
Age Structure and Spatial Distribution of Euphausia superba Larvae off the Antarctic Peninsula, Southern Ocean

Abstract
:1. Introduction
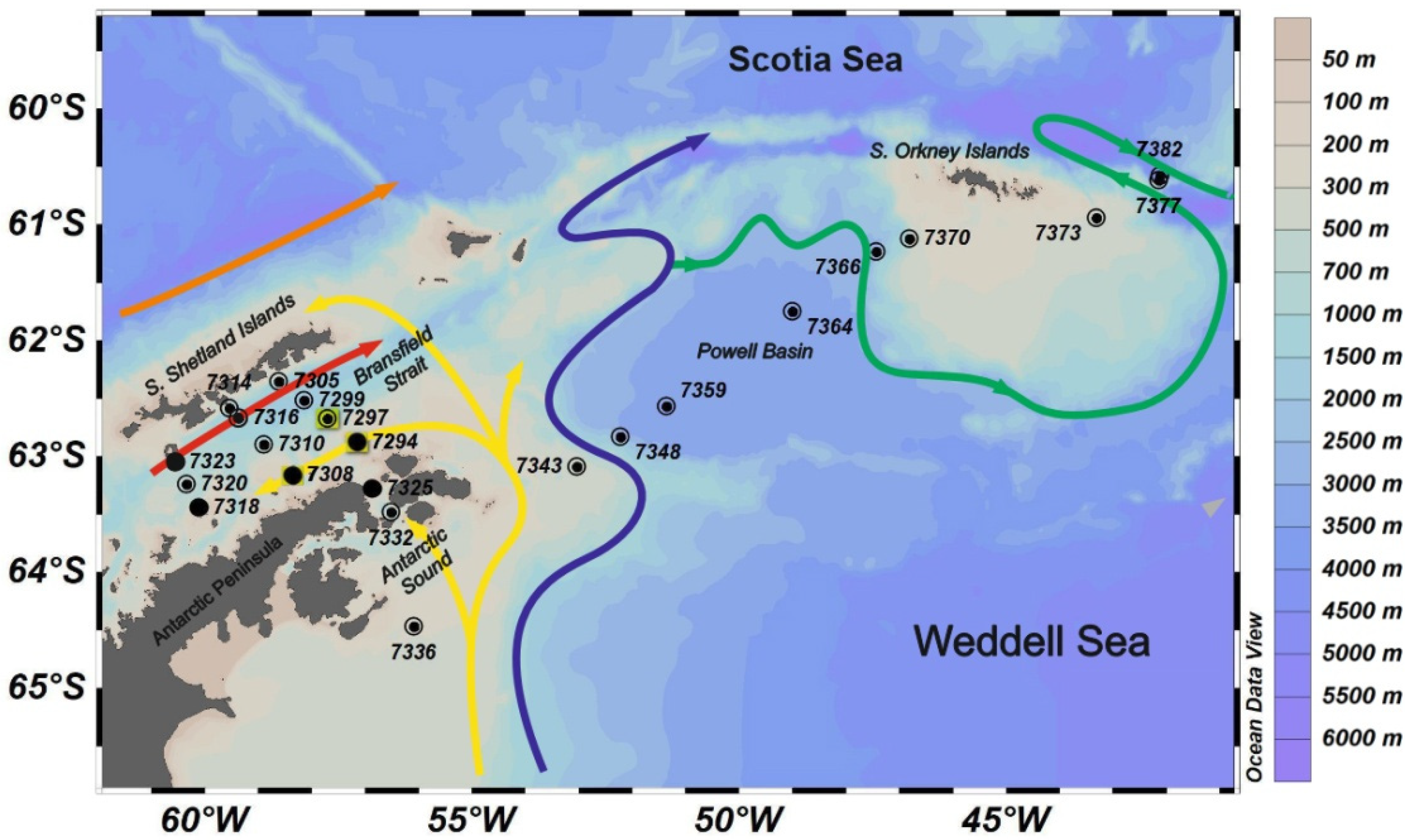
2. Materials and Methods
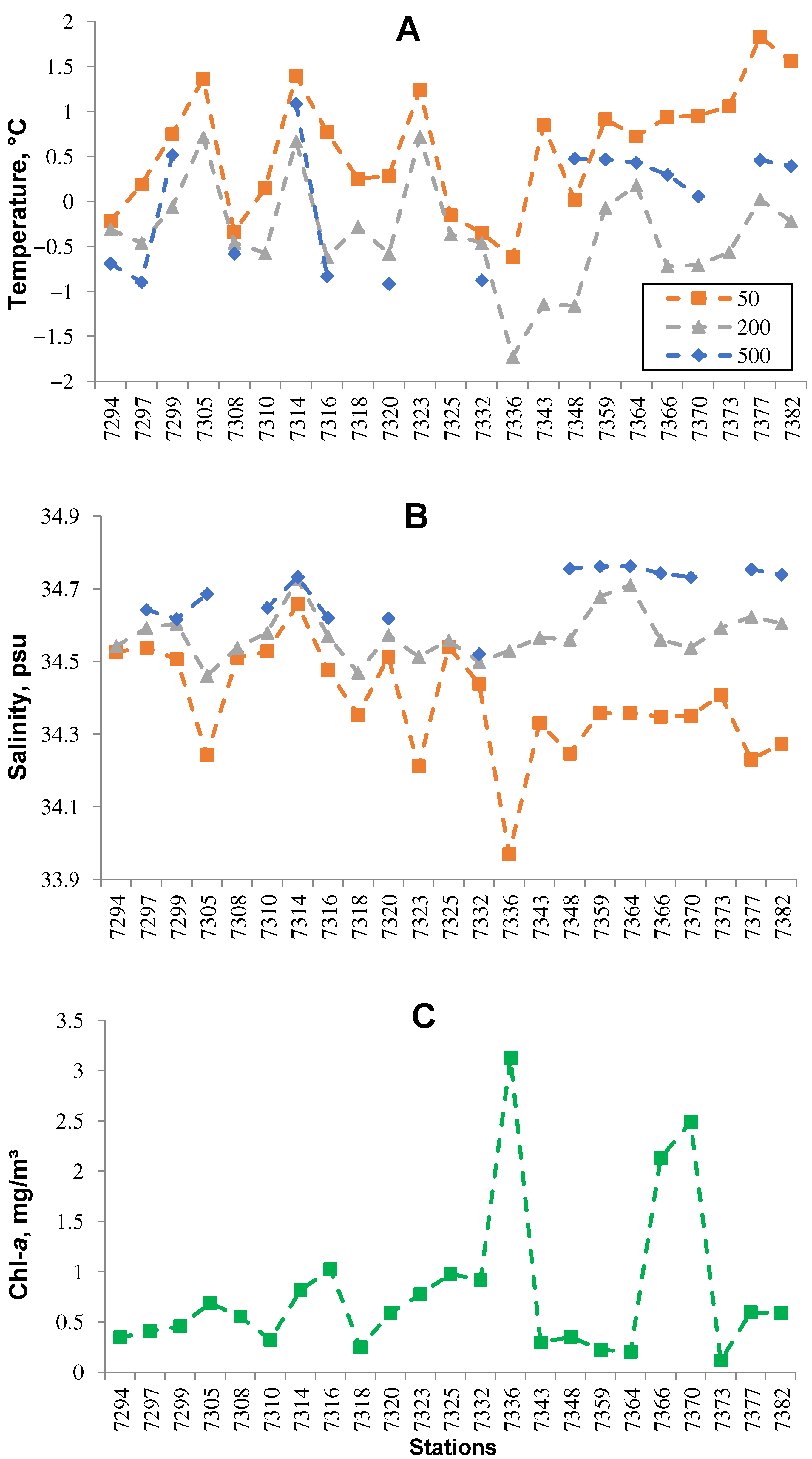
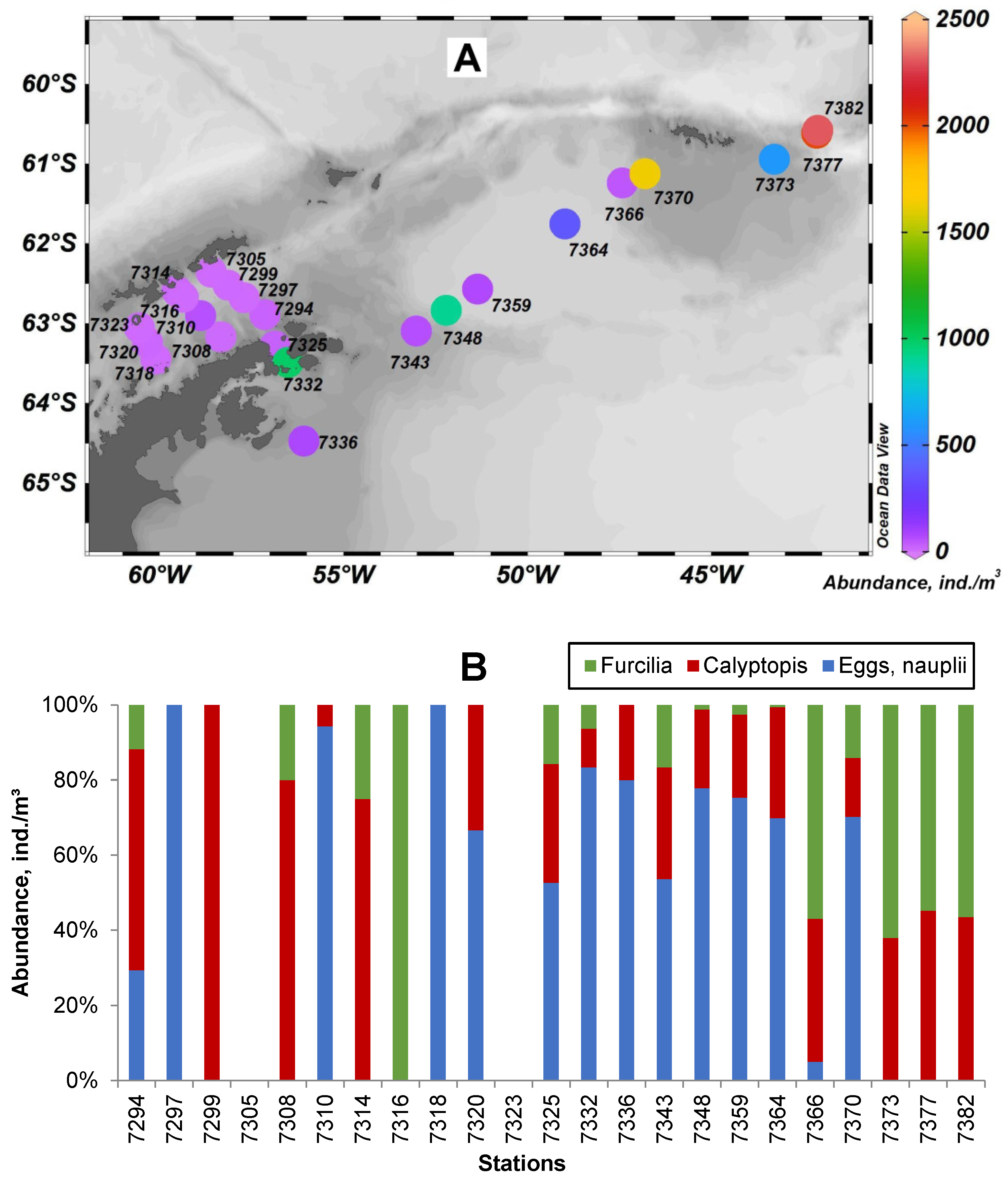
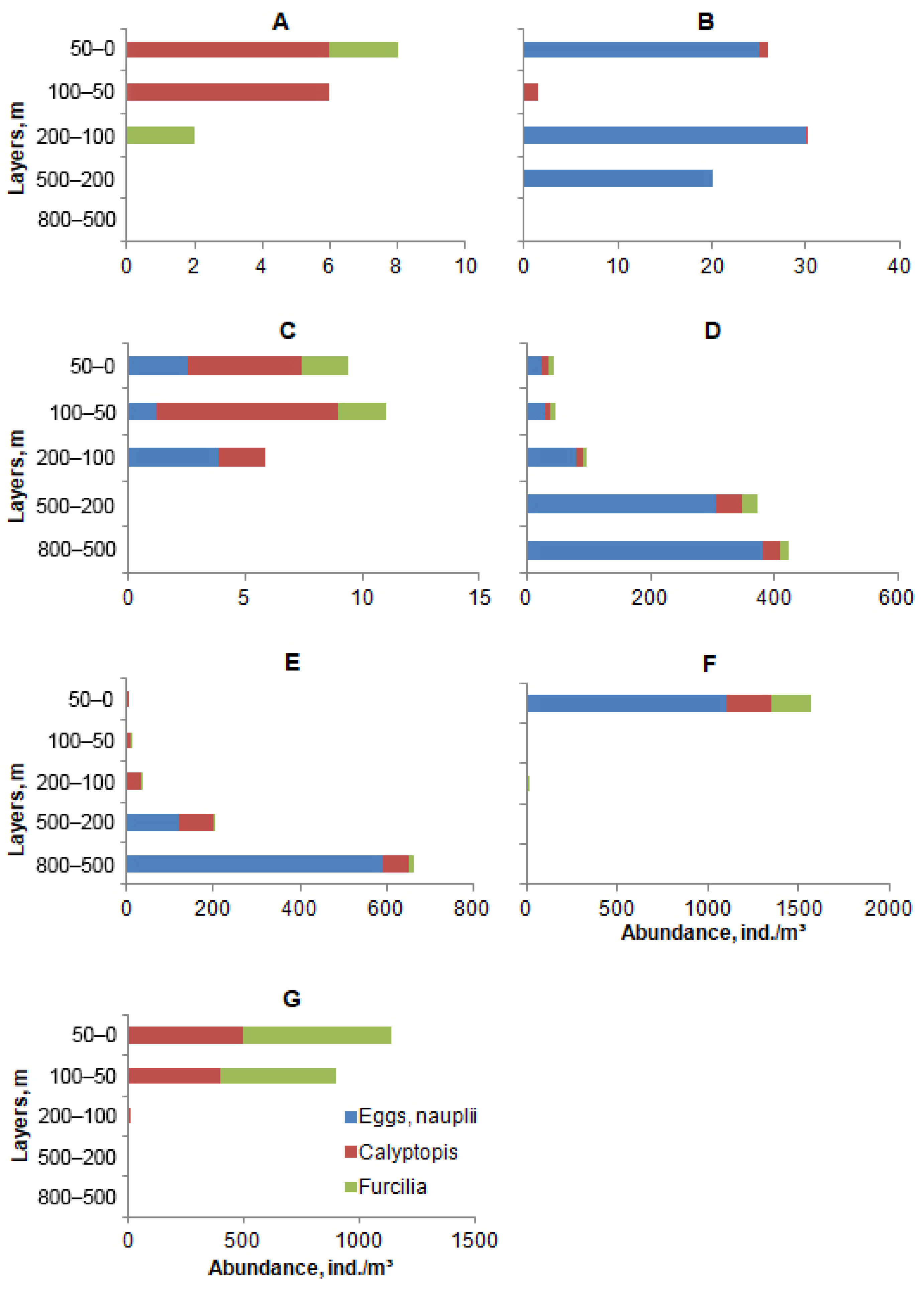
3. Results
4. Discussion and Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laws, R. Seals and whales of the Southern Ocean. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1977, 279, 81–96. [Google Scholar] [CrossRef]
- Kawaguchi, S.; Nicol, S. Learning about Antarctic krill from the fishery. Antar. Sci. 2007, 19, 219–230. [Google Scholar] [CrossRef]
- Santa Cruz, F.; Ernst, B.; Arata, J.A.; Parada, C. Spatial and temporal dynamics of the Antarctic krill fishery in fishing hotspots in the Bransfield Strait and South Shetland Islands. Fish. Res. 2018, 208, 157–166. [Google Scholar] [CrossRef]
- Atkinson, A.; Siegel, V.; Pakhomov, E.; Rothery, P. Long-term decline in krill stock and increase in salps within the Southern Ocean. Nature 2004, 432, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, A.; Hill, S.L.; Pakhomov, E.A.; Siegel, V.; Reiss, C.S.; Loeb, V.J.; Steinberg, D.K.; Schmidt, K.; Tarling, G.A.; Gerrish, L.; et al. Krill (Euphausia superba) distribution contracts southward during rapid regional warming. Nat. Clim. Change 2019, 9, 142–147. [Google Scholar] [CrossRef]
- Yang, G.; Atkinson, A.; Hill, S.L.; Guglielmo, L.; Granata, A.; Li, C. Changing circumpolar distributions and isoscapes of Antarctic krill: Indo-Pacific habitat refuges counter long-term degradation of the Atlantic sector. Limnol. Oceanogr. 2020, 66, 272–287. [Google Scholar] [CrossRef]
- Voronina, N.M. Comparative abundance and distribution of major filter-feeders in the Antarctic pelagic zone. J. Mar. Sys. 1998, 17, 375–390. [Google Scholar] [CrossRef]
- Hewes, C.D.; Reiss, C.S.; Holm-Hansen, O. A quantitative analysis of sources for summertime phytoplankton variability over 18 years in the South Shetland Islands (Antarctica) region. Deep-Sea Res. II 2009, 56, 1230–1241. [Google Scholar] [CrossRef]
- Garcia, M.D.; Dutto, M.S.; Chazarreta, C.J.; Berasategui, A.A.; Schloss, I.R.; Hoffmeyer, M.S. Micro- and mesozooplankton successions in an Antarctic coastal environment during a warm year. PLoS ONE 2020, 15, e0232614. [Google Scholar] [CrossRef] [PubMed]
- Pakhomov, E.; Pshenichnov, L.; Krot, A.; Paramonov, V.; Slypko, I.; Zabroda, P. Zooplankton distribution and community structure in the Pacific and Atlantic Sectors of the Southern Ocean during austral summer 2017–2018: A Pilot Study Conducted from Ukrainian Long-Liners. J. Mar. Sci. Eng. 2020, 8, 488. [Google Scholar] [CrossRef]
- Makarov, R.R. Larvae of Euphausia superba Dana in plankton from the Sea of Scotia. Tr. Vses. Nauchno-Issled. Ryb. Okeanogr. 1974, 99, 84–103. [Google Scholar]
- Shulenberger, E.; Wormuth, J.H.; Loeb, V.J. A large swarm of Euphausia superba: Overview of path structure and composition. J. Crustacean Biol. 1984, 4, 75–95. [Google Scholar] [CrossRef]
- Gao, Q.; Xu, Z.; Huang, H.; Chen, X.; Feng, C.; Li, L. Geographical distribution and age composition of Euphausia superba larvae (Crustacea: Euphausiacea) in the South Shetland Islands region and southern Scotia Sea in relation to environmental conditions. Acta Oceanol. Sin. 2013, 32, 59–67. [Google Scholar] [CrossRef]
- Sologub, D.O. Distribution and age structure of euphausiids larvae at the Western Antarctic Peninsula (Subarea 48.1). Trudy VNIRO 2015, 154, 3–15. [Google Scholar]
- Yakovenko, V.A.; Spiridonov, V.A.; Gorbatenko, K.M.; Shadrin, N.V.; Samyshev, E.Z.; Minkina, N.I. Macro- and mesozooplankton in the Powell Basin (Antarctica): Species composition and distribution of abundance and biomass in February 2020. In Antarctic Peninsula Region of the Southern Ocean; Morozov, E.G., Flint, M.V., Spiridonov, V.A., Eds.; Springer: Cham, Switzerland, 2021; Volume 6, pp. 131–141. [Google Scholar] [CrossRef]
- Meredith, M.P.; King, J.C. Rapid climate change in the ocean west of the Antarctic Peninsula during the second half of the 20th century. Geophys. Res. Let. 2005, 32, L19604. [Google Scholar] [CrossRef]
- Stammerjohn, S.E.; Martinson, D.G.; Smith, R.C.; Yuan, X.; Rind, D. Trends in Antarctic annual sea ice retreat and advance and their relation to El Nino-Southern Oscillation and Southern Annular Mode variability. J. Geophys. Res Ocean. 2008, 113, C03S90. [Google Scholar] [CrossRef] [Green Version]
- Meredith, M.; Sommerkorn, S.; Cassotta, C.; Derksen, A.; Ekaykin, A.; Hollowed, G.; Kofinas, A.; Mackintosh, J.; Melbourne-Thomas, M.M.C.; Muelbert, G.; et al. Polar Regions. In IPCC Special Report on the Ocean and Cryosphere in a Changing Climate; Pörtner, H.-O., Roberts, D.C., Masson-Delmotte, V., Zhai, P., Tignor, M., Poloczanska, E., Mintenbeck, K., Alegría, A., Nicolai, M., Okem, A., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2019; pp. 203–320. [Google Scholar] [CrossRef]
- Anisimov, O.A.; Vaughan, D.G.; Callaghan, T.V.; Furgal, C.; Marchant, H.; Prowse, T.D.; Vilhjalmsson, H.; Walsh, J.E. Polar regions (Arctic and Antarctic). In Climate Change 2007: Impacts, Adaptation and Vulnerability; Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Parry, M.L., Canziani, O.F., Palutikof, P.J., van der Linden, P., Hanson, C.E., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 653–685. [Google Scholar]
- Xiao, C. Changes in antarctic climate system: Past, present and future. Adv. Clim. Change Res. 2008, 4, 1–7. [Google Scholar]
- Behrenfeld, M.J. Climate-mediated dance of the plankton. Nat. Clim. Change 2014, 4, 880–887. [Google Scholar] [CrossRef]
- Johnston, N.M.; Murphy, E.J.; Atkinson, A.; Constable, A.J.; Cotte, C.; Cox, M.; Daly, K.L.; Driscoll, R.; Flores, H.; Halfter, S.; et al. Status, change, and futures of zooplankton in the Southern Ocean. Front. Ecol. Evol. 2022, 9, 624692. [Google Scholar] [CrossRef]
- Ross, R.M.; Quetin, L.B.; Kirsch, E. Effect of temperature on developmental times and survival of early larval stages of Euphausia superba Dana. J. Exp. Mar. Bio. Ecol. 1988, 121, 55–71. [Google Scholar] [CrossRef]
- Perry, F.A.; Kawaguchi, S.; Atkinson, A.; Sailley, S.F.; Tarling, G.A.; Mayor, D.J.; Lucas, C.H.; King, R.; Cooper, A. Temperature–induced hatch failure and nauplii malformation in Antarctic krill. Front. Mar. Sci. 2020, 7, 501. [Google Scholar] [CrossRef]
- Weikert, H.; John, H.-C. Experiences with a modified be multiple opening–closing plankton net. J. Plankton Res. 1981, 3, 167–176. [Google Scholar] [CrossRef]
- Tranter, D.J. Reviews on zooplankton sampling methods. In Zooplankton Sampling; Part I; Tranter, D.J., Ed.; UNESCO: Paris, France, 1968; pp. 11–144. [Google Scholar]
- Smith, P.E.; Richardson, S. Standard techniques for pelagic fish egg and larva surveys. FAO Fish. Tech. Pap. 1977, 175, 27–73. [Google Scholar]
- Pertsova, K.N. Larvae of euphausiids of the Antarctic. In Ecology and biogeography of plankton: Proceedings of the Institute of Oceanology RAS; Nauka: Moscow, Russia, 1976; Volume 105, pp. 147–170. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER–E Ltd.: Plymouth, UK, 2006; p. 192. [Google Scholar]
- Clarke, K.R.; Ainsworth, M. A method of linking multivariate community structure to environmental variables. Mar. Ecol. Prog. Ser. 1993, 92, 205–219. [Google Scholar] [CrossRef]
- Heywood, K.J.; Garabato, A.C.N.; Stevens, D.P.; Muench, R.D. On the fate of the Antarctic Slope Front and the origin of the Weddell Front. J. Geophys. Res. 2004, 109, C06021. [Google Scholar] [CrossRef] [Green Version]
- Thompson, A.F.; Heywood, K.J.; Thorpe, S.E.; Renner, A.H.H.; Trasvica, A. Surface circulation at the tip of the Antarctic Peninsula from drifters. J. Phys. Oceanog. 2009, 39, 3–26. [Google Scholar] [CrossRef]
- Morozov, E.G.; Flint, M.V.; Spiridonov, V.A. Antarctic Peninsula region of the Southern Ocean; Advances in Polar Ecology; Springer: Cham, Switzerland, 2021; Volume 6, p. 433. [Google Scholar] [CrossRef]
- Zhou, X.Q.; Zhu, G.P.; Hu, S. Influence of tides on mass transport in the Bransfield Strait and the adjacent areas. Antarctic Polar Sci. 2020, 23, 100506. [Google Scholar] [CrossRef]
- Fedotova, A.A.; Kashin, S.V. Interannual variations of water mass properties in the central basin of the Bransfield Strait. In Antarctic Peninsula Region of the Southern Ocean; Morozov, E.G., Flint, M.V., Spiridonov, V.A., Eds.; Springer: Cham, Switzerland, 2021; Volume 6, pp. 131–141. [Google Scholar] [CrossRef]
- Krechik, V.A.; Frey, D.I.; Morozov, E.G. Peculiarities of water circulation in the central part of the Bransfield Strait in January 2020. Dokl. Earth Sci. 2021, 496, 92–95. [Google Scholar] [CrossRef]
- Tokarczyk, R. Classification of water masses in the Bransfield Strait and southern part of the Drake Passage using a method of statistical multidimensional analysis. Polish Pol. Res. 1987, 8, 333–366. [Google Scholar]
- Krek, A.V.; Krek, E.V.; Krechik, V.A. The Circulation and Mixing Zone in the Antarctic Sound in February 2020. In Antarctic Peninsula Region of the Southern Ocean; Morozov, E.G., Flint, M.V., Spiridonov, V.A., Eds.; Springer: Cham, Switzerland, 2021; Volume 6, pp. 83–100. [Google Scholar] [CrossRef]
- Fedotova, A.A.; Stepanova, S.V. Water mass transformation in the Powell Basin. In Antarctic Peninsula Region of the Southern Ocean; Morozov, E.G., Flint, M.V., Spiridonov, V.A., Eds.; Springer: Cham, Switzerland, 2021; Volume 6, pp. 115–129. [Google Scholar] [CrossRef]
- Deacon, G.E.R. A general account of the hydrology of the South Atlantic Ocean. Discov. Rep. 1933, 7, 171–238. [Google Scholar]
- Kittel, W.; Jazdzewski, K. Studies on the larval stages of Euphausia superba Dana (Crustacea, Euphausiacea) in the southern Drake Passage and in the Bransfield Strait in February and March 1981 during the BIOMASS-FIBEX expedition. Pol. Polar Res. 1982, 3, 273–280. [Google Scholar]
- Spiridonov, V.A.; Zalota, A.K.; Yakovenko, V.A.; Gorbatenko, K.M. Composition of population and transport of juveniles of Antarctic krill in Powell Basin region (northwestern Weddell Sea) in January 2020. Trudy VNIRO 2020, 181, 33–51. [Google Scholar] [CrossRef]
- Voronina, N.M. Pelagic Ecosystems of the Southern Ocean; Nauka: Moscow, Russia, 1984; p. 206. [Google Scholar]
- Orsi, A.H.; Whitworth, T.; Nowlin, W.D., Jr. On the meridional extent and fronts of the Antarctic Circumpolar Current. Deep Sea Res. I 1995, 42, 641–673. [Google Scholar] [CrossRef]
- Morozov, E.G.; Krechik, V.A.; Frey, D.I.; Polukhin, A.A.; Artemiev, V.A.; Kasyan, V.V.; Sapozhnikov, P.V.; Mukhametianov, R.Z. Frontal Zone Between Relatively Warm and Cold Waters in the Northern Weddell Sea. In Antarctic Peninsula Region of the Southern Ocean; Morozov, E.G., Flint, M.V., Spiridonov, V.A., Eds.; Springer: Cham, Switzerland, 2021; Volume 6, pp. 31–54. [Google Scholar] [CrossRef]
- Melnikov, I.; Spiridonov, V. Antarctic krill under perennial sea ice in the western Weddell Sea. Antarct. Sci. 1996, 8, 323–429. [Google Scholar] [CrossRef]
- Ross, R.M.; Quetin, L.B.; Baker, K.S.; Vernet, M.; Smith, R.C. Growth limitation in young Euphausia superba under field conditions. Limnol. Oceanogr. 2000, 45, 31–43. [Google Scholar] [CrossRef] [Green Version]
- Hempel, I.; Hempel, G. Field observations on the developmental ascent of larval Euphausia superba (Crustacea). Polar Biol. 1986, 6, 123–126. [Google Scholar] [CrossRef]
- Spiridonov, V.A. Spatial and temporal variability in reproductive timing of Antarctic krill (Euphausia superba). Polar Biol. 1995, 15, 161–174. [Google Scholar] [CrossRef]
- Makarov, R.R.; Menshenina, L.L. On the study of distribution and phenology of euphausiid larvae in the Scotia Sea. Antarctica 1988, 28, 166–174. [Google Scholar]
- Hofmann, E.E.; Klinck, J.M.; Locarnini, R.A.; Fach, B. Krill transport in the Scotia Sea and environs. Antarct. Sci. 1998, 10, 406–415. [Google Scholar] [CrossRef] [Green Version]
- Siegel, V.; Watkins, J.L. Distribution, Biomass and Demography of Antarctic Krill, Euphausia superba. In Biology and Ecology of Antarctic Krill; Advances in Polar Ecology; Siegel, V., Ed.; Springer: Cham, Switzerland, 2016; pp. 21–100. [Google Scholar] [CrossRef]
- Meyer, B.; Atkinson, A.; Blume, B.; Bathmann, U.V. Feeding and energy budgets of larval Antarctic krill Euphausia superba in summer. Mar. Ecol. Prog. Ser. 2003, 57, 167–177. [Google Scholar] [CrossRef]
- Schmidt, K.; Atkinson, A.; Venables, H.J.; Pond, D.W. Early spawning of Antarctic krill in the Scotia Sea is fuelled by “superfluous” feeding on non–ice associated phytoplankton blooms. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2012, 59–60, 159–172. [Google Scholar] [CrossRef]
- Atkinson, A.; Ward, P.; Hunt, B.P.V.; Pakhomov, E.A.; Hosie, G.W. An overview of Southern Ocean zooplankton data: Abundance, biomass, feeding and functional relationships. CCAMLR Sci. 2012, 19, 171–218. [Google Scholar]
- McBride, M.M.; Dalpadado, P.; Drinkwater, K.F.; Godo, O.R.; Hobday, A.J.; Hollowed, A.B.; Kristiansen, T.; Murphy, E.J.; Ressler, P.H.; Subbey, S.; et al. Krill, climate, and contrasting future scenarios for Arctic and Antarctic fisheries. ICES J. Mar. Sci. 2014, 71, 1934–1955. [Google Scholar] [CrossRef] [Green Version]
- Hays, G.C.; Richardson, A.J.; Robinson, C. Climate change and marine plankton. Trends Ecol. Evol. 2005, 20, 337–344. [Google Scholar] [CrossRef]
- Constable, A.J.; Melbourne-Thomas, J.; Corney, S.P.; Arrigo, K.R.; Barbraud, C.; Barnes, D.K.A.; Bindoff, N.L.; Boyd, P.W.; Brandt, A.; Costa, D.P.; et al. Climate change and Southern Ocean ecosystems I: How changes in physical habitats directly affect marine biota. Glob. Change Biol. 2014, 20, 3004–3025. [Google Scholar] [CrossRef]
 —Multinet,
—Multinet,  —Bongo, and ●—WP-2.
—Multinet, —Bongo, and ●—WP-2.
—Bongo, and ●—WP-2.
—Multinet, —Bongo, and ●—WP-2.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Factors | ρ | Mean ± SD | |||
|---|---|---|---|---|---|
| Group A | Group B | Group C | Group D | ||
| SST, °C | 0.307 | 0.36 ± 0.65 | 1.48 ± 0.39 | 0.34 ± 0.61 | 0.22 ± 0.04 |
| 200 m T, °C | 0.096 | –0.48 ± 0.61 | –0.25 ± 0.30 | –0.54 ± 0.56 | –0.37 ± 0.13 |
| 500 m T, °C | 0.089 | 0.03 ± 0.75 | 0.43 ± 0.05 | 0.02 ± 0.63 | –0.90 ± 0.07 |
| SSS, psu | 0.101 | 34.43 ± 0.18 | 34.30 ± 0.09 | 34.35 ± 0.08 | 34.44 ± 0.13 |
| 200 m S, psu | 0.093 | 34.59 ± 0.06 | 34.61 ± 0.02 | 34.58 ± 0.09 | 34.53 ± 0.09 |
| 500 m S, psu | 0.081 | 34.69 ± 0.07 | 34.75 ± 0.01 | 34.69 ± 0.12 | 34.64 ± 0.03 |
| Chl a, mg/m3 | 0.163 | 0.89 ± 0.91 | 0.43 ± 0.27 | 0.99 ± 1.05 | 0.33 ± 0.11 |
| BEST combination of factors (SST and Chl a) | 0.369 | ||||
| SST | SSS | 200 m T | 200 m S | 500 m T | 500 m S | Chl a | E and N | C | F | |
|---|---|---|---|---|---|---|---|---|---|---|
| E and N | –0.38 | –0.16 | –0.44 | –0.02 | –0.08 | 0.29 | 0.36 | 1 | ||
| C | 0.22 | –0.32 | –0.20 | 0.37 | 0.41 | 0.57 | –0.09 | 0.58 | 1 | |
| F | 0.33 | –0.18 | –0.16 | 0.26 | 0.23 | 0.44 | –0.06 | 0.32 | 0.86 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasyan, V.V. Age Structure and Spatial Distribution of Euphausia superba Larvae off the Antarctic Peninsula, Southern Ocean. Water 2022, 14, 3196. https://doi.org/10.3390/w14203196
Kasyan VV. Age Structure and Spatial Distribution of Euphausia superba Larvae off the Antarctic Peninsula, Southern Ocean. Water. 2022; 14(20):3196. https://doi.org/10.3390/w14203196
Chicago/Turabian StyleKasyan, Valentina V. 2022. "Age Structure and Spatial Distribution of Euphausia superba Larvae off the Antarctic Peninsula, Southern Ocean" Water 14, no. 20: 3196. https://doi.org/10.3390/w14203196
APA StyleKasyan, V. V. (2022). Age Structure and Spatial Distribution of Euphausia superba Larvae off the Antarctic Peninsula, Southern Ocean. Water, 14(20), 3196. https://doi.org/10.3390/w14203196