotopic niche widths among and within communities: SIBER—Stable Isotope Bayesian Ellipses in R. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Layman, C.A.; Arrington, D.A.; Montaña, C.G.; Post, D.M. Can stable isotope ratios provide for community-wide measures of trophic structure? Ecology 2007, 88, 42–48. [Google Scholar] [CrossRef]
- Swanson, H.K.; Lysy, M.; Power, M.; Stasko, A.D.; Johnson, J.D.; Reist, J.D. A new probabilistic method for quantifying n-dimensional ecological niches and niche overlap. Ecology 2015, 96, 318–324. [Google Scholar] [CrossRef]
- Peterson, C.T.; Grubbs, R.D.; Mickle, A. An investigation of effects of the Deepwater Horizon oil spill on coastal fishes in the Florida big bend using fisher-independent surveys and stable isotope analysis. Southeast. Nat. 2017, 16, G93–G108. [Google Scholar] [CrossRef]
- Gedan, K.B.; Silliman, B.R.; Bertness, M.D. Centuries of human-driven change in salt marsh ecosystems. Annu. Rev. Mar. Sci. 2009, 1, 117–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, C.K.; Choy, E.J.; Son, Y.; Lee, J.Y.; Kim, J.K.; Kim, Y.; Lee, K.S. Food web structure of a restored macroalgal bed in the eastern Korean peninsula determined by C and N stable isotope analyses. Mar. Biol. 2008, 153, 1181–1198. [Google Scholar] [CrossRef]
- Kang, C.K.; Kim, J.B.; Lee, K.S.; Kim, J.B.; Lee, P.Y.; Hong, J.S. Trophic importance of benthic microalgae to macrozoobenthos in coastal bay systems in Korea: Dual stable C and N isotope analyses. Mar. Ecol. Prog. Ser. 2003, 259, 79–92. [Google Scholar] [CrossRef]
- Park, H.J.; Han, E.; Lee, Y.J.; Kang, C.K. Trophic linkage of a temperate intertidal macrobenthic food web under opportunistic macroalgal blooms: A stable isotope approach. Mar. Pollut. Bull. 2016, 111, 86–94. [Google Scholar] [CrossRef]
- Kim, C.; Kang, H.Y.; Lee, Y.J.; Yun, S.G.; Kang, C.K. Isotopic variation of macroinvertebrates and their sources of organic matter along an estuarine gradient. Estuar. Coasts 2020, 43, 496–511. [Google Scholar] [CrossRef]
- Graham, W.M.; Condon, R.H.; Carmichael, R.H.; D’Ambra, I.; Patterson, H.K.; Linn, L.J.; Hernandez, F.J. Oil carbon entered the coastal planktonic food web during the Deepwater Horizon oil spill. Environ. Res. Lett. 2010, 5, 045301. [Google Scholar] [CrossRef]
- Spies, R.B.; DesMarais, D.J. Natural isotope study of trophic enrichment of marine benthic communities by petroleum seepage. Mar. Biol. 1983, 73, 67–71. [Google Scholar] [CrossRef]
- Leahy, J.G.; Colwell, R.R. Microbial degradation of hydrocarbons in the environment. Microbiol. Rev. 1990, 54, 305–315. [Google Scholar] [CrossRef]
- Vizzini, S.; Mazzola, A. Seasonal variations in the stable carbon and nitrogen isotope ratios (13C/12C and 15N/14N) of primary producers and consumers in a western Mediterranean coastal lagoon. Mar. Biol. 2003, 142, 1009–1018. [Google Scholar] [CrossRef]
- Kang, H.Y.; Kim, C.; Kim, D.; Lee, Y.J.; Park, H.J.; Kundu, G.K.; Kim, Y.K.; Bibi, R.; Jang, J.; Lee, K.H.; et al. Identifying patterns in the multitrophic community and food-web structure of a low-turbidity temperate estuarine bay. Sci. Rep. 2020, 10, 16637. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.K.; Park, H.J.; Choy, E.J.; Choi, K.S.; Hwang, K.; Kim, J.B. Linking inter-tidal and subtidal food webs: Consumer-mediated transport of intertidal benthic microalgal carbon. PLoS ONE 2015, 10, e0139802. [Google Scholar] [CrossRef]
- Felder, D.L.; Thoma, B.P.; Schmidt, W.E.; Sauvage, T.; Self-Krayesky, S.L.; Chistoserdov, A.; Bracken-Grissom, H.D.; Fredericq, S. Seaweeds and decapod crustaceans on Gulf Deep Banks after the Macondo oil spill. BioScience 2014, 64, 808–819. [Google Scholar] [CrossRef] [Green Version]
- Abrantes, K.G.; Barnett, A.; Bouillon, S. Stable isotope-based community metrics as a tool to identify patterns in food web structure in east African estuaries. Funct. Ecol. 2014, 28, 270–282. [Google Scholar] [CrossRef]
- Romano, C.; Fanelli, E.; D’Anna, G.; Pipitone, C.; Vizzini, S.; Mazzola, A.; Badalamenti, F. Spatial variability of soft-bottom macrobenthic communities in northern Sicily (Western Mediterranean): Contrasting trawled vs. untrawled areas. Mar. Environ. Res. 2016, 122, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Newsome, S.D.; Martinez del Rio, C.; Bearhop, S.; Phillips, D.L. A niche for isotopic ecology. Front. Ecol. Environ. 2007, 5, 429–436. [Google Scholar] [CrossRef]
- Flaherty, E.A.; Ben-David, M. Overlap and partitioning of the ecological and isotopic niches. Oikos 2010, 119, 1409–1416. [Google Scholar] [CrossRef]
- Wlodarska-Kowalczuk, M.; Pearson, T.H. Benthic response to chronic natural physical disturbance by glacial sedimentation in an Arctic fjord. Mar. Ecol. Prog. Ser. 2005, 303, 31–41. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Gurepo | Padori | Yeonpo | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Potential Food Source | δ13C | δ15N | δ13C | δ15N | δ13C | δ15N | ||||||||||
| n | Mean | SD | Mean | SD | n | Mean | SD | Mean | SD | n | Mean | SD | Mean | SD | ||
| July | ||||||||||||||||
| SPOM | 8 | −20.4 | 0.6 | 7.3 | 1.0 | 8 | −20.2 | 1.1 | 7.8 | 1.2 | 6 | −20.3 | 0.9 | 7.5 | 0.8 | |
| Epilithic algae | 6 | −15.4 | 1.0 | 7.7 | 0.8 | 5 | −15.2 | 0.8 | 7.5 | 1.0 | 6 | −15.7 | 0.7 | 7.4 | 0.9 | |
| U. pertusa | 6 | −12.9 | 0.5 | 6.7 | 0.4 | 6 | −12.8 | 0.5 | 6.8 | 0.4 | 6 | −13.0 | 0.6 | 6.9 | 0.4 | |
| S. thunbergii | 6 | −23.6 | 0.4 | 3.5 | 0.4 | 6 | −23.4 | 0.5 | 3.7 | 0.3 | 6 | −23.4 | 0.6 | 3.8 | 0.5 | |
| November | ||||||||||||||||
| SPOM | 7 | −20.8 | 0.9 | 5.4 | 0.9 | 6 | −20.6 | 0.6 | 5.7 | 0.9 | 8 | −20.9 | 1.0 | 5.8 | 0.6 | |
| Epilithic algae | 6 | −14.5 | 0.6 | 5.9 | 0.6 | 6 | −14.9 | 0.7 | 5.6 | 0.6 | 6 | −14.4 | 0.6 | 5.6 | 0.6 | |
| U. pertusa | 6 | −13.8 | 0.5 | 5.8 | 0.4 | 6 | −13.6 | 0.5 | 6.1 | 0.5 | 6 | −13.6 | 0.6 | 5.9 | 0.4 | |
| S. thunbergii | 6 | −23.2 | 0.3 | 4.2 | 0.3 | 6 | −23.0 | 0.3 | 4.0 | 0.3 | 6 | −22.9 | 0.4 | 4.1 | 0.3 | |
| PERMANOVA | Season | Site | Interaction | |||||||||||||
| pseudo-F | p-value | pseudo-F | p-value | pseudo-F | p-value | |||||||||||
| SPOM | 30.52 | 0.001 | 0.47 | 0.746 | 0.01 | 0.966 | ||||||||||
| Epilithic algae | 40.87 | 0.001 | 0.11 | 0.911 | 0.50 | 0.695 | ||||||||||
| U. pertusa | 32.92 | 0.001 | 0.29 | 0.789 | 0.21 | 0.877 | ||||||||||
| S. thunbergii | 6.45 | 0.011 | 0.16 | 0.920 | 0.51 | 0.688 | ||||||||||
| No. | Species Name | Taxa | Gurepo | Padori | Yeonpo | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| δ13C | δ15N | δ13C | δ15N | δ13C | δ15N | ||||||||||||
| n | Mean | SD | Mean | SD | n | Mean | SD | Mean | SD | n | Mean | SD | Mean | SD | |||
| July | |||||||||||||||||
| Suspension feeder | |||||||||||||||||
| 1 | Balanus albicostatus | C | 4 | −17.9 | 0.4 | 6.0 | 0.2 | ||||||||||
| 2 | Chthamalus challengeri | C | 4 | −17.9 | 0.3 | 6.2 | 0.3 | 4 | −17.9 | 0.4 | 6.2 | 0.3 | 4 | −17.4 | 0.3 | 6.2 | 0.3 |
| 3 | Crassostrea gigas | B | 4 | −17.5 | 0.4 | 7.2 | 0.5 | 4 | −17.4 | 0.2 | 6.9 | 0.6 | 4 | −17.7 | 0.3 | 7.1 | 0.5 |
| 4 | Ditrupa arietina | P | 3 | −19.1 | 0.4 | 7.1 | 0.3 | ||||||||||
| 5 | Hydroides ezoensis | P | 3 | −17.6 | 0.3 | 7.0 | 0.3 | 3 | −17.9 | 0.5 | 6.8 | 0.6 | |||||
| 6 | Lasaea undulata | B | 3 | −18.1 | 0.5 | 6.8 | 0.5 | 4 | −18.1 | 0.4 | 6.3 | 0.3 | 4 | −18.4 | 0.4 | 6.4 | 0.4 |
| 7 | Mytilus galloprovincialis | B | 4 | −18.4 | 0.3 | 6.4 | 0.3 | 4 | −18.4 | 0.4 | 6.5 | 0.5 | 4 | −18.4 | 0.4 | 6.8 | 0.5 |
| Deposit feeder | |||||||||||||||||
| 8 | Barleeia angustata | G | 4 | −15.8 | 0.4 | 7.9 | 0.5 | 4 | −15.6 | 0.6 | 7.9 | 0.4 | 4 | −15.8 | 0.4 | 8.1 | 0.4 |
| 9 | Gammarian amphipod | C | 4 | −16.4 | 0.3 | 7.6 | 0.5 | 4 | −16.3 | 0.4 | 7.8 | 0.4 | |||||
| 10 | Nainereis laevigata | P | 3 | −15.3 | 0.4 | 7.4 | 0.4 | ||||||||||
| 11 | Parhyale sp. | C | 4 | −14.8 | 0.3 | 9.5 | 0.5 | 4 | −14.9 | 0.5 | 9.7 | 0.4 | 4 | −14.9 | 0.3 | 9.6 | 0.4 |
| Grazer | |||||||||||||||||
| 12 | Chlorostoma lischkei | G | 4 | −16.2 | 0.5 | 9.1 | 0.2 | 4 | −15.6 | 0.5 | 8.7 | 0.5 | |||||
| 13 | Chlorostoma turbinata | G | 4 | −16.6 | 0.3 | 9.2 | 0.7 | 4 | −16.6 | 0.4 | 8.7 | 0.6 | |||||
| 14 | Heminerita japonica | G | 4 | −16.0 | 0.4 | 7.9 | 0.5 | ||||||||||
| 15 | Littorina brevicula | G | 4 | −13.9 | 0.4 | 8.6 | 0.4 | 4 | −13.1 | 0.6 | 8.8 | 0.4 | |||||
| 16 | Lottia dorsuosa | G | 4 | −13.6 | 0.6 | 8.6 | 0.3 | 4 | −13.5 | 0.2 | 9.0 | 0.6 | |||||
| 17 | Lottia kogamogai | G | 4 | −14.1 | 0.5 | 9.7 | 0.4 | ||||||||||
| 18 | Lottia tenuisculpta | G | 4 | −13.1 | 0.3 | 9.0 | 0.5 | 4 | −13.0 | 0.4 | 8.9 | 0.3 | 4 | −13.0 | 0.3 | 9.5 | 0.6 |
| 19 | Monodonta neritoides | G | 4 | −15.0 | 0.4 | 8.9 | 0.6 | ||||||||||
| 20 | Nipponacmaea schrenckii | G | 4 | −15.7 | 0.4 | 9.0 | 0.6 | ||||||||||
| 21 | Nodilittorina radiata | G | 4 | −15.9 | 0.4 | 8.6 | 0.3 | ||||||||||
| 22 | Omphalius rusticus | G | 4 | −15.0 | 0.4 | 9.0 | 0.3 | ||||||||||
| 23 | Patelliida conulus | G | 4 | −13.2 | 0.4 | 9.6 | 0.4 | ||||||||||
| 24 | Patelloida pygmaea | G | 4 | −15.8 | 0.4 | 9.0 | 0.7 | ||||||||||
| 25 | Turbo coronata coreensis | G | 4 | −13.2 | 0.5 | 10.2 | 0.5 | 4 | −13.4 | 0.5 | 10.5 | 0.5 | |||||
| 26 | Acanthoschitona achates | Pp | 4 | −13.8 | 0.4 | 8.1 | 0.8 | 4 | −13.6 | 0.4 | 8.1 | 0.6 | |||||
| Omnivore | |||||||||||||||||
| 27 | Asterina pectinifera | E | 4 | −12.7 | 0.5 | 10.9 | 0.4 | 4 | −12.7 | 0.3 | 11.0 | 0.4 | 4 | −12.7 | 0.5 | 10.9 | 0.4 |
| 28 | Hemigrapsus penicillatus | C | 4 | −17.7 | 0.3 | 11.2 | 0.6 | 4 | −16.0 | 0.5 | 11.0 | 0.6 | 4 | −15.7 | 0.4 | 10.7 | 0.5 |
| 29 | Pagurus sp. | C | 4 | −14.7 | 0.4 | 10.6 | 0.4 | 4 | −14.4 | 0.5 | 11.3 | 0.3 | 4 | −15.3 | 0.4 | 10.9 | 0.3 |
| Carnivore | |||||||||||||||||
| 30 | Ceratostoma inornatum | G | 4 | −15.3 | 0.5 | 11.9 | 0.6 | 4 | −15.6 | 0.5 | 11.8 | 0.8 | 4 | −15.4 | 0.5 | 12.0 | 0.3 |
| 31 | Ceratostoma rorifluum | G | 4 | −16.5 | 0.5 | 12.2 | 0.7 | 4 | −16.5 | 0.6 | 11.8 | 0.5 | 4 | −16.2 | 0.4 | 12.1 | 0.6 |
| 32 | Hima fratercula hypolia | G | 4 | −14.6 | 0.4 | 12.3 | 0.4 | ||||||||||
| 33 | Thais clavigera | G | 4 | −16.6 | 0.4 | 11.9 | 0.5 | 4 | −16.7 | 0.4 | 12.0 | 0.4 | 4 | −16.5 | 0.5 | 12.5 | 0.4 |
| November | |||||||||||||||||
| Suspension feeder | |||||||||||||||||
| 1 | Balanus albicostatus | C | 4 | −18.9 | 0.5 | 4.7 | 0.6 | ||||||||||
| 2 | Chthamalus challengeri | C | 4 | −18.8 | 0.4 | 5.1 | 0.3 | 4 | −18.7 | 0.3 | 5.1 | 0.2 | 4 | −19.1 | 0.2 | 4.9 | 0.3 |
| 3 | Crassostrea gigas | B | 4 | −18.2 | 0.2 | 5.7 | 0.6 | 4 | −18.0 | 0.3 | 6.0 | 0.5 | 4 | −18.5 | 0.3 | 6.1 | 0.3 |
| 4 | Hydroides ezoensis | P | 3 | −18.1 | 0.2 | 5.8 | 0.5 | 3 | −18.2 | 0.4 | 5.8 | 0.5 | |||||
| 5 | Lasaea undulata | B | 4 | −18.7 | 0.3 | 5.9 | 0.3 | 4 | −18.7 | 0.3 | 5.5 | 0.2 | 4 | −19.1 | 0.3 | 5.4 | 0.3 |
| 6 | Mytilus galloprovincialis | B | 4 | −19.2 | 0.2 | 5.4 | 0.4 | 4 | −19.0 | 0.3 | 5.8 | 0.4 | 4 | −19.2 | 0.3 | 6.0 | 0.3 |
| Deposit feeder | |||||||||||||||||
| 7 | Barleeia angustata | G | 4 | −16.2 | 0.3 | 6.7 | 0.3 | 4 | −16.5 | 0.3 | 6.7 | 0.4 | 4 | −16.8 | 0.3 | 7.0 | 0.2 |
| 8 | Gammarian amphipod | C | 4 | −17.2 | 0.4 | 6.1 | 0.3 | 4 | −17.8 | 0.3 | 6.6 | 0.3 | 4 | −17.4 | 0.6 | 6.8 | 0.4 |
| 9 | Nainereis laevigata | P | 3 | −16.2 | 0.3 | 8.4 | 0.5 | ||||||||||
| 10 | Parhyale sp. | C | 4 | −15.8 | 0.2 | 8.7 | 0.5 | 4 | −16.2 | 0.5 | 8.5 | 0.5 | 4 | −15.9 | 0.4 | 8.6 | 0.4 |
| Grazer | |||||||||||||||||
| 11 | Chlorostoma lischkei | G | 4 | −16.9 | 0.4 | 8.5 | 0.6 | 4 | −16.2 | 0.5 | 8.5 | 0.4 | |||||
| 12 | Chlorostoma turbinata | G | 4 | −17.2 | 0.4 | 8.8 | 0.5 | 4 | −17.2 | 0.4 | 8.4 | 0.3 | |||||
| 13 | Heminerita japonica | G | 4 | −16.7 | 0.4 | 7.8 | 0.4 | ||||||||||
| 14 | Littorina brevicula | G | 3 | −15.0 | 0.3 | 8.1 | 0.6 | 3 | −14.9 | 0.3 | 8.2 | 0.2 | |||||
| 15 | Lottia dorsuosa | G | 4 | −14.1 | 0.4 | 7.2 | 0.4 | 4 | −14.2 | 0.5 | 8.0 | 0.6 | |||||
| 16 | Lottia tenuisculpta | G | 4 | −13.8 | 0.4 | 7.4 | 0.6 | 4 | −14.0 | 0.3 | 7.2 | 0.2 | 4 | −13.8 | 0.2 | 8.5 | 0.6 |
| 17 | Omphalius rusticus | G | 3 | −15.0 | 0.6 | 7.5 | 0.4 | 3 | −15.9 | 0.4 | 7.6 | 0.5 | |||||
| 18 | Turbo coronata coreensis | G | −14.1 | 0.3 | 8.8 | 0.3 | 4 | −14.6 | 0.4 | 9.2 | 0.3 | ||||||
| 19 | Acanthoschitona achates | Pp | 4 | −14.2 | 0.3 | 6.2 | 0.2 | 4 | −14.6 | 0.5 | 6.9 | 0.4 | |||||
| Omnivore | |||||||||||||||||
| 20 | Asterina pectinifera | E | 4 | −14.0 | 0.5 | 9.7 | 0.3 | 4 | −14.8 | 0.4 | 9.9 | 0.4 | 4 | −14.8 | 0.6 | 9.9 | 0.4 |
| 21 | Hemigrapsus penicillatus | C | 4 | −18.1 | 0.2 | 10.0 | 0.5 | 4 | −16.8 | 0.4 | 9.8 | 0.5 | 4 | −16.9 | 0.4 | 9.8 | 0.4 |
| 22 | Pagurus sp. | C | 4 | −15.2 | 0.3 | 9.7 | 0.2 | 4 | −15.3 | 0.4 | 10.0 | 0.3 | 4 | −15.3 | 0.5 | 9.9 | 0.5 |
| Carnivore | |||||||||||||||||
| 23 | Ceratostoma inornatum | G | 4 | −16.0 | 0.4 | 11.0 | 0.4 | 4 | −16.2 | 0.3 | 10.9 | 0.2 | 4 | −15.9 | 0.4 | 11.1 | 0.4 |
| 24 | Ceratostoma rorifluum | G | 4 | −17.0 | 0.2 | 10.8 | 0.4 | 4 | −17.1 | 0.5 | 10.6 | 0.4 | 4 | −17.0 | 0.3 | 10.8 | 0.4 |
| 25 | Hima fratercula hypolia | G | 4 | −15.5 | 0.4 | 11.3 | 0.2 | ||||||||||
| 26 | Thais clavigera | G | 4 | −17.2 | 0.4 | 11.0 | 0.3 | 4 | −17.1 | 0.3 | 11.0 | 0.2 | 4 | −17.0 | 0.3 | 11.4 | 0.5 |
| Gurepo | Padori | Yeonpo | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Potential Food Source | δ13C | δ15N | δ13C | δ15N | δ13C | δ15N | ||||||||||
| n | Mean | SD | Mean | SD | n | Mean | SD | Mean | SD | n | Mean | SD | Mean | SD | ||
| July | ||||||||||||||||
| Suspension feeder | 22 | −17.9 | 0.4 | 6.6 | 0.6 | 22 | −18.1 | 0.6 | 6.6 | 0.5 | 16 | −17.9 | 0.5 | 6.6 | 0.5 | |
| Deposit feeder | 11 | −15.3 | 0.6 | 8.3 | 2.5 | 12 | −15.6 | 0.8 | 8.4 | 1.0 | 12 | −15.7 | 0.7 | 8.5 | 0.9 | |
| Grazer | 28 | −14.4 | 1.4 | 8.9 | 0.6 | 28 | −14.9 | 1.3 | 9.0 | 0.7 | 36 | −14.3 | 1.3 | 9.0 | 0.9 | |
| Omnivore | 12 | −15.0 | 2.2 | 10.9 | 0.5 | 12 | −14.4 | 1.5 | 11.1 | 0.4 | 12 | −14.6 | 1.5 | 10.8 | 0.4 | |
| Carnivore | 12 | −16.2 | 0.7 | 12.0 | 0.6 | 16 | −15.8 | 1.0 | 12.0 | 0.5 | 12 | −16.0 | 0.6 | 12.2 | 0.5 | |
| November | ||||||||||||||||
| Suspension feeder | 23 | −18.7 | 0.5 | 5.4 | 0.6 | 19 | −18.6 | 0.5 | 5.6 | 0.5 | 16 | −19.0 | 0.4 | 5.6 | 0.5 | |
| Deposit feeder | 15 | −16.4 | 0.6 | 7.4 | 1.2 | 12 | −16.8 | 0.8 | 7.3 | 1.0 | 12 | −16.7 | 0.8 | 7.5 | 0.9 | |
| Grazer | 26 | −15.2 | 1.4 | 7.7 | 0.9 | 20 | −15.5 | 1.3 | 8.1 | 0.7 | 23 | −14.8 | 1.0 | 8.1 | 0.8 | |
| Omnivore | 12 | −15.8 | 1.8 | 9.8 | 0.4 | 12 | −15.6 | 1.0 | 9.9 | 0.4 | 12 | −15.7 | 1.0 | 9.9 | 0.4 | |
| Carnivore | 12 | −16.8 | 0.6 | 11.0 | 0.3 | 16 | −16.5 | 0.8 | 11.0 | 0.3 | 12 | −16.6 | 0.6 | 11.1 | 0.4 | |
| PERMANOVA | Season | Site | Interaction | |||||||||||||
| pseudo-F | p-value | pseudo-F | p-value | pseudo-F | p-value | |||||||||||
| Suspension feeder | 104.90 | 0.001 | 0.55 | 0.663 | 1.01 | 0.391 | ||||||||||
| Deposit feeder | 35.58 | 0.001 | 1.07 | 0.355 | 0.36 | 0.735 | ||||||||||
| Grazer | 36.33 | 0.001 | 1.47 | 0.212 | 0.36 | 0.815 | ||||||||||
| Omnivore | 33.20 | 0.001 | 0.58 | 0.626 | 0.27 | 0.816 | ||||||||||
| Carnivore | 50.05 | 0.001 | 0.79 | 0.533 | 0.11 | 0.901 | ||||||||||
| Isotopic Niche Metrics | ||||||||
|---|---|---|---|---|---|---|---|---|
| Feeding Group | Gurepo | Padori | Yeonpo | Percentage Overlap (%) | ||||
| TA | SEAc | TA | SEAc | TA | SEAc | G and Y | P and Y | |
| July | ||||||||
| Suspension feeder | 2.32 | 0.80 | 2.77 | 0.99 | 2.01 | 0.92 | 80 | 84 |
| Deposit feeder | 3.05 | 1.51 | 2.92 | 1.64 | 2.67 | 1.23 | 60 | 82 |
| Grazer | 8.18 | 2.98 | 8.31 | 2.40 | 9.45 | 3.23 | 76 | 62 |
| Omnivore | 6.38 | 3.45 | 4.00 | 2.20 | 3.27 | 1.89 | 71 | 60 |
| Carnivore | 2.73 | 1.48 | 3.30 | 1.70 | 2.13 | 0.99 | 66 | 55 |
| November | ||||||||
| Suspension feeder | 2.04 | 0.80 | 1.97 | 0.70 | 1.31 | 0.66 | 41 | 40 |
| Deposit feeder | 3.89 | 1.83 | 3.62 | 1.95 | 3.21 | 1.70 | 54 | 83 |
| Grazer | 7.29 | 3.17 | 6.02 | 2.86 | 6.08 | 2.71 | 48 | 62 |
| Omnivore | 5.22 | 2.53 | 2.34 | 1.24 | 2.55 | 1.38 | 57 | 92 |
| Carnivore | 1.37 | 0.77 | 2.09 | 0.91 | 1.57 | 0.80 | 61 | 61 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, H.J.; Kim, C.; Kang, C.-K. Recovery of Macrobenthic Food Web on Rocky Shores Following the Hebei Spirit Oil Spill as Revealed by C and N Stable Isotopes. Water 2022, 14, 2335. https://doi.org/10.3390/w14152335
Park HJ, Kim C, Kang C-K. Recovery of Macrobenthic Food Web on Rocky Shores Following the Hebei Spirit Oil Spill as Revealed by C and N Stable Isotopes. Water. 2022; 14(15):2335. https://doi.org/10.3390/w14152335
Chicago/Turabian StylePark, Hyun Je, Changseong Kim, and Chang-Keun Kang. 2022. "Recovery of Macrobenthic Food Web on Rocky Shores Following the Hebei Spirit Oil Spill as Revealed by C and N Stable Isotopes" Water 14, no. 15: 2335. https://doi.org/10.3390/w14152335
APA StylePark, H. J., Kim, C., & Kang, C. -K. (2022). Recovery of Macrobenthic Food Web on Rocky Shores Following the Hebei Spirit Oil Spill as Revealed by C and N Stable Isotopes. Water, 14(15), 2335. https://doi.org/10.3390/w14152335