

Interactions between Cultivated Gracilariopsis lemaneiformis and Floating Sargassum horneri under Controlled Laboratory Conditions


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Maintenance
2.2. Culture Experiment
2.3. Measurement of Photosynthesis
2.4. Measurement of NO3-N Uptake Rate
2.5. Measurement of Chlorophyll a (Chl a)
2.6. Measurements of Soluble Protein and Carbohydrate
2.7. Statistical Analysis
3. Results
3.1. Growth
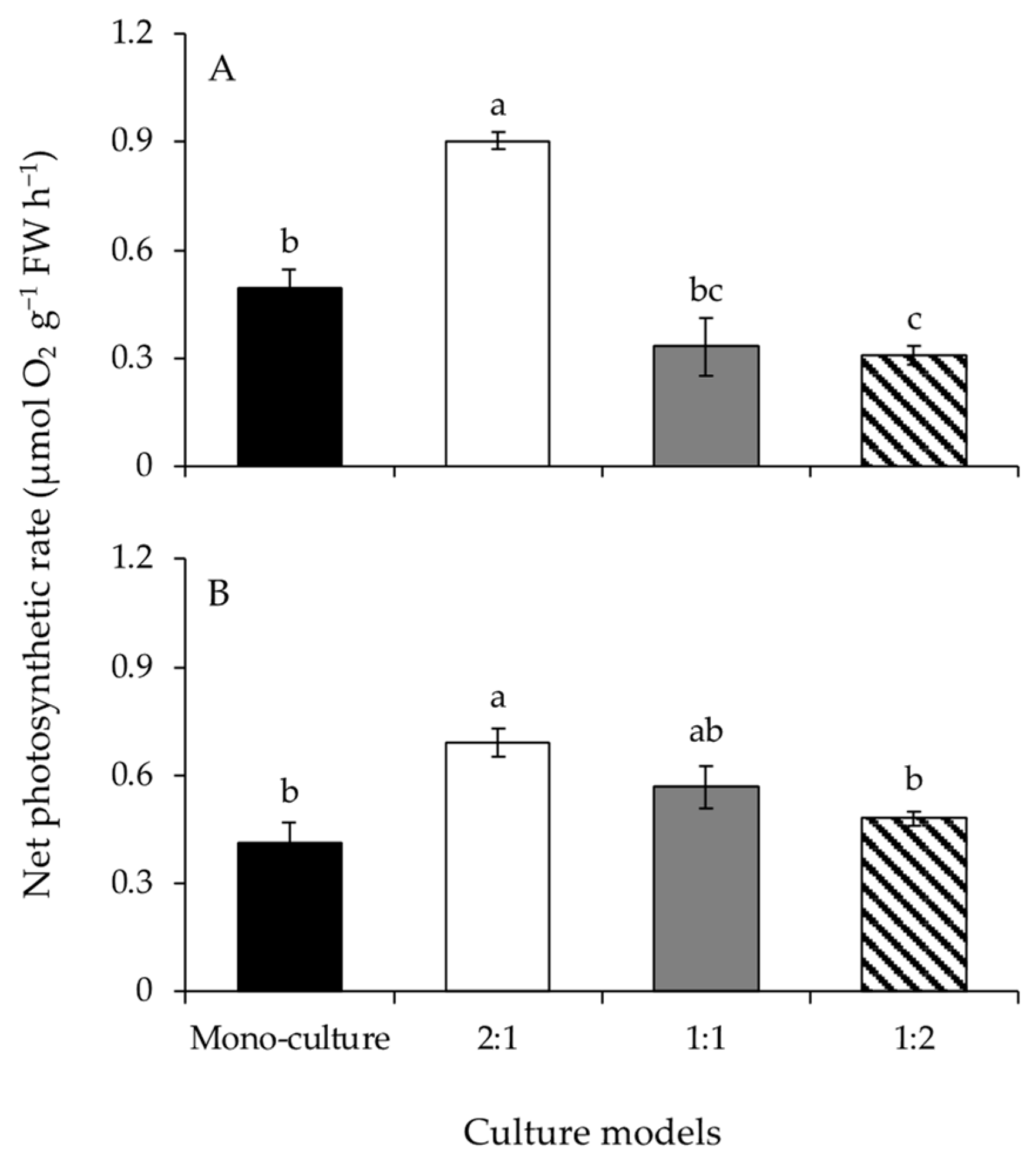
3.2. Photosynthesis
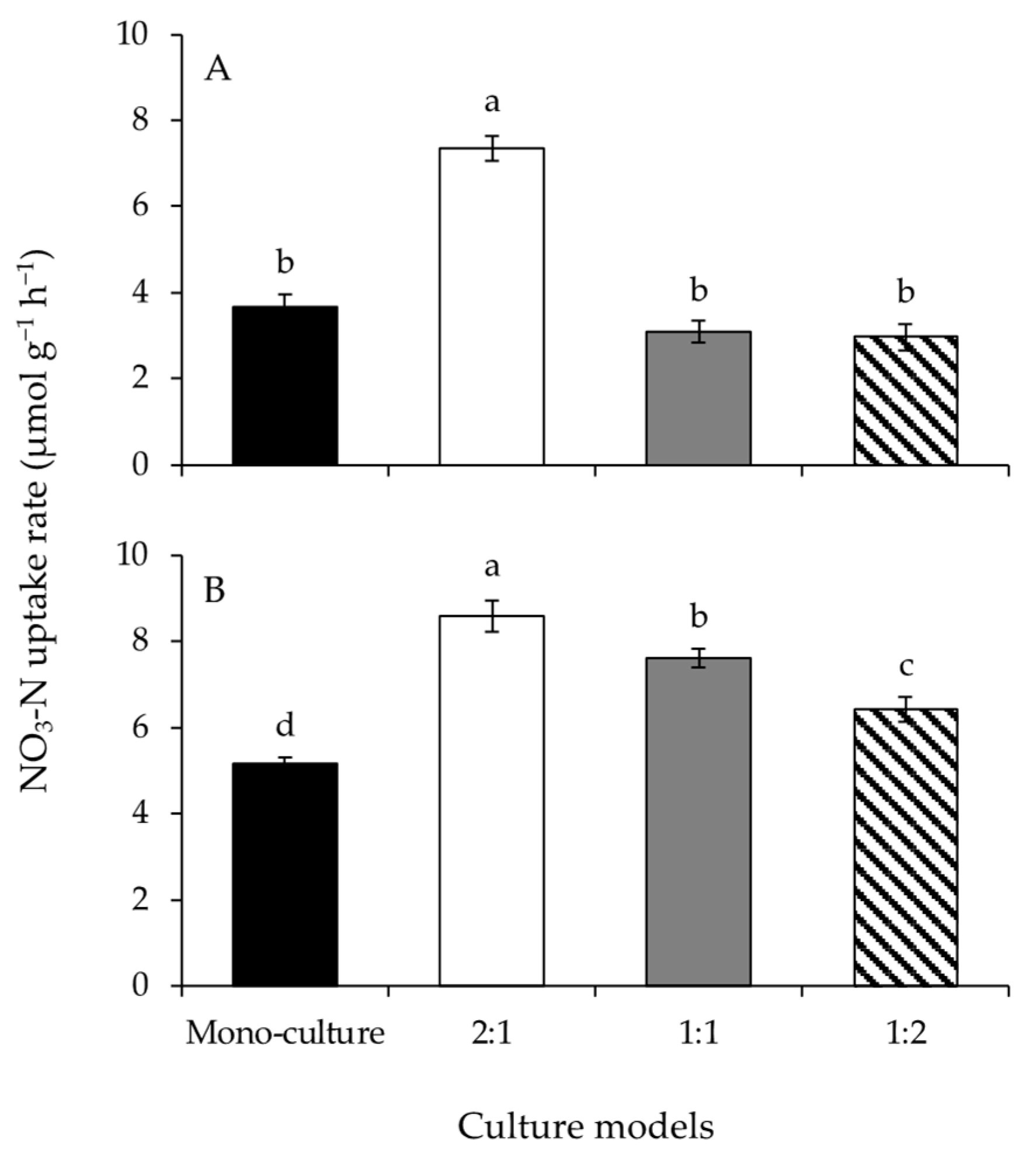
3.3. NO3-N Uptake Rate
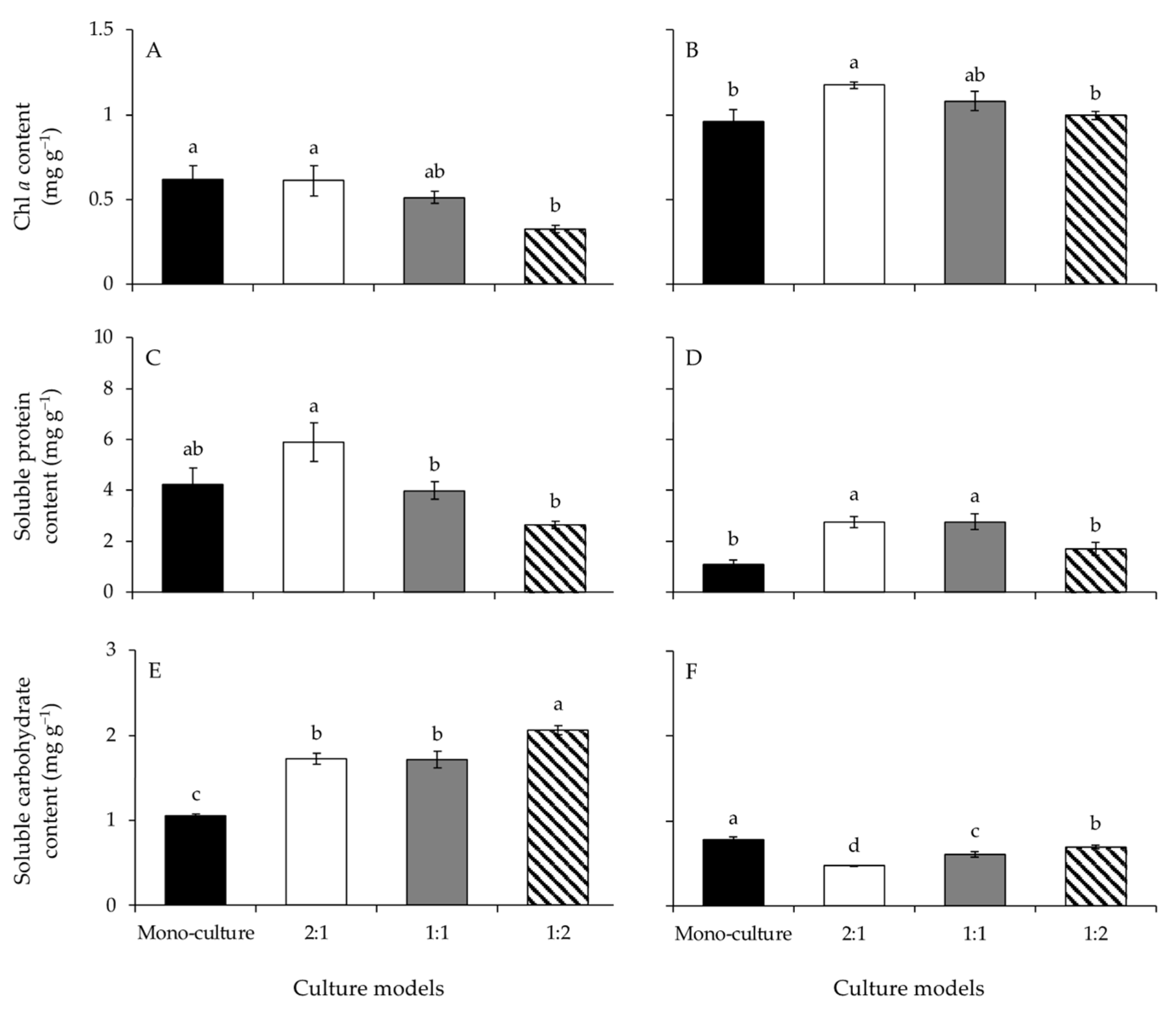
3.4. Chl a
3.5. Soluble Protein and Carbohydrate
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Komatsu, T.; Tatsukawa, K.; Filippi, J.B.; Sagawa, T.; Matsunaga, D.; Mikami, A.; Ishida, K.; Ajisaka, T.; Tanaka, K.; Aoki, M.; et al. Distribution of drifting seaweeds in eastern East China Sea. J. Mar. Syst. 2007, 67, 245–252. [Google Scholar] [CrossRef]
- Schell, J.M.; Goodwin, D.S.; Siuda, A.N.S. Recent Sargassum inundation events in the Caribbean: Shipboard observations reveal dominance of a previously rare form. Oceanography 2015, 28, 8–10. [Google Scholar] [CrossRef]
- Xing, Q.G.; Guo, R.H.; Wu, L.L.; An, D.Y.; Cong, M.; Qin, S.; Li, X.R. High-resolution satellite observations of a new hazard of golden tides caused by floating Sargassum in winter in the Yellow Sea. IEEE Geosci. Remote Sens. Lett. 2017, 14, 1815–1819. [Google Scholar] [CrossRef]
- García-Sánchez, M.; Graham, C.; Vera, E.; Escalante-Mancera, E.; Álvarez-Filip, L.; van Tussenbroek, B.I. Temporal changes in the composition and biomass of beached pelagic Sargassum species in the Mexican Caribbean. Aquat. Bot. 2020, 167, 103275. [Google Scholar] [CrossRef]
- Lapointe, B.E. A comparison of nutrient-limited productivity in Sargassum natans from neritic vs. oceanic waters of the western North Atlantic Ocean. Limnol. Oceanogr. 1995, 40, 625–633. [Google Scholar] [CrossRef]
- Huffard, C.L.; von Thun, S.; Sherman, A.D.; Sealey, K.; Smith, K.L. Pelagic Sargassum community change over a 40-year period: Temporal and spatial variability. Mar. Biol. 2014, 161, 2735–2751. [Google Scholar] [CrossRef]
- Hu, Z.M.; Uwai, S.; Yu, S.H.; Komatsu, T.; Ajisaka, T.; Duan, D.L. Phylogeographic heterogeneity of the brown macroalga Sargassum horneri (Fucaceae) in the northwestern Pacific in relation to late Pleistocene glaciation and tectonic configurations. Mol. Ecol. 2011, 20, 3894–3909. [Google Scholar] [CrossRef]
- Xu, M.; Sakamoto, S.; Komatsu, T. Attachment strength of the subtidal seaweed Sargassum horneri (Turner) C. Agardh varies among development stages and depths. J. Appl. Phycol. 2016, 28, 3679–3687. [Google Scholar] [CrossRef]
- Sun, J.Z.; Chen, W.D.; Zhuang, D.G.; Zheng, H.Y.; Li, L.; Pang, S.J. In situ ecological studies of the subtidal brown alga Sargasssum horneri at Nanji Island of China. S. China Fish. Sci. 2008, 4, 58–63, (In Chinese with English abstract). [Google Scholar]
- Qi, L.; Hu, C.M.; Wang, M.Q.; Shang, S.L.; Wilson, C. Floating algae blooms in the East China Sea. Geophys. Res. Lett. 2017, 44, 11501–11509. [Google Scholar] [CrossRef]
- Smetacek, V.; Zingone, A. Green and golden seaweed tides on the rise. Nature 2013, 504, 84–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz-Rivera, E.; Flores-Díaz, M.; Hawkins, A. A fish kill coincident with dense Sargassum accumulation in a tropical bay. Bull. Mar. Sci. 2015, 91, 455–456. [Google Scholar] [CrossRef]
- Solarin, B.B.; Bolaji, D.A.; Fakayode, O.S.; Akinnigbagbe, R.O. Impacts of an invasive seaweed Sargassum hystrix var. fluitans (Børgesen 1914) on the fisheries and other economic implications for the Nigerian coastal waters. IOSR J. Agric. Vet. Sci. 2014, 7, 1–6. [Google Scholar]
- Milledge, J.J.; Harvey, P.J. Golden tides: Problem or golden opportunity? The valorisation of Sargassum from beach inundations. J. Mar. Sci. Eng. 2016, 4, 60. [Google Scholar] [CrossRef]
- Wang, X.M.; Shan, T.F.; Pang, S.J.; Su, L. Assessment of the genetic relationship between the recently established benthic population and the adjacent floating populations of Sargassum horneri (Phaeophyceae) in Dalian of China by newly developed trinucleotide microsatellite markers. J. Appl. Phycol. 2019, 31, 3989–3996. [Google Scholar] [CrossRef]
- Dickie, I.A.; Schnitzer, S.A.; Reich, P.B.; Hobbie, S.E. Spatially disjunct effects of co-occurring competition and facilitation. Ecol. Lett. 2005, 8, 1191–1200. [Google Scholar] [CrossRef] [PubMed]
- Narwani, A.; Bentlage, B.; Alexandrou, M.A.; Fritschie, K.J.; Delwiche, C.; Oakley, T.H.; Cardinale, B.J. Ecological interactions and coexistence are predicted by gene expression similarity in freshwater green algae. J. Ecol. 2017, 105, 580–591. [Google Scholar] [CrossRef]
- Cameron, H.; Coulson, T.; Marshall, D.J. Size and density mediate transitions between competition and facilitation. Ecol. Lett. 2019, 22, 1879–1888. [Google Scholar] [CrossRef]
- Choi, H.G.; Norton, T.A. Competition and facilitation between germlings of Ascophyllum nodosum and Fucus vesiculosus. Mar. Biol. 2005, 147, 525–532. [Google Scholar] [CrossRef]
- White, L.F.; Shurin, J.B. Density dependent effects of an exotic marine macroalga on native community diversity. J. Exp. Mar. Biol. Ecol. 2011, 405, 111–119. [Google Scholar] [CrossRef]
- Gavina, M.K.A.; Tahara, T.; Tainaka, K.; Ito, H.; Morita, S.; Ichinose, G.; Okabe, T.; Togashi, T.; Nagatani, T.; Yoshimura, J. Multi-species coexistence in Lotka–Volterra competitive systems with crowding effects. Sci. Rep. 2018, 8, 1198. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Lin, L.; Ma, Z.; Zhang, T.; Chen, W.; Zou, D. Carbon and nitrogen accumulation and interspecific competition in two algae species, Pyropia haitanensis and Ulva lactuca, under ocean acidification conditions. Aquac. Int. 2019, 27, 721–733. [Google Scholar] [CrossRef]
- Li, X.S.; Xu, J.T.; He, P.M. Comparative research on inorganic carbon acquisition by the macroalgae Ulva prolifera (Chlorophyta) and Pyropia yezoensis (Rhodophyta). J. Appl. Phycol. 2016, 28, 491–497. [Google Scholar] [CrossRef]
- Sun, J.Y.; Bao, M.L.; Xu, T.P.; Li, F.T.; Wu, H.L.; Li, X.S.; Xu, J.T. Elevated CO2 influences competition for growth, photosynthetic performance and biochemical composition in Neopyropia yezoensis and Ulva prolifera. Algal Res. 2021, 56, 102313. [Google Scholar] [CrossRef]
- Sukenik, A.; Eshkol, R.; Livne, A.; Hadas, O.; Rom, M.; Tchernov, D.; Vardi, A.; Kaplan, A. Inhibition of growth and photosynthesis of the dinoflagellate Peridinium gatunense by Microcystis sp. (Cyanobacteria): A novel allelopathic mechanism. Limnol. Oceanogr. 2002, 47, 1656–1663. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, Z.M.; Song, X.X.; Tang, X.X.; Zhang, S.D. Effects of macroalgae Ulva pertusa (Chlorophyta) and Gracilaria lemaneiformis (Rhodophyta) on growth of four species of bloom-forming dinoflagellates. Aquat. Bot. 2007, 86, 139–147. [Google Scholar] [CrossRef]
- He, D.; Liu, J.; Hao, Q.; Ran, L.H.; Zhou, B.; Tang, X.X. Interspecific competition and allelopathic interaction between Karenia mikimotoi and Dunaliella salina in laboratory culture. Chin. J. Oceanol. Limnol. 2016, 34, 301–313. [Google Scholar] [CrossRef]
- Yuan, C.Y.; Zhao, Y.; Yang, S.; Cui, Q.M. Allelopathic effect of Sargassum fusiforme on growth of Karenia mikimotoi. Appl. Mech. Mater. 2014, 522–524, 721–724. [Google Scholar] [CrossRef]
- Patil, V.; Abate, R.; Yang, Y.F.; Zhang, J.W.; Lin, H.N.; Chen, C.P.; Liang, J.R.; Sun, L.; Li, X.S.; Gao, Y.H. Allelopathic effect of Pyropia haitanensis (Rhodophyta) on the bloom-forming Skeletonema costatum (Bacillariophyta). J. Appl. Phycol. 2020, 32, 1275–1286. [Google Scholar] [CrossRef]
- Steentoft, M.; Irvine, L.M.; Farnham, W.F. Two terete species of Gracilaria and Gracilariopsis (Gracilariales, Rhodophyta) in Britain. Phycologia 1995, 34, 113–127. [Google Scholar] [CrossRef]
- Yoshida, T. Marine Algae of Japan; Uchida Rokakuho Publishing Company: Tokyo, Japan, 1998. (In Japanese) [Google Scholar]
- Iyer, R.; De Clerck, O.; Bolton, J.J.; Coyne, V.E. Morphological and taxonomic studies of Gracilaria and Gracilariopsis species (Gracilariales, Rhodophyta) from South Africa. S. Afr. J. Bot. 2004, 70, 521–539. [Google Scholar] [CrossRef]
- Qi, Z.H.; Liu, H.M.; Li, B.; Mao, Y.Z.; Jiang, Z.J.; Zhang, J.H.; Fang, J.G. Suitability of two seaweeds, Gracilaria lemaneiformis and Sargassum pallidum, as feed for the abalone Haliotis discus hannai Ino. Aquaculture 2010, 300, 189–193. [Google Scholar] [CrossRef]
- Liu, X.J.; Zhang, Q.F.; Huan, Z.Y.; Zhong, M.Q.; Chen, W.Z.; Du, H. Identification and characterization of glutamine synthetase isozymes in Gracilaria lemaneiformis. Aquat. Bot. 2018, 146, 23–30. [Google Scholar] [CrossRef]
- Yuan, S.L.; Duan, Z.H.; Lu, Y.N.; Ma, X.L.; Wang, S. Optimization of decolorization process in agar production from Gracilaria lemaneiformis and evaluation of ntioxidant activities of the extract rich in natural pigments. 3 Biotech 2018, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Fishery Bureau of Agriculture Ministry of China. Chinese Fishery Statistical Yearbook; China Agriculture Press: Beijing, China, 2021; pp. 23–28. (In Chinese)
- Yang, Y.F.; Chai, Z.Y.; Wang, Q.; Chen, W.Z.; He, Z.L.; Jiang, S.J. Cultivation of seaweed Gracilaria in Chinese coastal waters and its contribution to environmental improvements. Algal Res. 2015, 9, 236–244. [Google Scholar] [CrossRef]
- Kim, J.K.; Yarish, C.; Hwang, E.K.; Park, M.; Kim, Y. Seaweed aquaculture: Cultivation technologies, challenges and its ecosystem services. Algae 2017, 32, 1–13. [Google Scholar] [CrossRef]
- Gao, X.; Kim, J.H.; Park, S.K.; Yu, O.H.; Kim, Y.S.; Choi, H.G. Diverse responses of sporophytic photochemical efficiency and gametophytic growth for two edible kelps, Saccharina japonica and Undaria pinnatifida, to ocean acidification and warming. Mar. Pollut. Bull. 2019, 142, 315–320. [Google Scholar] [CrossRef]
- Li, S.F.; Liu, Y.; Gong, Q.L.; Gao, X.; Li, J.Y. Physiological and ultrastructural responses of the brown seaweed Undaria pinnatifida to triphenyltin chloride (TPTCL) stress. Mar. Pollut. Bull. 2020, 153, 110978. [Google Scholar] [CrossRef]
- Liu, C.X.; Zou, D.H.; Yang, Y.F.; Chen, B.B.; Jiang, H. Temperature responses of pigment contents, chlorophyll fluorescence characteristics, and antioxidant defenses in Gracilariopsis lemaneiformis (Gracilariales, Rhodophyta) under different CO2 levels. J. Appl. Phycol. 2017, 29, 983–991. [Google Scholar] [CrossRef]
- Wang, Y.; Feng, Y.Q.; Liu, X.J.; Zhong, M.Q.; Chen, W.Z.; Wang, F.; Du, H. Response of Gracilaria lemaneiformis to nitrogen deprivation. Algal Res. 2018, 34, 82–96. [Google Scholar] [CrossRef]
- Yang, Y.L.; Li, W.; Li, Y.H.; Xu, N.J. Photophysiological responses of the marine macroalga Gracilariopsis lemaneiformis to ocean acidification and warming. Mar. Environ. Res. 2020, 163, 105204. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.R.; Pang, T.; Liu, J.G. Changes in the temperature tolerance profile of Gracilariopsis lemaneiformis from the perspective of photosynthesis, respiration, and biochemical markers after many years of vegetative propagation. J. Appl. Phycol. 2022, 34, 1045–1058. [Google Scholar] [CrossRef]
- Tatewaki, M. Formation of a crustaceous sporophyte with unilocular sporangia in Scytosiphon lomentaria. Phycologia 1966, 6, 62–66. [Google Scholar] [CrossRef]
- Li, S.; Liu, D. Improvement of spectrophotometric determination of phosphorus in water by phospho–molybdenum blue. Environ. Prot. Chem. Ind. 2006, 26, 78–80, (In Chinese with English abstract). [Google Scholar]
- Sun, X.Y.; Hong, L.C.; Ye, H.M. Experiment determining nitrate nitrogen in water samples by on-line cadmium column reduction-flow injection method. Water Resour. Prot. 2010, 26, 75–77, (In Chinese with English abstract). [Google Scholar]
- Kochert, G. Protein determination by dye binding. In Handbook of Phycological Methods. Physiological and Biochemical Methods; Hellebust, J.A., Craigie, J.S., Eds.; Cambridge University Press: Cambridge, UK, 1978; pp. 91–93. [Google Scholar]
- Tang, S.Q.; Gong, M.G.; Chen, D.C. Study of methods for determination of carbohydrates in sea water 2. The anthrone method for total amount of particulate carbohydrate. J. Shandong Coll. Oceanol. 1985, 15, 47–54. (In Chinese) [Google Scholar]
- Hirsch, A.M.; Bauer, W.D.; Bird, D.M.; Cullimore, J.; Tyler, B.; Yoder, J.I. Molecular signals and receptors: Controlling rhizosphere interactions between plants and other organisms. Ecology 2003, 84, 858–868. [Google Scholar] [CrossRef]
- Xu, D.; Gao, Z.Q.; Zhang, X.F.; Fan, X.; Wang, Y.T.; Li, D.M.; Wang, W.; Zhuang, Z.M.; Ye, N.H. Allelopathic interactions between the opportunistic species Ulva prolifera and the native macroalga Gracilaria lichvoides. PLoS ONE 2012, 7, e33648. [Google Scholar] [CrossRef]
- Bieberich, J.; Lauerer, M.; Drachsler, M.; Heinrichs, J.; Müller, S.; Feldhaar, H. Species- and developmental stage-specific effects of allelopathy and competition of invasive Impatiens glandulifera on cooccurring plants. PLoS ONE 2018, 13, e0205843. [Google Scholar] [CrossRef]
- Pan, Z.L.; Yu, Y.Y.; Chen, Y.L.; Yu, C.Z.; Xu, N.J.; Li, Y.H. Combined effects of biomass density and low-nighttime temperature on the competition for growth and physiological performance of Gracilariopsis lemaneiformis and Ulva prolifera. Algal Res. 2022, 62, 102638. [Google Scholar] [CrossRef]
- Bais, H.P.; Venkatachalam, L.; Biedrzycki, M.L. Stimulation or inhibition: Conflicting evidence for (±)-catechin’s role as a chemical facilitator and disease protecting agent. Plant Signal. Behav. 2010, 5, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.J.; Huang, G.H.; Fu, H.Y.; An, C.J.; Yao, Y.; Cheng, G.H.; Suo, M.Q. Allelopathy inhibitory effects of Hydrodictyon reticulatum on Chlorella pyrenoidosa under co-culture and liquor-cultured conditions. Water 2017, 9, 416. [Google Scholar] [CrossRef]
- Belz, R.G. Stimulation versus inhibition–bioactivity of parthenin, a phytochemical from Parthenium hysterophorus L. Dose-Response 2008, 6, 80–96. [Google Scholar] [CrossRef]
- Belz, R.G. Investigating a potential auxin–related mode of hermetic/inhibitory action of the phytotoxin parthenin. J. Chem. Ecol. 2016, 42, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Budzałek, G.; Śliwińska-Wilczewska, S.; Wiśniewska, K.; Wochna, A.; Bubak, I.; Latała, A.; Wiktor, J.M. Macroalgal defense against competitors and herbivores. Int. J. Mol. Sci. 2021, 22, 7865. [Google Scholar] [CrossRef]
- Xie, Z.H.; Xiao, H.; Cai, H.J.; Wang, R.J.; Tang, X.X. Influence of UV-B irradiation on the interspecific growth interaction between Heterosigma akashiwo and Prorocentrum donghaiense. Int. Rev. Hydrobiol. 2006, 91, 555–573. [Google Scholar] [CrossRef]
- Nagayama, K.; Shibata, T.; Fujimoto, K.; Honjo, T.; Nakamura, T. Algicidal effect of phlorotannins from the brown alga Ecklonia kurome on red tide microalgae. Aquaculture 2003, 218, 601–611. [Google Scholar] [CrossRef]
- Accoroni, S.; Percopo, I.; Cerino, F.; Romagnoli, T.; Pichierri, S.; Perrone, C.; Totti, C. Allelopathic interactions between the HAB dinoflagellate Ostreopsis cf. ovata and macroalgae. Harmful Algae 2015, 49, 147–155. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, B.Y.; Ge, F.J.; Liu, Q.; Zhang, Y.Y.; Zhou, Q.H.; Xu, D.; Wu, Z.B. Interspecific competition for nutrients between submerged macrophytes (Vallisneria natans, Ceratophyllum demersum) and filamentous green algae (Cladophora oligoclona) in a co-culture system. Pol. J. Environ. Stud. 2019, 28, 1483–1494. [Google Scholar] [CrossRef]
- Zhang, S.D.; Yu, Z.M.; Song, X.X.; Song, F.; Wang, Y. Competition about nutrients between Gracilaria lemaneiformis and Prorocentrum donghaiense. Acta Ecol. Sin. 2005, 25, 2676–2680, (In Chinese with English abstract). [Google Scholar]
- Liu, Z.Y.; Sun, P.; Qin, S.; Li, J.J.; Zhuang, L.C.; Song, W.L.; Bi, Y.X.; Zhong, Z.H. Comparative analysis of growth and photosynthetic characteristics in benthic and floating Sargassum horneri. Chin. J. Ecol. 2021, 40, 76–83, (In Chinese with English abstract). [Google Scholar]
- Chen, B.B.; Zou, D.H.; Jiang, H. Elevated CO2 exacerbates competition for growth and photosynthesis between Gracilaria lemaneiformis and Ulva lactuca. Aquaculture 2015, 443, 49–55. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, B.; Tang, X. Effects of two species of macroalgae—Ulva pertusa and Gracilaria lemaneiformis—On growth of Heterosigma akashiwo (Raphidophyceae). J. Appl. Phycol. 2009, 21, 375–385. [Google Scholar] [CrossRef]
- Lu, H.M.; Xie, H.H.; Gong, Y.X.; Wang, Q.; Yang, Y.F. Secondary metabolites from the seaweed Gracilaria lemaneiformis and their allelopathic effects on Skeletonema costatum. Biochem. Syst. Ecol. 2011, 39, 397–400. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Gracilariopsis lemaneiformis | Sargassum horneri | ||||
|---|---|---|---|---|---|---|
| df | F | p | df | F | p | |
| RGR | 3 | 112.721 | <0.001 | 3 | 16.595 | <0.001 |
| Net photosynthetic rate | 3 | 29.975 | <0.001 | 3 | 6.836 | <0.05 |
| NO3-N uptake rate | 3 | 52.763 | <0.001 | 3 | 32.004 | <0.001 |
| Chl a | 3 | 4.510 | <0.05 | 3 | 4.157 | <0.05 |
| Soluble protein | 3 | 6.398 | <0.05 | 3 | 11.253 | <0.01 |
| Soluble carbohydrate | 3 | 39.980 | <0.001 | 3 | 33.784 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, H.; Liu, Y.; Li, J.; Gong, Q.; Gao, X. Interactions between Cultivated Gracilariopsis lemaneiformis and Floating Sargassum horneri under Controlled Laboratory Conditions. Water 2022, 14, 2664. https://doi.org/10.3390/w14172664
Song H, Liu Y, Li J, Gong Q, Gao X. Interactions between Cultivated Gracilariopsis lemaneiformis and Floating Sargassum horneri under Controlled Laboratory Conditions. Water. 2022; 14(17):2664. https://doi.org/10.3390/w14172664
Chicago/Turabian StyleSong, Hanmo, Yan Liu, Jingyu Li, Qingli Gong, and Xu Gao. 2022. "Interactions between Cultivated Gracilariopsis lemaneiformis and Floating Sargassum horneri under Controlled Laboratory Conditions" Water 14, no. 17: 2664. https://doi.org/10.3390/w14172664
APA StyleSong, H., Liu, Y., Li, J., Gong, Q., & Gao, X. (2022). Interactions between Cultivated Gracilariopsis lemaneiformis and Floating Sargassum horneri under Controlled Laboratory Conditions. Water, 14(17), 2664. https://doi.org/10.3390/w14172664