
Biogeochemical Budgets of Nutrients and Metabolism in the Curonian Lagoon (South East Baltic Sea): Spatial and Temporal Variations

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
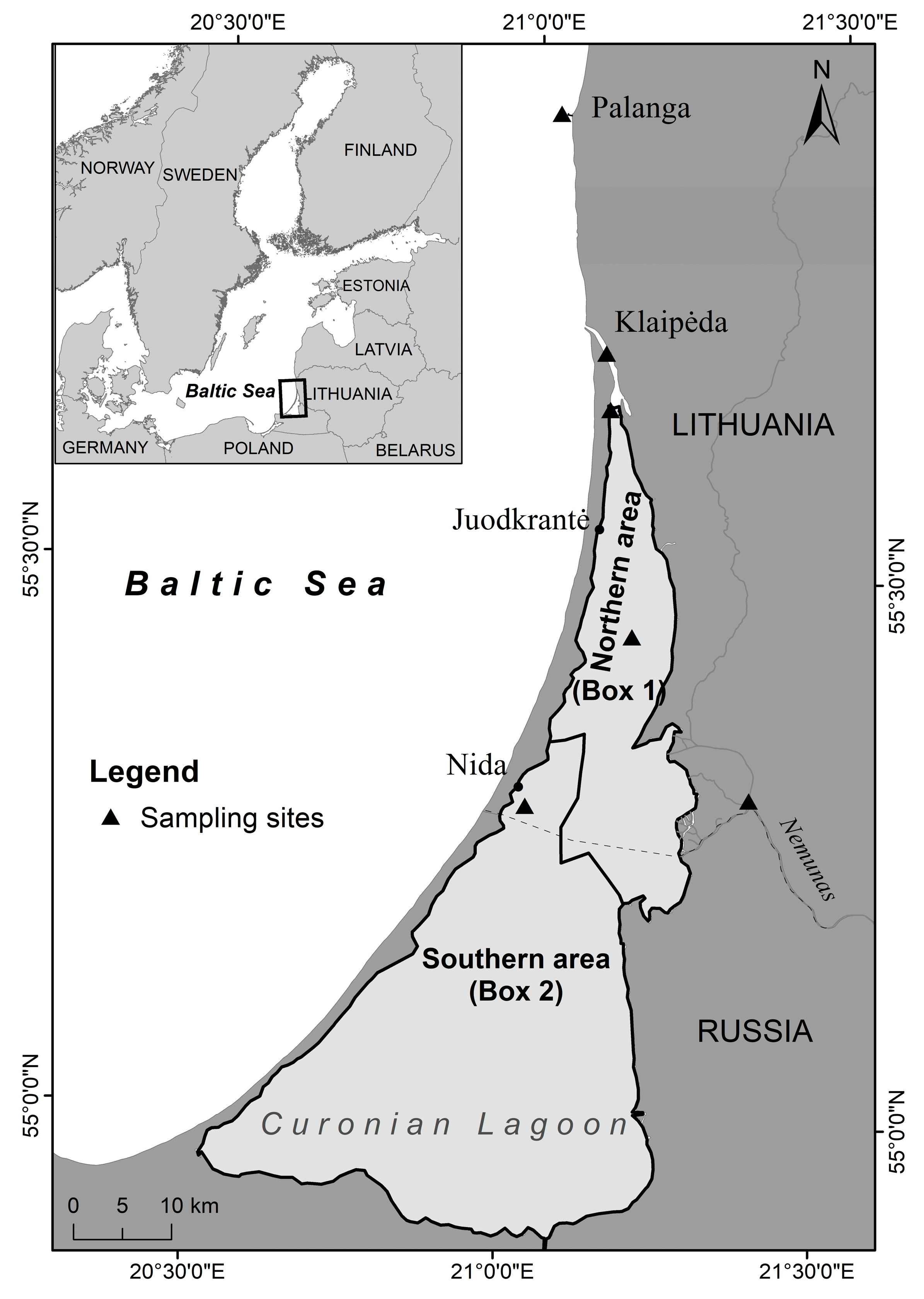
2.1. Study Site
2.2. Water Sampling and Nutrient Analysis
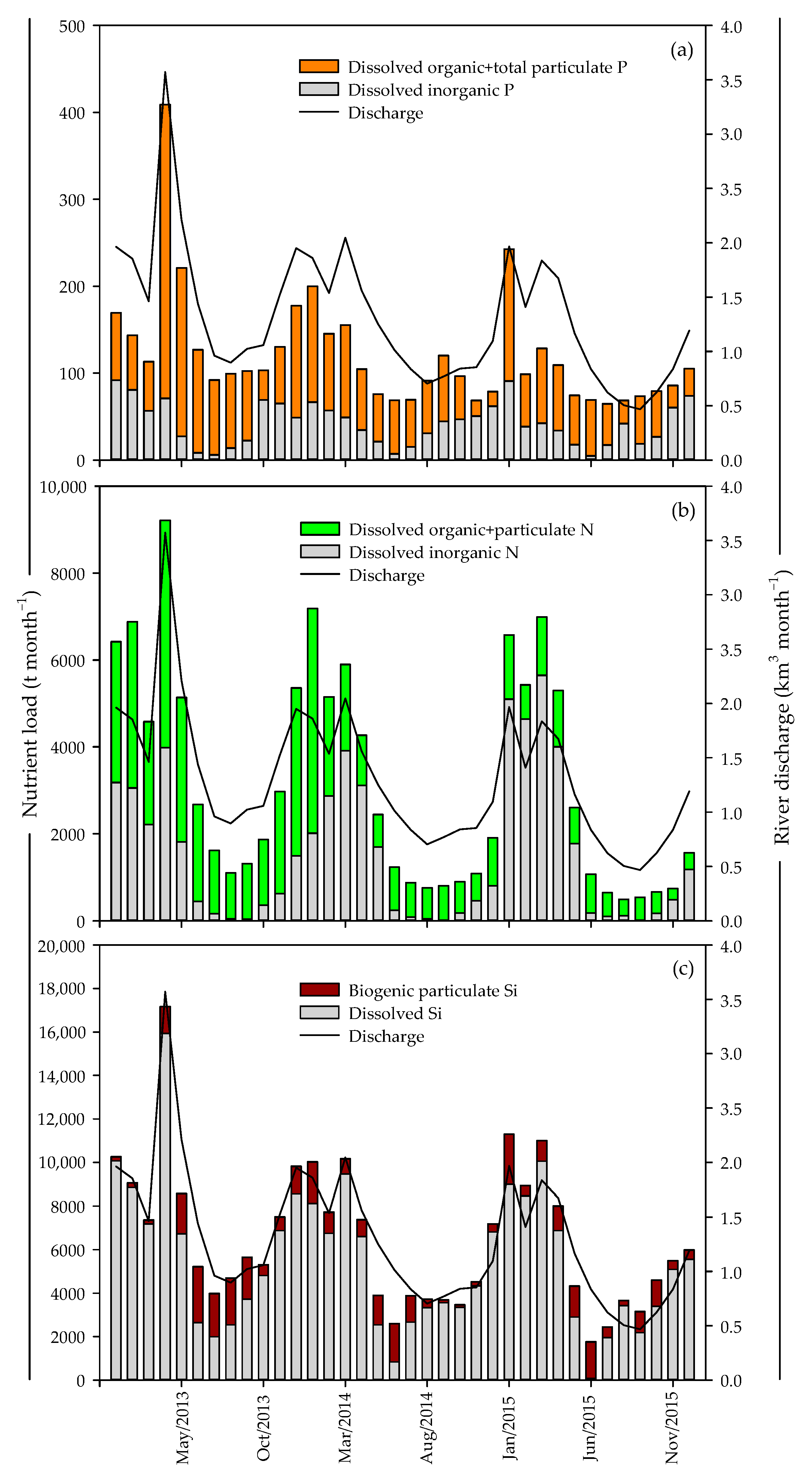
2.3. Discharge Data and Nutrient Input Calculation
2.4. Nutrient Balance and Metabolism Calculations
3. Results
3.1. Water Balance
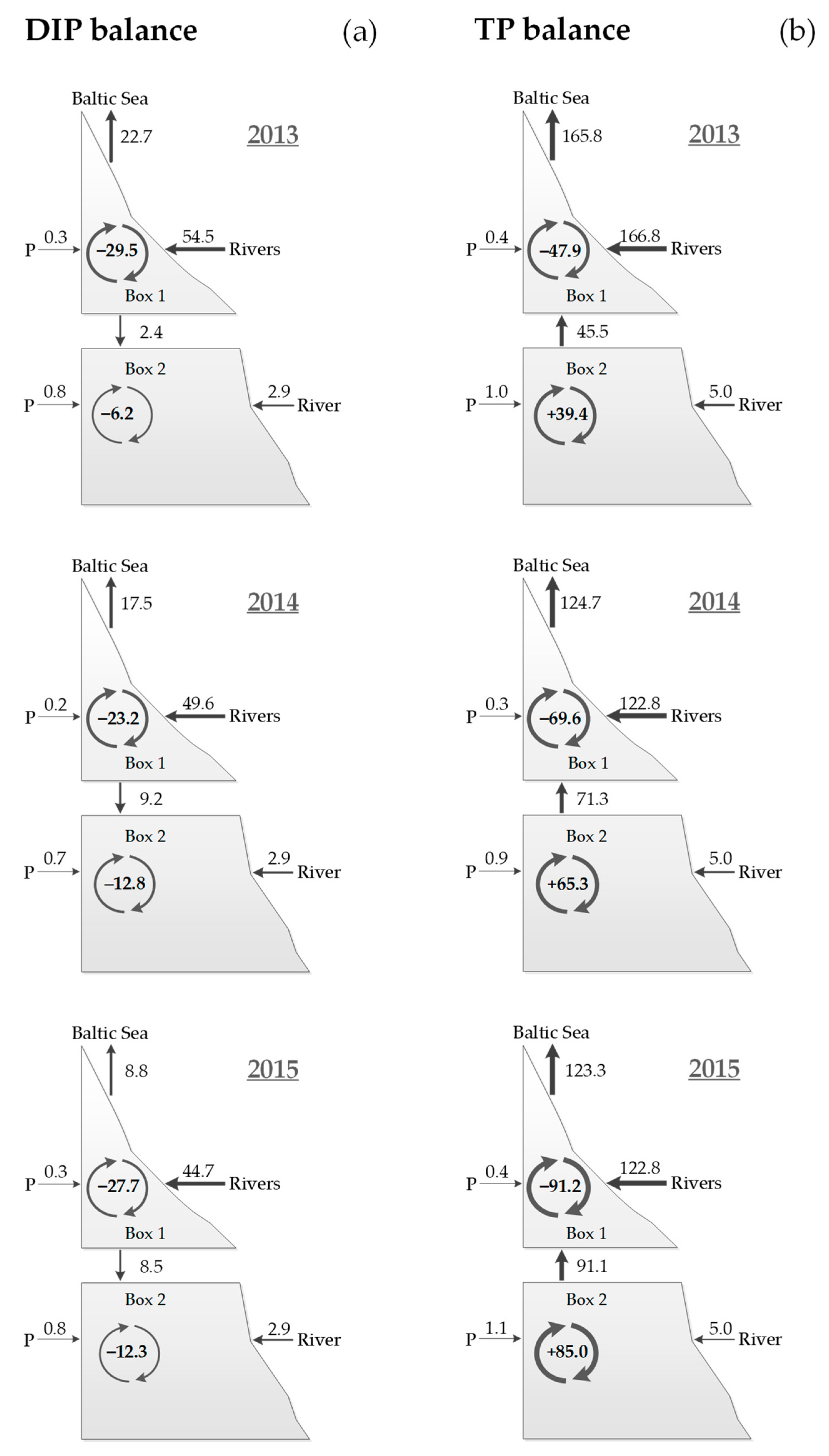
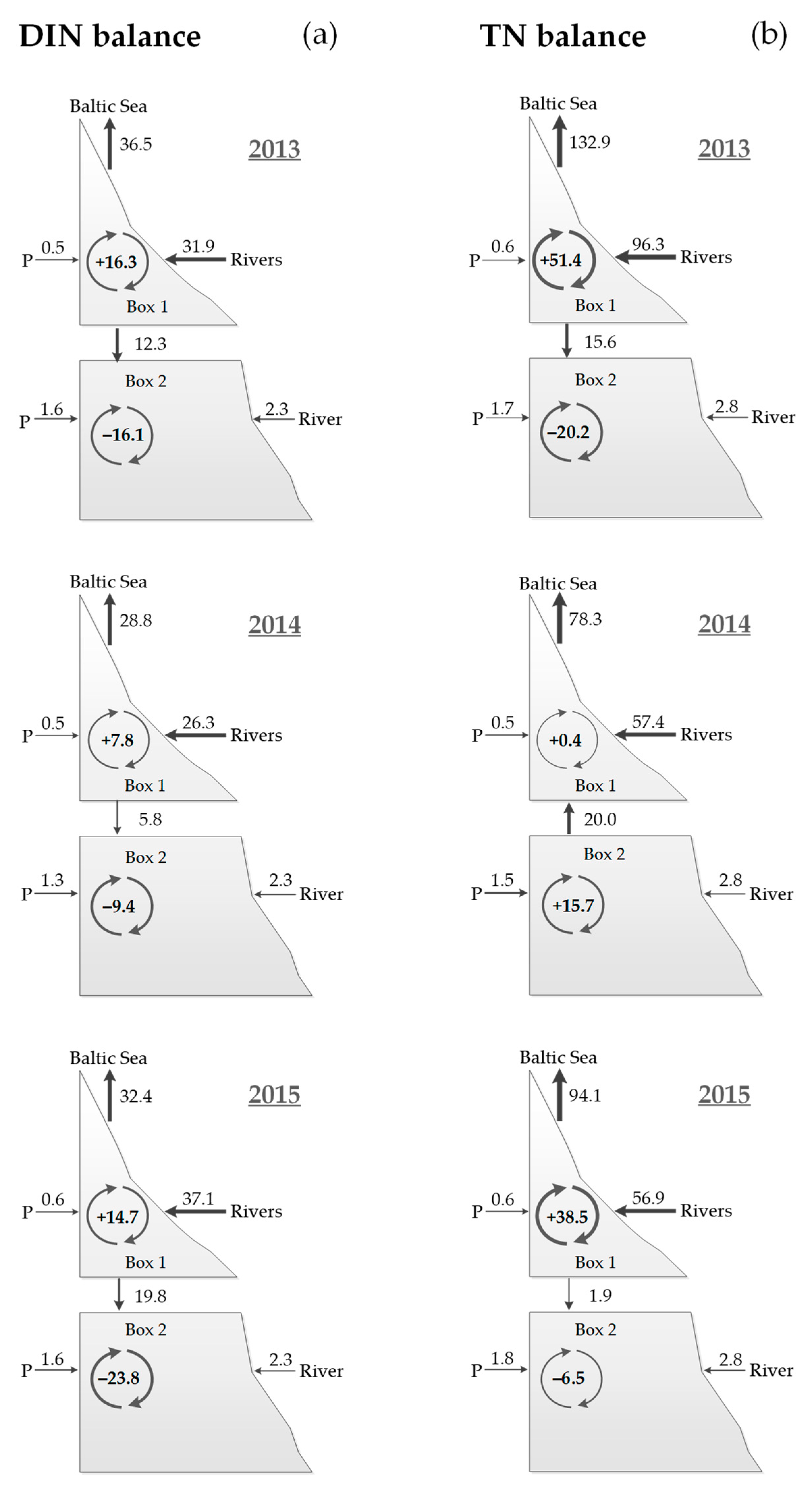
3.2. Nutrient Balances

3.3. Ecosystems Functions
4. Discussion
4.1. Water Balance
4.2. Nutrient Balances
4.3. Ecosystem Functions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gazeau, F.; Smith, S.V.; Gentili, B.; Frankignoulle, M.; Gattuso, J. The European coastal zone: Characterization and first assessment of ecosystem metabolism. Estuar. Coast. Shelf Sci. 2004, 60, 673–694. [Google Scholar] [CrossRef]
- Danielsson, Å.; Papush, L.; Rahm, L. Alterations in nutrient limitations—Scenarios of a changing Baltic Sea. J. Mar. Syst. 2008, 73, 263–283. [Google Scholar] [CrossRef]
- Nowicki, B.L.; Oviatt, C.A. Are estuaries traps for anthropogenic nutrients? Evidence from estuarine mesocosms. Mar. Ecol. Prog. Ser. 1990, 66, 131–146. [Google Scholar] [CrossRef]
- McGuirk, F.A. Organic matter and nutrient cycling in a coastal plain estuary: Carbon, nitrogen and phosphorus distributions, budgets and fluxes. J. Coast. Res. 2008, 10055, 76–94. [Google Scholar] [CrossRef] [Green Version]
- Carstensen, J.; Sánchez-Camacho, M.; Duarte, C.M.; Krause-Jensen, D.; Marba, N. Connecting the dots: Responses of coastal ecosystems to changing nutrient concentrations. Environ. Sci. Technol. 2011, 45, 9122–9132. [Google Scholar] [CrossRef]
- Vybernaite-Lubiene, I.; Zilius, M.; Giordani, G.; Petkuviene, J.; Vaiciute, D.; Bukaveckas, P.A.; Bartoli, M. Effect of algal blooms on retention of N, Si and P in Europe’s largest coastal lagoon. Estuar. Coast. Shelf Sci. 2017, 194, 217–228. [Google Scholar] [CrossRef]
- Zilius, M.; Vybernaite-Lubiene, I.; Vaiciute, D.; Petkuviene, J.; Zemlys, P.; Liskow, I.; Voss, M.; Bartoli, M.; Bukaveckas, P.A. The influence of cyanobacteria blooms on the attenuation of nitrogen throughputs in a Baltic coastal lagoon. Biogeochemistry 2018, 141, 143–165. [Google Scholar] [CrossRef]
- Nixon, S.W.; Ammerman, J.W.; Atkinson, L.P.; Berounsky, V.M.; Billen, G.; Boicourt, W.C.; Boynton, W.R.; Church, T.M.; Ditoro, D.M.; Elmgren, R.; et al. The fate of nitrogen and phosphorus at the land-sea margin of the North Atlantic Ocean. Biogeochemistry 1996, 35, 141–180. [Google Scholar] [CrossRef]
- Boyer, E.W.; Howarth, R.W. Nitrogen fluxes from rivers to the coastal oceans. In Nitrogen in the Marine Environment; Elsevier Inc.: Amsterdam, The Netherlands, 2008; pp. 1565–1587. [Google Scholar]
- Asmala, E.; Carstensen, J.; Conley, D.J.; Slomp, C.P.; Stadmark, J.; Voss, M. Efficiency of the coastal filter: Nitrogen and phosphorus removal in the Baltic Sea. Limnol. Oceanogr. 2017, 62, S222–S238. [Google Scholar] [CrossRef]
- Schubel, J.R.; Kennedy, V.S. Estuary as a Filter: An Introduction; Academic Press: Orlando, FL, USA, 1984; Volume 10, pp. 1–11. [Google Scholar]
- Nedwell, D.B.; Jickells, T.D.; Trimmer, M.; Sanders, R. Nutrients in Estuaries. In Advances in Ecological Research; Nedwell, D.B., Raffaelli, D.G., Eds.; Academic Press: Cambridge, MA, USA, 1999; Volume 29, pp. 43–92. [Google Scholar]
- Sundbäck, K.; Linares FLarson, F.; Wulff, A.; Engelsen, A. Benthic nitrogen fluxes along a depth gradient in a microtidal fjord: The role of denitrification and microphytobenthos. Limnol. Oceanogr. 2004, 49, 1095–1107. [Google Scholar] [CrossRef]
- Brion, N.; Jans, S.; Chou, L.; Rousseau, V. Nutrient loads to the Belgian coastal zone. In Current Status of Eutrophication in the Belgian Coastal Zone; Presse Universitaire de Bruxelles: Brussels, Belgium, 2008. [Google Scholar]
- Eyre, B.D.; Maher, D.T.; Sanders, C. The contribution of denitrification and burial to the nitrogen budgets of three geomorphically distinct Australian estuaries: Importance of seagrass habitats. Limnol. Oceanogr. 2016, 61, 1144–1156. [Google Scholar] [CrossRef] [Green Version]
- Jensen, H.S.; Mortensen, P.B.; Andersen, F.O.; Rasmussen, E.; Jensen, A. Phosphorus cycling in a coastal marine sediment, Aarhus Bay, Denmark. Limnol. Oceanogr. 1995, 40, 908–917. [Google Scholar] [CrossRef]
- Howarth, R.W.; Billen, G.; Swaney, D.; Townsend, A.; Jawoski, N.; Lajtha, K.; Downing, J.A.; Elmgren, R.; Caraco, N.; Jordan, T.; et al. Regional nitrogen budgets and riverine N & P fluxes for the drainages to the North Atlantic Ocean: Natural and human influences. Biogeochemistry 1996, 35, 75–139. [Google Scholar]
- Bukaveckas, P.A.; Isenberg, W.N. Loading, transformation, and retention of nitrogen and phosphorus in the tidal freshwater James River (Virginia). Estuaries Coasts 2013, 36, 1219–1236. [Google Scholar] [CrossRef]
- Viaroli, P.; Bartoli, M.; Giordani, G.; Naldi, M.; Orfanidis, S.; Zaldivar, J.M. Community shifts, alternative stable states, biogeochemical controls and feedbacks in eutrophic coastal lagoons: A brief overview. Aquat. Conserv. Mar. Freshw. Ecosyst. 2008, 18, S105–S117. [Google Scholar] [CrossRef]
- Zilius, M.; Bartoli, M.; Bresciani, M.; Katarzyte, M.; Ruginis, T.; Petkuviene, J.; Lubiene, I.; Giardino, C.; Bukaveckas, P.A.; de Wit, R.; et al. Feedback mechanisms between cyanobacterial blooms, transient hypoxia, and benthic phosphorus regeneration in shallow coastal environments. Estuaries Coasts 2014, 37, 680–694. [Google Scholar] [CrossRef]
- Nixon, S.W.; Hunt, C.D.; Nowicki, B.L. The retention of nutrients (C, N, P), heavy metals (Mn. Cd, Pb, Cu), and petroleum hydrocarbons in Narragansett Bay. In Biogeochemical Processes at the Land-Sea Boundary; Elsevier: Amsterdam, The Netherlands, 1986; pp. 99–122. [Google Scholar]
- Nixon, S.W. Chesapeake Bay nutrient budgets: A reassessment. Biogeochemistry 1987, 4, 77–90. [Google Scholar] [CrossRef]
- Petkuviene, J.; Zilius, M.; Lubiene, I.; Ruginis, T.; Giordani, G.; Razinkovas-Baziukas, A.; Bartoli, M. Phosphorus cycling in a freshwater estuary impacted by cyanobacterial blooms. Estuaries Coasts 2016, 39, 1386–1402. [Google Scholar] [CrossRef]
- Herbert, R.A. Nitrogen cycling in coastal marine ecosystems. FEMS Microbiol. Rev. 1999, 23, 563–590. [Google Scholar] [CrossRef]
- Le Tissier, M.; Buddemeier, R.; Parslow, J.; Swaney, D.P.; Crossland, C.J.; Smith, S.V.; Kremer, H.H. The Role of the Coastal Ocean in the Disturbed and Undisturbed Nutrient and Carbon Cycles—A Management Perspective; LOICZ: Geesthacht, Germany, 2006; p. 44. [Google Scholar]
- Boynton, W.R.; Hagy, J.D.; Cornwel, J.C.; Kemp, W.M.; Greene, S.M.; Owens, M.S.; Baker, J.E.; Larsen, R.K. Nutrient budgets and management actions in the Patuxent river estuary, Maryland. Estuaries Coasts 2008, 31, 623–651. [Google Scholar] [CrossRef]
- Gordon, D.C.; Boudreau, P.R.; Mann, K.H.; Ong, J.E.; Silvert, W.L.; Smith, S.V.; Wattayakorn, G.; Wulff, F.; Yanagi, T. LOICZ Biogeochemical Modelling Guidelines; LOICZ: Texel, The Netherlands, 1996. [Google Scholar]
- Ramesh, R.; Chen, Z.; Cummins, V.; Day, J.; D’Elia, C.; Dennison, B.; Forbes, D.L.; Glaeser, B.; Glaser, M.; Glavovic, B.; et al. Land–Ocean Interactions in the Coastal Zone: Past, present & future. Anthr. Rev. 2015, 12, 85–98. [Google Scholar]
- Swaney, D.P.; Giordani, G. LOICZ Research & Studies No. 37. Helmholtz-Zentrum Geesthacht. In Proceedings of the LOICZ Workshop on Biogeochemical Budget Methodology and Applications, Providence, RI, USA, 9–10 November 2007; p. 195. [Google Scholar]
- Rabalais, N.N. Nitrogen in Aquatic Ecosystems. Ambio 2002, 102–112. [Google Scholar] [CrossRef]
- Borja, Á.; Elliott, M.; Carstensen, J.; Heiskanen, A.S.; van de Bund, W. Marine management–towards an integrated implementation of the European Marine Strategy Framework and the Water Framework Directives. Mar. Pollut. Bull. 2010, 60, 2175–2186. [Google Scholar] [CrossRef]
- Voss, M.; Dippner, J.; Humborg, C.; Korth, F.; Neumann, T.; Hürdler, J.; Schernewski, G.; Venohr, M. History and scenarios of future development of Baltic Sea eutrophication. Estuar. Coast. Shelf Sci. 2011, 92, 307–322. [Google Scholar] [CrossRef]
- HELCOM. HELCOM Guidelines for the Annual and Periodical Compilation and Reporting of Waterborne Pollution Inputs to the Baltic Sea (PLC-Water). 2015. Available online: http://www.helcom.fi/Lists/Publications/PLC-Water%20Guidelines.pdf (accessed on 8 November 2018).
- Vybernaite-Lubiene, I.; Zilius, M.; Saltyte-Vaisiauske, L.; Bartoli, M. Recent Trends (2012–2016) of N, Si, and P Export from the Nemunas River Watershed: Loads, Unbalanced Stoichiometry, and Threats for Downstream Aquatic Ecosystems. Water 2018, 10, 1178. [Google Scholar] [CrossRef] [Green Version]
- Zemlys, P.; Ferrarin, C.; Umgiesser, G.; Gulbinskas, S.; Bellafiore, D. Investigation of saline water intrusions into the Curonian Lagoon (Lithuania) and two-layer flow in the Klaipėda Strait using finite element hydrodynamic model. Ocean Sci. J. 2013, 9, 573–584. [Google Scholar] [CrossRef] [Green Version]
- Jakimavičius, D.; Kovalenkovienė, M. Long-term water balance of the Curonian Lagoon in the context of anthropogenic factors and climate change. Baltica 2010, 23, 33–46. [Google Scholar]
- Vaitkuviene, D.; Dagys, M. CORINE 2006 Land Cover, Lithuania. Implementing CLC2006 Project in Lithuania, Report; Institute of Ecology of Vilnius University: Vilnius, Lithuania, 2008. [Google Scholar]
- Umgiesser, G.; Zemlys, P.; Erturk, A.; Razinkova-Baziukas, A.; Mežinė, J.; Ferrarin, C. Seasonal renewal time variability in the Curonian Lagoon caused by atmospheric and hydrographical forcing. Ocean Sci. J. 2016, 12, 391–402. [Google Scholar] [CrossRef] [Green Version]
- Pilkaitytė, R.; Razinkovas, A. Factors controlling phytoplankton blooms in a temperate estuary: Nutrient limitation and physical forcing. Hydrobiologia 2006, 555, 41–48. [Google Scholar] [CrossRef]
- Jakimavicius, D. Changes of Water Balance Elements of the Curonian Lagoon and Their Forecast Due to Anthropogenic and Natural Factors. Ph.D. Thesis, Lithuanian Energy Institute, Kaunas, Lithuania, 2012. [Google Scholar]
- Caffrey, J.M. Production, respiration and net ecosystem metabolism in US estuaries. In Coastal Monitoring through Partnerships; Springer: Dordrecht, The Netherlands, 2003; pp. 207–219. [Google Scholar]
- Ferrarin, C.; Razinkovas, A.; Gulbinskas, S.; Umgiesser, G.; Bliudziute, L. Hydraulic regime-based zonation scheme of the Curonian Lagoon. Hydrobiologia 2008, 611, 133–146. [Google Scholar] [CrossRef] [Green Version]
- Jakimavičius, D.; Kriaučiūnienė, J. The climate change impact on the water balance of the Curonian Lagoon. Water Resour. Res. 2013, 40, 120–132. [Google Scholar] [CrossRef]
- Bresciani, M.; Adamo, M.; De Carolis, G.; Matta, E.; Pasquariello, G.; Vaičiūtė, D.; Giardino, C. Monitoring blooms and surface accumulation of cyanobacteria in the Curonian Lagoon by combining MERIS and ASAR data. Remote Sens. Environ. 2014, 146, 124–135. [Google Scholar] [CrossRef]
- Fulweiler, R.W.; Nixon, S.W. Export of nitrogen, phosphorus, and suspended solids from a southern New England watershed to Little Narragansett Bay. Biogeochemistry 2005, 76, 567–593. [Google Scholar] [CrossRef]
- Korth, F.; Fry, B.; Liskow, I.; Voss, M. Nitrogen turnover during the spring outflows of the nitrate-rich Curonian and Szczecin lagoons using dual nitrate isotopes. Mar. Chem. 2013, 154, 1–11. [Google Scholar] [CrossRef]
- Bauer, A. Status of Nutrient Bookkeeping in the Baltic Sea Countries. Report document Texte 95/2015. 2015. Available online: http://www.umweltbundesamt.de/publikationen/status-of-nutrient-bookkeeping-in-the-baltic-sea.pdf (accessed on 13 December 2021).
- Mangalaa, K.R.; Cardinal, D.; Brajard, J.; Rao, D.B.; Sarma, N.S.; Djouraev, I.; Chiranjeevulu, G.; Murty, K.N.; Sarma, V.V.S.S. Silicon cycle in Indian estuaries and its control by biogeochemical and anthropogenic processes. Cont. Shelf Res. 2017, 148, 64–88. [Google Scholar] [CrossRef]
- Rendell, A.R.; Horrobin, T.M.; Jickells, T.D.; Edmunds, H.M.; Brown, J.; Malcolm, S.J. Nutrient cycling in the Great Ouse estuary and its impact on nutrient fluxes to The Wash, England. Estuar. Coast. Shelf Sci. 1997, 45, 653–668. [Google Scholar] [CrossRef]
- Grelowski, A.; Pastuszak, M.; Sitek, S.; Witek, Z. Budget calculations of nitrogen, phosphorus and BOD 5 passing through the Oder estuary. J. Mar. Syst. 2000, 5, 221–237. [Google Scholar] [CrossRef]
- Bukaveckas, P.A.; Beck, M.; Devore, D.; Lee, W.M. Climatic variability and its role in regulating C, N and P retention in the James River Estuary. Estuar. Coast. Shelf Sci. 2018, 205, 161–173. [Google Scholar] [CrossRef] [Green Version]
- Edman, M.K.; Eilola, K.; Almroth-Rosell, E.; Meier, H.E.; Wåhlström, I.; Arneborg, L. Nutrient retention in the Swedish coastal zone. Front. Mar. Sci. 2018, 5, 415. [Google Scholar] [CrossRef] [Green Version]
- Amann, T.; Weiss, A.; Hartmann, J. Silica fluxes in the inner Elbe Estuary, Germany. Biogeochemistry 2014, 118, 389–412. [Google Scholar] [CrossRef]
- Soetaert, K.; Middelburg, J.J.; Heip, C.; Meire, P.; Van Damme, S.; Maris, T. Long-term change in dissolved inorganic nutrients in the heterotrophic Scheldt estuary (Belgium, The Netherlands). Limnol. Oceanogr. 2006, 51, 409–423. [Google Scholar] [CrossRef] [Green Version]
- Romero, E.; Garnier, J.; Billen, G.; Ramarson, A.; Riou, P.; Le Gendre, R. Modeling the biogeochemical functioning of the Seine estuary and its coastal zone: Export, retention, and transformations. Limnol. Oceanogr. 2019, 64, 895–912. [Google Scholar] [CrossRef]
- Radtke, H.; Neumann, T.; Voss, M.; Fennel, W. Modeling pathways of riverine nitrogen and phosphorus in the Baltic Sea. J. Geophys. Res. 2012, 117. [Google Scholar] [CrossRef]
- Bartoli, M.; Zilius, M.; Bresciani, M.; Vaičiūtė, D.; Lubiene-Vybernaite, I.; Petkuviene, J.; Giordani, G.; Daunys, D.; Ruginis, T.; Benelli, S.; et al. Drivers of cyanobacterial blooms in a hypertrophic lagoon. Front. Mar. Sci. 2018, 5, 434. [Google Scholar] [CrossRef] [Green Version]
- Crossland, C.J.; Kremer, H.H.; Lindeboom, H.; Crossland, J.I.M.; Le Tissier, M. (Eds.) Coastal Fluxes in the Anthropocene: The Land-Ocean Interactions in the Coastal Zone Project of the International Geosphere-Biosphere Programme; Springer Science & Business Media: Berlin, Germany, 2005. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Macro Area | Year | River Discharge (106 m3 d−1) | Net Outflow (106 m3 d−1) | Exchange Flow (106 m3 d−1) | τw (Days) |
|---|---|---|---|---|---|
| Box 1 | 2013 | 57.8 | 61.9 | 46.3 (Box 1–Sea) | 4 |
| 2014 | 41.5 | 43.8 | 40.2 (Box 1–Sea) | 4 | |
| 2015 | 39.2 | 41.7 | 49.7 (Box 1–Sea) | 4 | |
| Box 2 | 2013 | 4.0 | 4.1 | 85.6 (Box 1–Box 2) | 57 |
| 2014 | 2.3 | 2.2 | 76.8 (Box 1–Box 2) | 65 | |
| 2015 | 2.3 | 2.3 | 90.5 (Box 1–Box 2) | 55 |
| Nutrient | Nutrient Form | Year | Box 1 | Box 2 | Whole Lagoon |
|---|---|---|---|---|---|
| Phosphorus | DIP | 2013 | −29.5 (−0.073) | −6.2 (−0.005) | −35.7 (−0.023) |
| 2014 | −23.2 (−0.057) | −12.8 (−0.011) | −36.0 (−0.023) | ||
| 2015 | −27.7 (−0.069) | −12.3 (−0.010) | −40.0 (−0.025) | ||
| TP | 2013 | −47.9 (−0.118) | +39.4 (+0.033) | −8.5 (−0.005) | |
| 2014 | −69.6 (−0.172) | +65.3 (+0.056) | −4.3 (0.003) | ||
| 2015 | −91.2 (−0.226) | +85.0 (+0.072) | −6.2 (0.004) | ||
| Nitrogen | DIN | 2013 | +16.3 (+4.031) | −16.1 (−1.369) | +0.2 (+0.013) |
| 2014 | +7.8 (+1.929) | −9.4 (−0.799) | −1.6 (−0.101) | ||
| 2015 | +14.7 (+3.635) | −23.8 (−2.023) | −9.1 (−0.576) | ||
| TN | 2013 | +51.4 (+12.711) | −20.2 (−1.717) | +31.2 (+1.974) | |
| 2014 | +0.4 (+0.099) | +15.7 (+1.335) | +16.1 (+1.019) | ||
| 2015 | +38.5 (+9.521) | −6.5 (−0.553) | +32.0 (+2.024) | ||
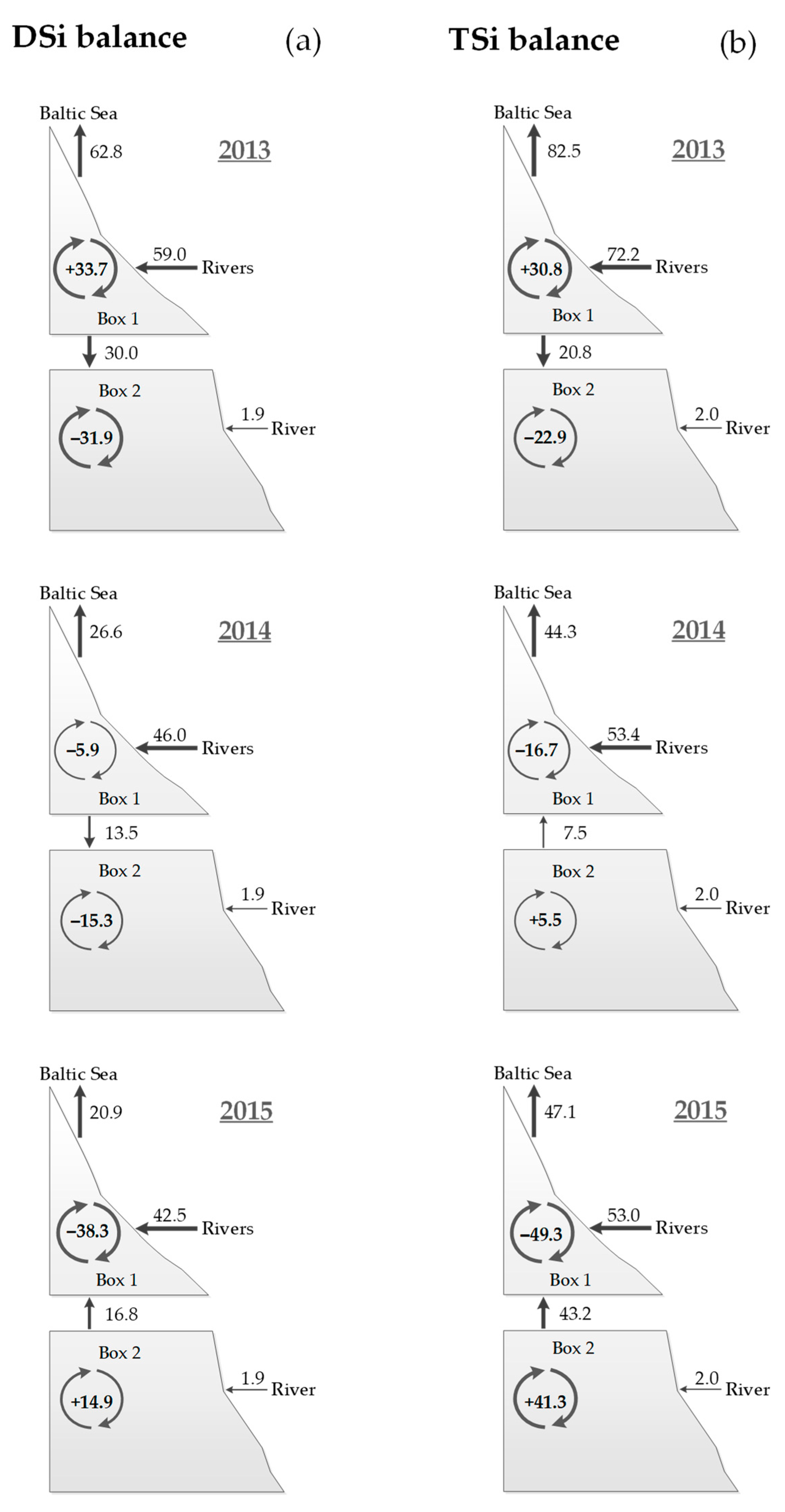
| Silica | DSi | 2013 | +33.7 (+8,334) | −31.9 (−2.712) | +1.8 (+0.114) |
| 2014 | −5.9 (−1.459) | −15.3 (−1.301) | −21.2 (−1.341) | ||
| 2015 | −38.3 (−9.472) | +14.9 (+1.267) | −23.4 (−1.480) | ||
| TSi | 2013 | +30.8 (+7.617) | −22.9 (−1.947) | +7.9 (+0.500) | |
| 2014 | −16.7 (−4.130) | +5.5 (+0.468) | −11.1 (−0.702) | ||
| 2015 | −49.3 (−12.192) | +41.3 (+3.511) | −8.0 (−0.506) |
| Metabolism | Year | Box 1 | Box 2 | Whole Lagoon |
|---|---|---|---|---|
| NEM 105 mol d−1 (mmol C m−2 d−1) | 2013 | 31.3 (7.73) | 6.5 (0.56) | 37.8 (2.39) |
| 2014 | 24.6 (6.08) | 13.5 (1.15) | 38.1 (2.41) | |
| 2015 | 29.3 (7.26) | 13.0 (1.11) | 42.4 (2.68) | |
| (nfix-denit) 105 mol d−1 (mmol N m−2 d−1) | 2013 | 21.0 (5.20) | −15.1 (−1.29) | 5.9 (0.37) |
| 2014 | 11.5 (2.85) | −7.4 (−0.63) | 4.2 (0.26) | |
| 2015 | 19.1 (4.72) | −21.8 (−1.85) | −2.7 (−0.17) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vybernaite-Lubiene, I.; Zilius, M.; Bartoli, M.; Petkuviene, J.; Zemlys, P.; Magri, M.; Giordani, G. Biogeochemical Budgets of Nutrients and Metabolism in the Curonian Lagoon (South East Baltic Sea): Spatial and Temporal Variations. Water 2022, 14, 164. https://doi.org/10.3390/w14020164
Vybernaite-Lubiene I, Zilius M, Bartoli M, Petkuviene J, Zemlys P, Magri M, Giordani G. Biogeochemical Budgets of Nutrients and Metabolism in the Curonian Lagoon (South East Baltic Sea): Spatial and Temporal Variations. Water. 2022; 14(2):164. https://doi.org/10.3390/w14020164
Chicago/Turabian StyleVybernaite-Lubiene, Irma, Mindaugas Zilius, Marco Bartoli, Jolita Petkuviene, Petras Zemlys, Monia Magri, and Gianmarco Giordani. 2022. "Biogeochemical Budgets of Nutrients and Metabolism in the Curonian Lagoon (South East Baltic Sea): Spatial and Temporal Variations" Water 14, no. 2: 164. https://doi.org/10.3390/w14020164
APA StyleVybernaite-Lubiene, I., Zilius, M., Bartoli, M., Petkuviene, J., Zemlys, P., Magri, M., & Giordani, G. (2022). Biogeochemical Budgets of Nutrients and Metabolism in the Curonian Lagoon (South East Baltic Sea): Spatial and Temporal Variations. Water, 14(2), 164. https://doi.org/10.3390/w14020164