
Contribution of Small Phytoplankton to Primary Production in the Northern Bering and Chukchi Seas

 , ,
, ,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
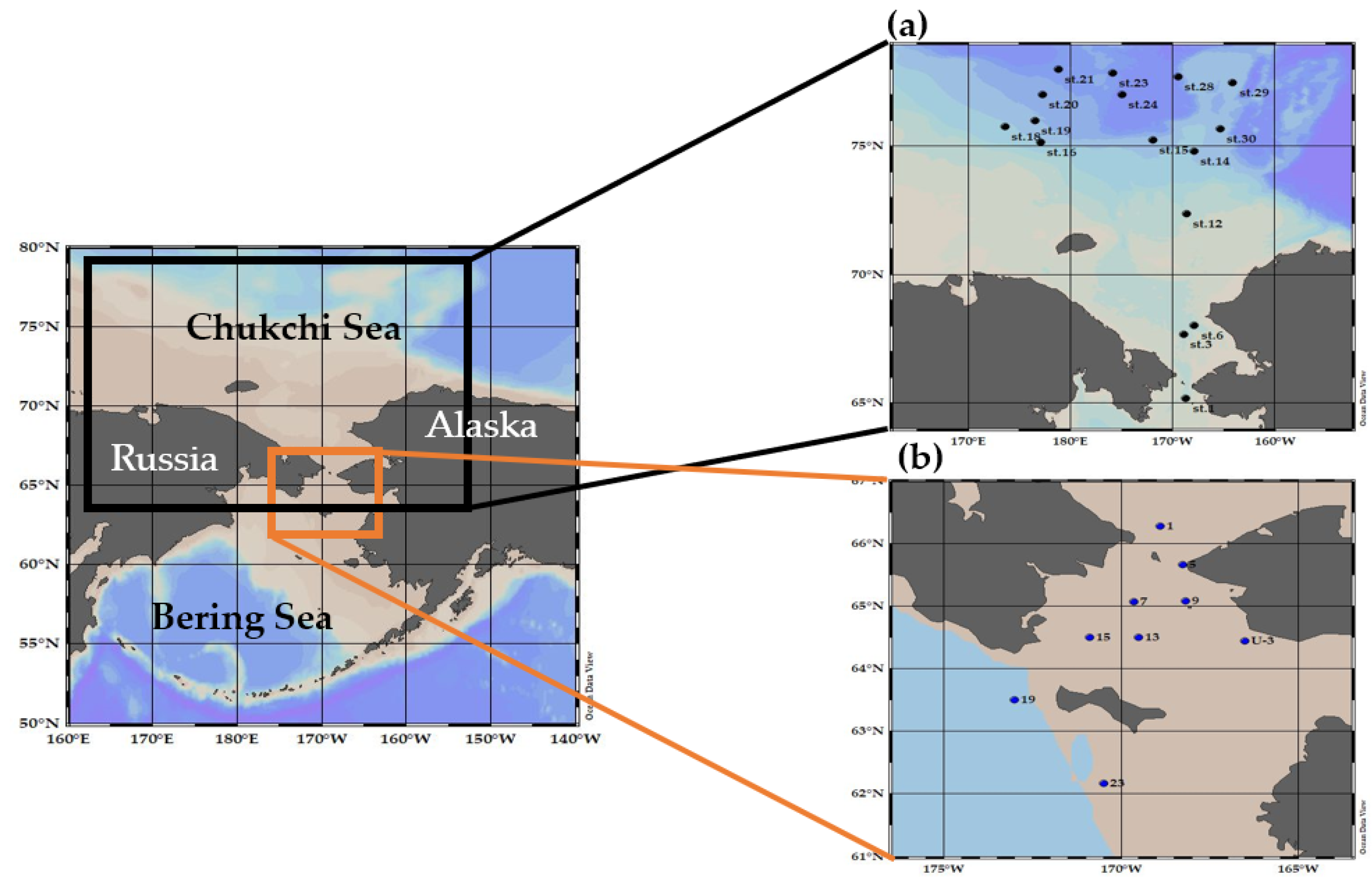
2.1. Study Area and Water Sampling
2.2. Chlorophyll-a Analysis
2.3. High-Performance Liquid Chromatography Analysis for Accessory Pigment Concentration
2.4. Particulate Organic Carbon and Primary Productivity
2.5. Statistical Analysis
3. Results and Discussion
3.1. Spatial Distribution of Temperature and Salinity
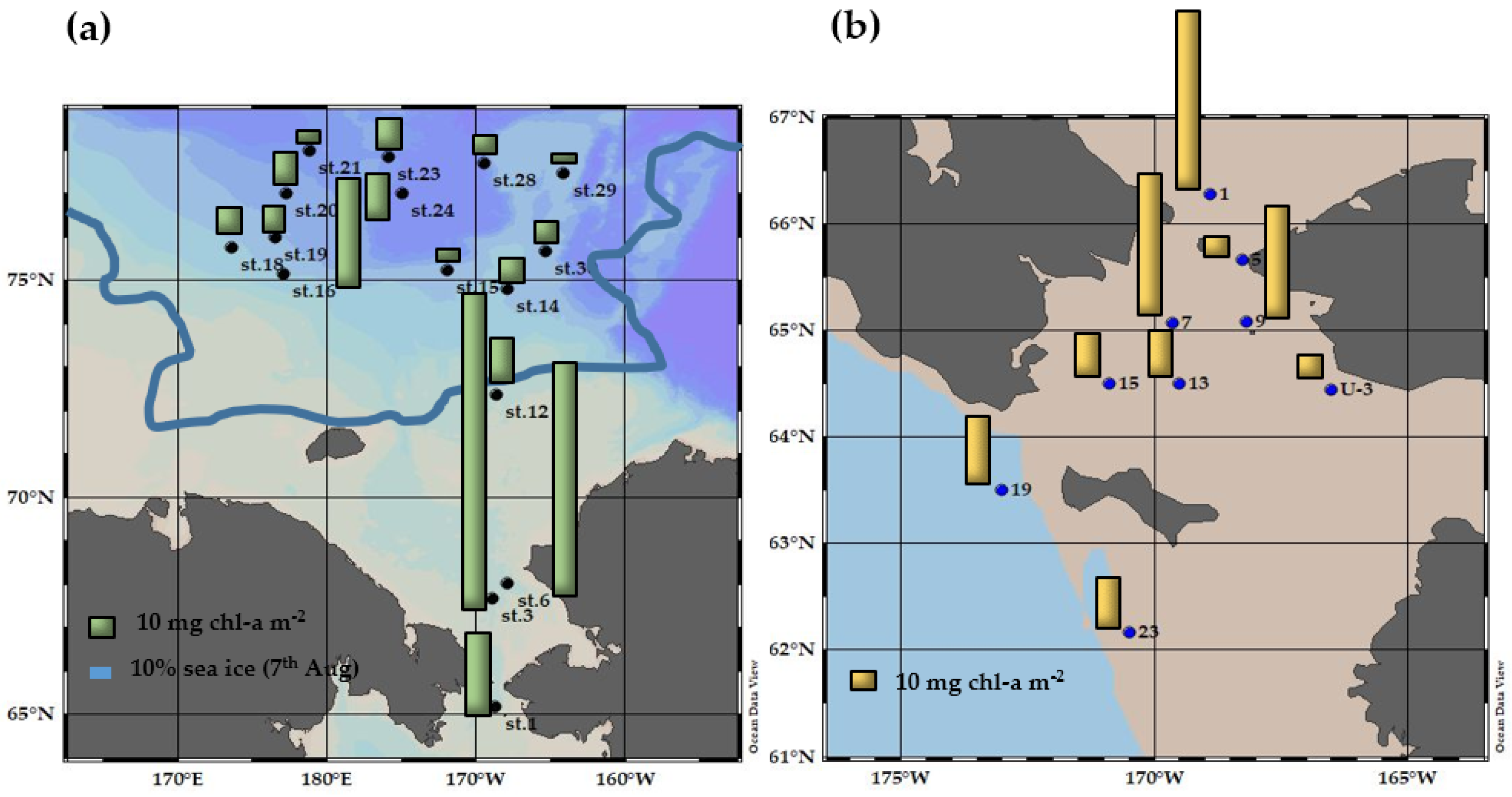
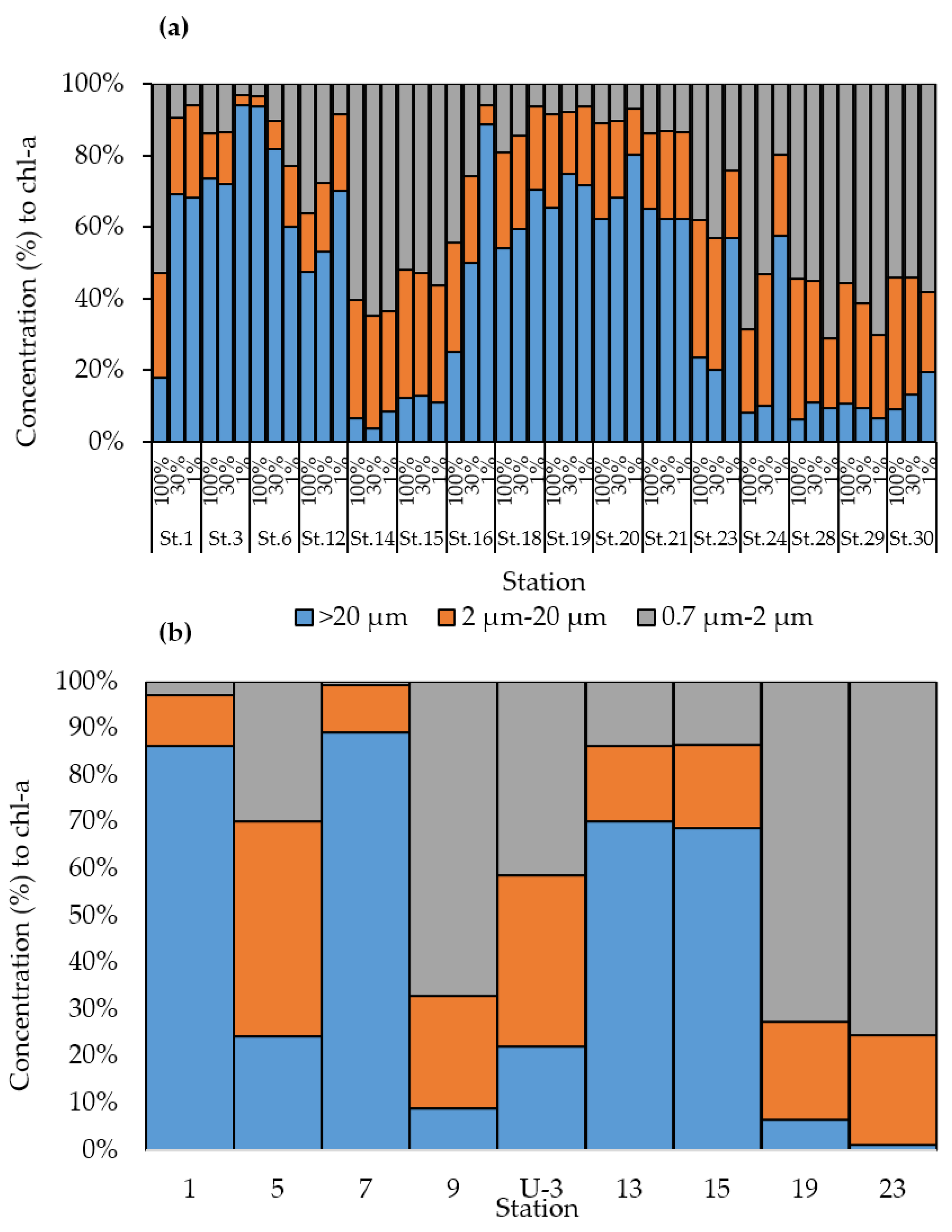
3.2. Chlorophyll-a Concentration and Different Size Chlorophyll-a Compositions in the Northern Bering and Chukchi Seas
3.3. Pigment Composition and Major Dominant Phytoplankton Groups
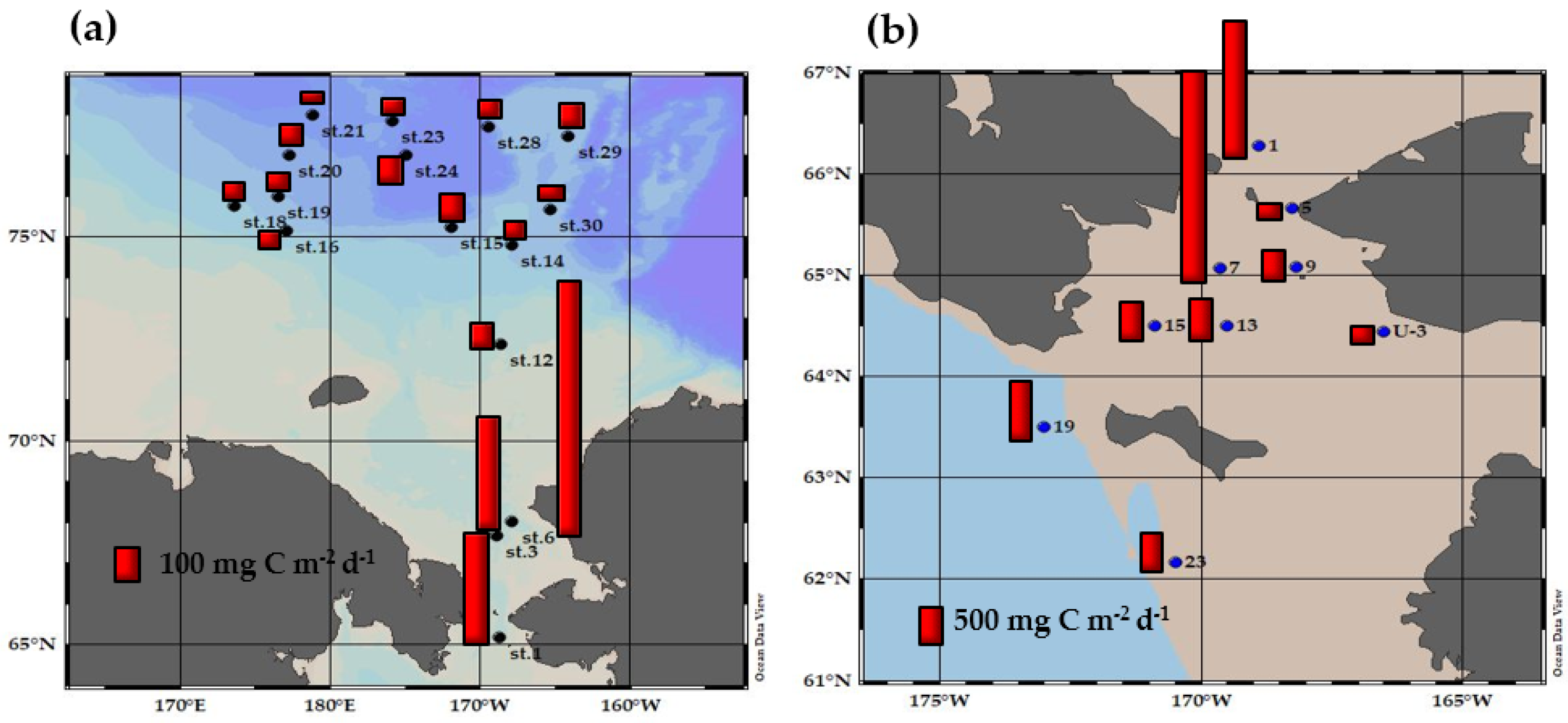
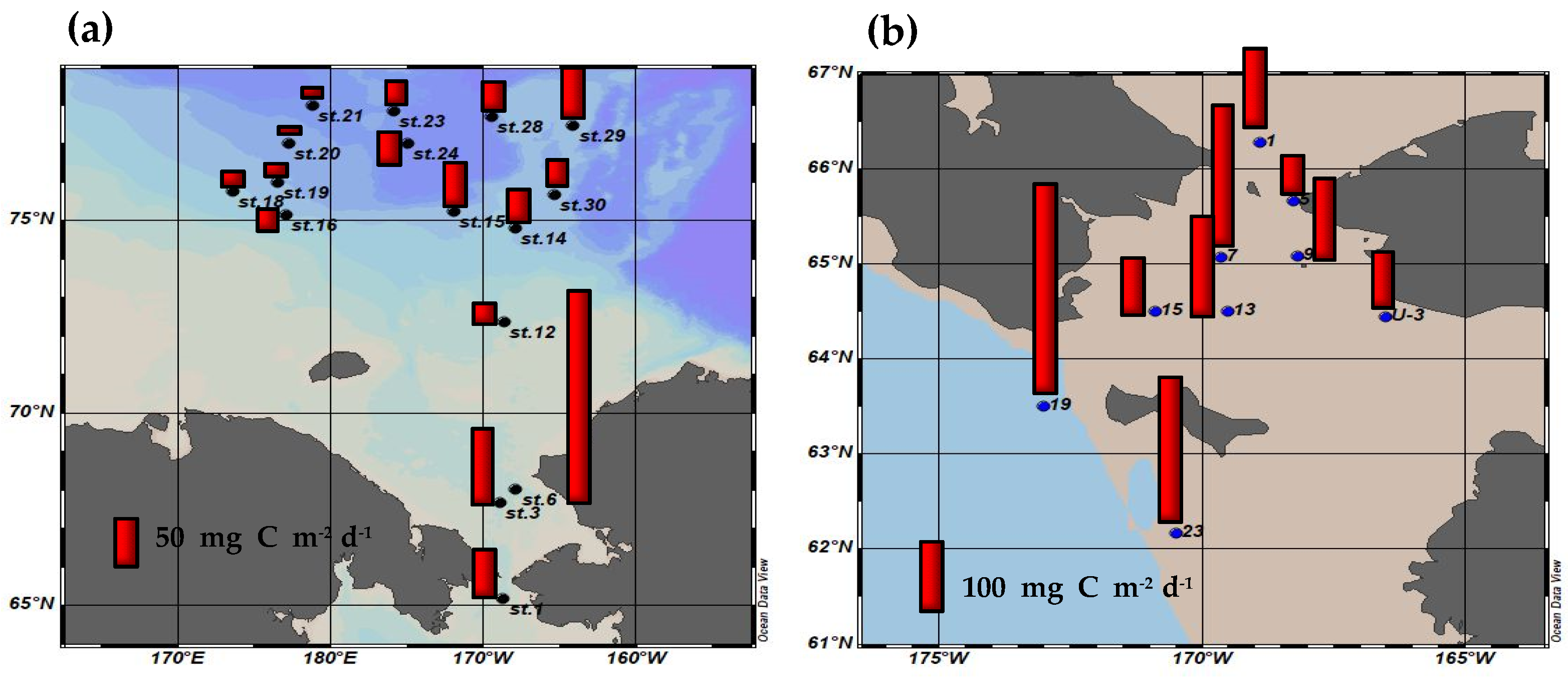
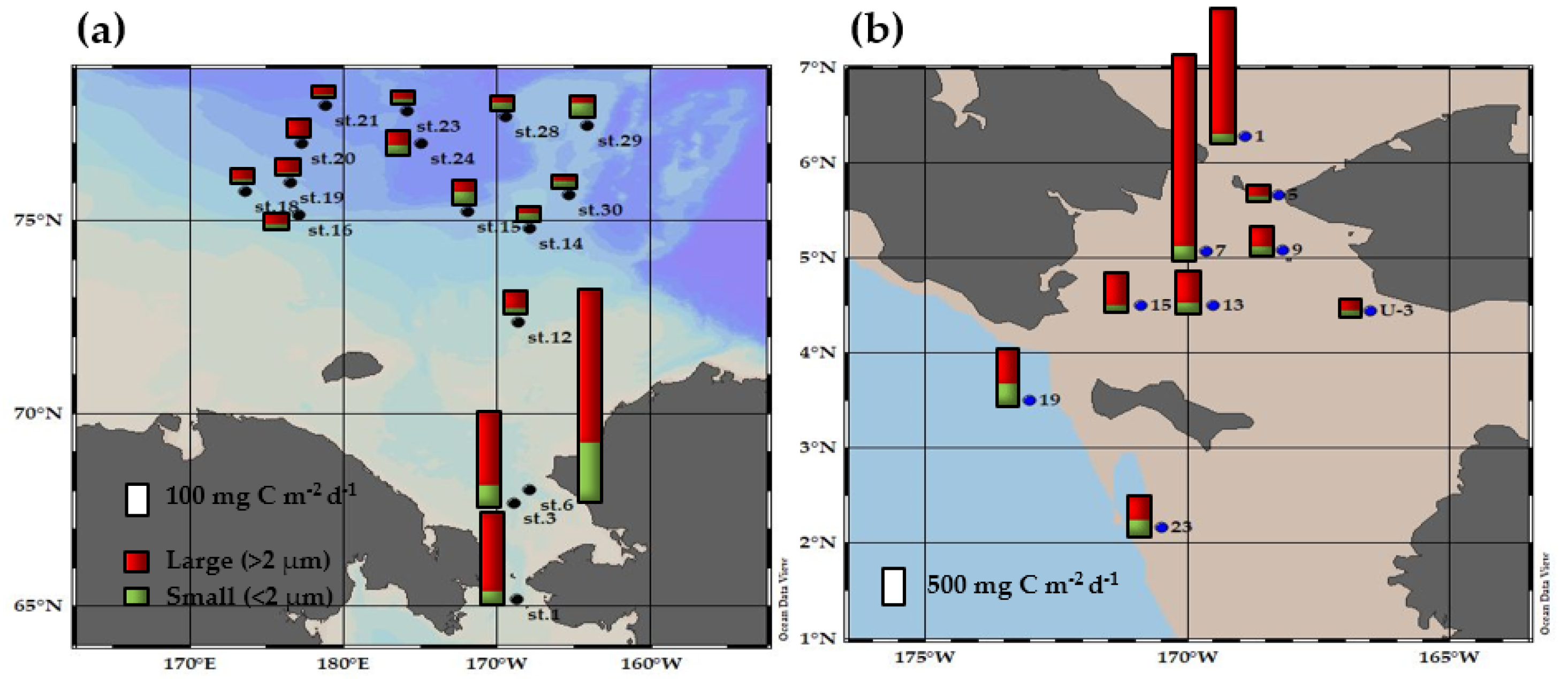
3.4. Primary Production Contribution of Small Phytoplankton and Their Ecological Roles

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Area | Year | Season | Smallcontribution | Methods | Size | References |
|---|---|---|---|---|---|---|
| Northern Chukchi Sea and Canada Basin | 2008 | August–September | 19.8–60.3% | In situ | <5 μm | [41] |
| Bering Strait and Chukchi Sea | 2004 | August–September | 31.72 ± 23.59% | In situ | <5 μm | [22] |
| Kara, Laptev and East Siberian Sea | 2013 | August–September | 52.7–71.2% | In situ | <5 μm | [43] |
| Barents Sea | 2003–2005 | Early to late bloom period | 31–87% | In situ | <10 μm | [44] |
| North water polynya | 1998 | April–July | 19% | In situ | <5 μm | [45] |
| Chukchi Sea and Bering Strait | 2016 | August | 38.0 ± 19.9% | In situ | <2 μm | This study |
| Northern Bering Sea and Bering Strait | 2017 | July | 25.0 ± 12.8% | In situ | <2 μm |
4. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sambrotto, R.N.; Goering, J.J.; McRoy, C.P. Large yearly production of phytoplankton in the western Bering Sea. Science 1984, 225, 1147–1150. [Google Scholar] [CrossRef] [PubMed]
- Springer, A.M.; McRoy, C.P. The paradox of pelagic food webs in the northern Bering Sea. III. Patterns of primary production. Cont. Shelf Res. 1993, 13, 575–599. [Google Scholar] [CrossRef]
- Lee, S.H.; Whitledge, T.E.; Kang, S.H. Recent carbon and nitrogen uptake rates of phytoplankton in Bering Strait and the Chukchi Sea. Cont. Shelf Res. 2007, 27, 2231–2249. [Google Scholar] [CrossRef]
- Bluhm, B.; Gradinger, R. Regional Variability in Food Availability for Arctic Marine Mammals. Ecol. Appl. 2008, 18, S77–S96. [Google Scholar] [CrossRef] [PubMed]
- Grebmeier, J.M.; Bluhm, B.A.; Cooper, L.W.; Denisenko, S.G.; Iken, K.; Kędra, M.; Serratos, C. Time-series benthic community composition and biomass and associated environmental characteristics in the Chukchi Sea during the RUSALCA 2004–2012 Program. Oceanography 2015, 28, 116–133. [Google Scholar] [CrossRef]
- Overland, J.; Stabeno, P. Is the Climate of the Bering Sea Warming and Affecting the Ecosystem? EOS Trans. AGU 2004, 85, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Grebmeier, J.; Cooper, L.; Feder, H.; Sirenko, B. Ecosystem Dynamics of the Pacific-Influenced Northern Bering and Chukchi Seas in the Amerasian Arctic. Prog. Oceanogr. 2006, 71, 331–361. [Google Scholar] [CrossRef]
- Danielson, S.L.; Ahkinga, O.; Ashjian, C.; Basyuk, E.; Cooper, L.W.; Eisner, L.; Farley, E.; Iken, K.B.; Grebmeier, J.M.; Juranek, L.; et al. Manifestation and consequences of warming and altered heat fluxes over the Bering and Chukchi Sea continental shelves. Deep-Sea Res. II Top. Stud. Oceanogr. 2020, 177, 104871. [Google Scholar] [CrossRef]
- Woodgate, R. Increases in the Pacific inflow to the Arctic from 1990 to 2015, and insights into seasonal trends and driving mechanisms from year-round Bering Strait mooring data. Prog. Oceanogr. 2018, 160, 124–154. [Google Scholar] [CrossRef] [Green Version]
- Frey, K.E.; Moore, G.W.K.; Cooper, L.W.; Grebmeier, J.M. Divergent patterns of recent sea ice cover across the Bering, Chukchi, and Beaufort seas of the Pacific Arctic Region. Prog. Oceanogr. 2015, 136, 32–49. [Google Scholar] [CrossRef]
- Grebmeier, J.M. Shifting Patterns of Life in the Pacific Arctic and Sub-Arctic Seas. Annu. Rev. Mar. Sci. 2012, 4, 63–78. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Ryu, J.; Lee, D.; Park, J.W.; Kwon, J.I.; Zhao, J.; Son, S. Spatial variations of small phytoplankton contributions in the Northern Bering Sea and the Southern Chukchi Sea. GISci. Remote Sens. 2019, 56, 794–810. [Google Scholar] [CrossRef]
- Coupel, P.; Matsuoka, A.; Ruiz-Pino, D.; Gosselin, M.; Marie, D.; Tremblay, J.-É.; Babin, M. Pigment signatures of phyto- plankton communities in the Beaufort Sea. Biogeosciences 2015, 12, 991–1006. [Google Scholar] [CrossRef] [Green Version]
- Yun, M.S.; Whitledge, T.E.; Stockwell, D.; Son, S.H.; Lee, J.H.; Park, J.W.; Lee, D.B.; Park, J.; Lee, S.H. Primary production in the Chukchi Sea with potential effects of freshwater content. Biogeosciences 2016, 13, 737–749. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, A.; Hirawake, T.; Suzuki, K.; Eisner, L.; Imai, I.; Nishino, S.; Kikuchi, T.; Saitoh, S.-I. Influence of timing of sea ice retreat on phytoplankton size during marginal ice zone bloom period on the Chukchi and Bering shelves. Biogeosciences 2016, 13, 115–131. [Google Scholar] [CrossRef] [Green Version]
- Kahru, M.; Brotas, V.; Manzano-Sarabia, M.; Mitchell, B.G. Are phytoplankton blooms occurring earlier in the Arctic? Glob. Chang. Biol. 2011, 17, 1733–1739. [Google Scholar] [CrossRef]
- Arrigo, K.R.; van Dijken, G.L. Continued increases in Arctic Ocean primary production. Prog. Oceanogr. 2015, 136, 60–70. [Google Scholar] [CrossRef]
- Li, W.; McLaughlin, F.; Lovejoy, C.; Carmack, E. Smallest Algae Thrive as the Arctic Ocean Freshens. Science 2009, 326, 539. [Google Scholar] [CrossRef] [Green Version]
- Neeley, A.; Harris, L.; Frey, K. Unraveling phytoplankton community dynamics in the Northern Chukchi Sea under sea-ice covered, and sea-ice-free conditions. Geophys. Res. Lett. 2018, 45, 7663–7671. [Google Scholar] [CrossRef]
- Lim, Y.J.; Kim, T.W.; Lee, S.; Lee, D.; Park, J.; Kim, B.K.; Lee, S.H. Seasonal Variations in the Small Phytoplankton Contribution to the Total Primary Production in the Amundsen Sea, Antarctica. J. Geophys. Res. Ocean. 2019, 124, 8324–8341. [Google Scholar] [CrossRef]
- Lee, S.H.; Yun, M.S.; Kim, B.K.; Joo, H.T.; Kang, S.H.; Kang, C.K.; Whitledge, T.E. Contribution of small phytoplankton to total primary production in the Chukchi Sea. Cont. Shelf Res. 2013, 68, 43–50. [Google Scholar] [CrossRef]
- Joo, H.T.; Park, J.W.; Son, S.H.; Noh, J.H.; Jeong, J.Y.; Kwak, J.H.; Saux-Picart, S.; Choi, J.H.; Kang, C.K.; Lee, S.H. Long-term annual primary production in the Ulleung Basin as a biological hot spot in the East/Japan Sea. J. Geophys. Res. Ocean. 2014, 119, 5. [Google Scholar] [CrossRef] [Green Version]
- Yun, M.S.; Joo, H.T.; Park, J.W.; Kang, J.J.; Kang, S.H.; Lee, S.H. Lipid-rich and protein-poor carbon allocation patterns of phytoplankton in the northern Chukchi Sea, 2011. Cont. Shelf Res. 2018, 158, 26–32. [Google Scholar] [CrossRef]
- Frigstad, H.; Andersen, T.; Bellerby, R.G.J.; Silyakova, A.; Hessen, D. Variation in the seston C:N ratio of the Arctic Ocean and pan-Arctic shelves. J. Mar. Syst. 2014, 129, 214–223. [Google Scholar] [CrossRef] [Green Version]
- Kirk, J.T.O. Effects of suspensoids (turbidity) on penetration of solar radiation in aquatic ecosystems, in Perspectives in Southern Hemisphere. Hydrobiologia 1985, 28, 195–208. [Google Scholar] [CrossRef]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. A manual of chemical & biological methods for seawater analysis. Mar. Pollut. Bull. 1984, 1, 101–112. [Google Scholar]
- Suzuki, K.; Hinuma, A.; Saito, H.; Kiyosawa, H.; Liu, H.; Saino, T.; Tsuda, A. Responses of phytoplankton and heterotrophic bacteria in the northwest subarctic Pacific to in situ iron fertilization as estimated by HPLC pigment analysis and flow cytometry. Prog. Oceanogr. 2005, 64, 167–187. [Google Scholar] [CrossRef]
- Wright, S.W.; Jeffrey, S.W.; Mantoura, R.F.C.; Llewellyn, C.A.; Bjørnland, T.; Repeta, D.; Welschmeyer, N. Improved HPLC method for the analysis of chlorophylls and carotenoids from marine phytoplankton. Mar. Ecol. Prog. Ser. 1991, 77, 183–196. [Google Scholar] [CrossRef]
- Vidussi, F.; Roy, S.; Lovejoy, C.; Gammelgaard, M.; Thomsen, H.A.; Booth, B.; Tremblay, J.-É.; Mostajir, B. Spatial and temporal variability of the phytoplankton community structure in the North Water Polynya, investigated using pigment biomarkers. Can. J. Fish. Aquat. Sci. 2004, 61, 2038–2052. [Google Scholar] [CrossRef]
- Fujiwara, A.; Hirawake, T.; Suzuki, K.; Saitoh, S.-I. Remote sensing of size structure of phytoplankton communities using optical properties of the Chukchi and Bering Sea shelf region. Biogeosciences 2011, 8, 3567–3580. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, A.; Hirawake, T.; Suzuki, K.; Imai, I.; Saitoh, S.-I. Timing of sea ice retreat can alter phytoplankton community structure in the western Arctic Ocean. Biogeosciences 2014, 11, 1705–1716. [Google Scholar] [CrossRef] [Green Version]
- Ardyna, M.; Mundy, C.J.; Mills, M.M.; Oziel, L.; Grondin, P.-L.; Lacour, L.; Verin, G.; Dijken, G.; Ras, J.; Alou-Font, E.; et al. Environmental drivers of under-ice phytoplankton bloom dynamics in the Arctic Ocean. Elem. Sci. Anthr. 2020, 8, 30. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, H.J.; Whitledge, T.E. High incorporation of carbon into proteins by the phytoplankton of the Bering Strait and Chukchi Sea. Cont. Shelf Res. 2009, 29, 1689–1696. [Google Scholar] [CrossRef]
- Hama, T.; Miyazaki, T.; Ogawa, Y.; Iwakuma, T.; Takahashi, M.; Otsuki, A.; Ichmura, S. Measurement of photosynthetic production of a marine phytoplankton population using a sTable 13C isotope. Mar. Biol. 1983, 73, 31–36. [Google Scholar] [CrossRef]
- Danielson, S.A.; Eisner, L.; Ladd, C.; Mordy, C.; Sousa, L.; Weingartner, T.J. A comparison between late summer 2012 and 2013 water masses, macronutrients, and phytoplankton standing crops in the northern Bering and Chukchi Seas. Deep-Sea Res. Part II 2017, 135, 7–26. [Google Scholar] [CrossRef] [Green Version]
- Coachman, L.K.; Aagaard, K.; Trip, R.B. Bering Strait: The Regional Physical Oceanography; University of Washington Press: Seattle, WA, USA, 1975. [Google Scholar]
- Springer, A.M.; McRoy, C.P.; Turco, K.R. The paradox of pelagic food webs in the northern Bering Sea—II. Zooplankton communities. Cont. Shelf Res. 1989, 9, 359–386. [Google Scholar] [CrossRef]
- Yun, M.S.; Kim, B.K.; Joo, H.T.; Yang, E.J.; Nishino, S.; Chung, K.H.; Kang, S.H.; Lee, S.H. Regional productivity of phytoplankton in the Western Arctic Ocean during summer in 2010. Deep Sea Res. II 2015, 120, 61–71. [Google Scholar] [CrossRef]
- Yun, M.S.; Whitledge, T.E.; Kong, M.; Lee, S.H. Low primary production in the Chukchi Sea shelf, 2009. Cont. Shelf Res. 2014, 76, 1–11. [Google Scholar] [CrossRef]
- Lee, S.H.; Joo, H.M.; Yun, M.S.; Whiteledge, T.E. Recent phytoplankton productivity of the northern Bering Sea during early summer in 2007. Polar Biol. 2012, 35, 83–98. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, B.K.; Lim, Y.J.; Joo, H.; Kang, J.J.; Lee, D.; Park, J.; Ha, S.-Y.; Lee, S.H. Small phytoplankton contribution to the standing stocks and the total primary production in the Amundsen Sea. Biogeosciences 2017, 14, 3705–3713. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Joo, H.T.; Lee, J.H.; Lee, J.H.; Kang, J.J.; Lee, H.W.; Lee, D.B.; Kang, C.K. Seasonal carbon uptake rates of phytoplankton in the northern East/Japan Sea. Deep Sea Res. Part II Top. Stud. Oceanogr. 2017, 143, 45–53. [Google Scholar] [CrossRef]
- Bhavya, P.S.; Lee, J.H.; Lee, H.W.; Kang, J.J.; Lee, J.H.; Lee, D.B.; Ahn, S.H.; Stockwell, D.; Whitledge, T.E.; Lee, S.H. First in situ estimations of small phytoplankton carbon and nitrogen uptake rates in the Kara, Laptev, and East Siberian seas. Biogeosciences 2018, 15, 5503–5517. [Google Scholar] [CrossRef] [Green Version]
- Hodal, H.; Kristiansen, S. The importance of small-celled phytoplankton in spring blooms at the marginal ice zone in the northern Barents Sea. Deep Sea Res. Part II Top. Stud. Oceanogr. 2008, 55, 2176–2185. [Google Scholar] [CrossRef] [Green Version]
- Mei, Z.-P.; Legendre, L.; Gratton, Y.; Tremblay, J.-É.; LeBlanc, B.; Klein, B.; Goseelin, M. Phytoplankton production in the North Water Polynya: Size-fractions and carbon fluxes, April to July 1998. Mar. Ecol. Prog. Ser. 2003, 256, 13–27. [Google Scholar] [CrossRef]
- Kim, J.-M.; Lee, K.; Suh, Y.-S.; Han, I.-S. Phytoplankton Do Not Produce Carbon-Rich Organic Matter in High CO2 Oceans. Geophys. Res. Lett. 2018, 45, 4189–4197. [Google Scholar] [CrossRef]











| Station | Date | Latitude (°N) | Longitude (°E) | Bottom depth (m) | |
|---|---|---|---|---|---|
| ARA07B | st. 1 | 5 Aug 2016 | 65.17 | −168.69 | 49 |
| st. 3 | 6 Aug 2016 | 67.67 | −168.96 | 50 | |
| st. 6 | 6 Aug 2016 | 68.01 | −167.87 | 52.3 | |
| st. 12 | 8 Aug 2016 | 72.36 | −168.67 | 55 | |
| st. 14 | 9 Aug 2016 | 74.80 | −167.90 | 223 | |
| st. 15 | 17 Aug 2016 | 75.24 | −171.98 | 512 | |
| st. 16 | 10 Aug 2016 | 75.15 | −176.00 | 331.57 | |
| st. 18 | 11 Aug 2016 | 75.77 | 177.07 | 486 | |
| st. 19 | 12 Aug 2016 | 76.00 | 173.60 | 282 | |
| st. 20 | 12 Aug 2016 | 77.00 | 176.57 | 1232 | |
| st. 21 | 13 Aug 2016 | 78.00 | 177.28 | 1693 | |
| st. 23 | 15 Aug 2016 | 77.87 | −175.91 | 1564 | |
| st. 24 | 16 Aug 2016 | 77.00 | −175.00 | 2008 | |
| st. 28 | 18 Aug 2016 | 77.70 | −169.50 | 1756 | |
| st. 29 | 18 Aug 2016 | 77.47 | −164.12 | 280 | |
| st. 30 | 19 Aug 2016 | 76.58 | −165.38 | 987 | |
| OS040 | 1 | 9 Jul 2017 | 66.28 | −168.90 | 57 |
| 5 | 11 Jul 2017 | 65.66 | −168.26 | 45 | |
| 7 | 12 Jul 2017 | 65.06 | −169.64 | 51 | |
| 9 | 14 Jul 2017 | 65.07 | −168.19 | 42 | |
| U−3 | 16 Jul 2017 | 64.44 | −166.52 | 27 | |
| 13 | 17 Jul 2017 | 64.50 | −169.52 | 42 | |
| 15 | 18 Jul 2017 | 64.50 | −170.89 | 46 | |
| 19 | 19 Jul 2017 | 63.50 | −173.02 | 66 | |
| 23 | 21 Jul 2017 | 62.17 | −170.50 | 46 |
| Class | chl-b | chl-c3 | fucox | perid | allox | 19butfu | 19hexfu | chl-c | neox | prasinox | lut |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Initial ratio matrix | |||||||||||
| Diatoms | 0 | 0 | 0.425 | 0 | 0 | 0 | 0 | 0.171 | 0 | 0 | 0 |
| Dinoflagellates | 0 | 0 | 0 | 0.6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Cryptophytes | 0 | 0 | 0 | 0 | 0.673 | 0 | 0 | 0 | 0 | 0 | 0 |
| Chryso-Pelago | 0 | 0.114 | 0.285 | 0 | 0 | 0.831 | 0 | 0.285 | 0 | 0 | 0 |
| Prasino-2 | 0.812 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.033 | 0 | 0.096 |
| Prasino-3 | 0.764 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.078 | 0.248 | 0.009 |
| Chlorophytes | 0.339 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.036 | 0 | 0.187 |
| Phaeocystis | 0 | 0.208 | 0.35 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Hapto-7 | 0 | 0.171 | 0.259 | 0 | 0 | 0.013 | 0.491 | 0.276 | 0 | 0 | 0 |
| ARA07B Final ratio matrix | |||||||||||
| Diatoms | 0 | 0 | 0.785 | 0 | 0 | 0 | 0 | 0.395 | 0 | 0 | 0 |
| Dinoflagellates | 0 | 0 | 0 | 0.6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Cryptophytes | 0 | 0 | 0 | 0 | 0.673 | 0 | 0 | 0 | 0 | 0 | 0 |
| Chryso-Pelago | 0 | 0.114 | 0.285 | 0 | 0 | 0.831 | 0 | 0.285 | 0 | 0 | 0 |
| Prasino-2 | 0.593 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.007 | 0 | 0.007 |
| Prasino-3 | 4.006 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.166 | 0.803 | 0.027 |
| Chlorophytes | 0.339 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.036 | 0 | 0.187 |
| Phaeocystis | 0 | 0.1791 | 0.301 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Hapto-7 | 0 | 0.171 | 0.259 | 0 | 0 | 0.013 | 0.508 | 0.276 | 0 | 0 | 0 |
| OS040Final ratio matrix | |||||||||||
| Diatoms | 0 | 0 | 0.722 | 0 | 0 | 0 | 0 | 0.328 | 0 | 0 | 0 |
| Dinoflagellates | 0 | 0 | 0 | 1.409 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Cryptophytes | 0 | 0 | 0 | 0 | 0.673 | 0 | 0 | 0 | 0 | 0 | 0 |
| Chryso-Pelago | 0 | 0.114 | 0.285 | 0 | 0 | 0.831 | 0 | 0.285 | 0 | 0 | 0 |
| Prasino-2 | 0.812 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.033 | 0 | 0.096 |
| Prasino-3 | 0.280 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.107 | 0.471 | 0.011 |
| Chlorophytes | 0.643 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.029 | 0 | 0.969 |
| Phaeocystis | 0 | 0.558 | 1.457 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Hapto-7 | 0 | 0.171 | 0.259 | 0 | 0 | 0.013 | 0.617 | 0.276 | 0 | 0 | 0 |
| Cluster | T (°C) | S (psu) | Small Contribution to PP | Small Contribution to Surface chl-a | Small Contribution to POC | PP (mg C m−2 d−1) | Chl-a (mg m−3) | POC (mg m−3) |
|---|---|---|---|---|---|---|---|---|
| 1 | 3.5 | 32.7 | 6.0% | 2.9% | 25.0% | 1992.9 | 66.3 | 349.0 |
| 2 | 5.5 | 32.7 | 6.9% | 0.7% | 18.6% | 3100.1 | 128.8 | 479.5 |
| 3 | 0.1 | 29.7 | 40.5% | 39.1% | 40.9% | 79.6 | 13.9 | 177.7 |
| 4 | 5.6 | 31.7 | 26.7% | 39.1% | 43.9% | 559.2 | 60.7 | 236.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.-W.; Kim, Y.; Kim, K.-W.; Fujiwara, A.; Waga, H.; Kang, J.J.; Lee, S.-H.; Yang, E.-J.; Hirawake, T. Contribution of Small Phytoplankton to Primary Production in the Northern Bering and Chukchi Seas. Water 2022, 14, 235. https://doi.org/10.3390/w14020235
Park J-W, Kim Y, Kim K-W, Fujiwara A, Waga H, Kang JJ, Lee S-H, Yang E-J, Hirawake T. Contribution of Small Phytoplankton to Primary Production in the Northern Bering and Chukchi Seas. Water. 2022; 14(2):235. https://doi.org/10.3390/w14020235
Chicago/Turabian StylePark, Jung-Woo, Yejin Kim, Kwan-Woo Kim, Amane Fujiwara, Hisatomo Waga, Jae Joong Kang, Sang-Heon Lee, Eun-Jin Yang, and Toru Hirawake. 2022. "Contribution of Small Phytoplankton to Primary Production in the Northern Bering and Chukchi Seas" Water 14, no. 2: 235. https://doi.org/10.3390/w14020235
APA StylePark, J. -W., Kim, Y., Kim, K. -W., Fujiwara, A., Waga, H., Kang, J. J., Lee, S. -H., Yang, E. -J., & Hirawake, T. (2022). Contribution of Small Phytoplankton to Primary Production in the Northern Bering and Chukchi Seas. Water, 14(2), 235. https://doi.org/10.3390/w14020235