
Fresh Air for the Mire-Breathing Hypothesis: Sphagnum Moss and Peat Structure Regulate the Response of CO2 Exchange to Altered Hydrology in a Northern Peatland Ecosystem



Abstract
:1. Introduction
2. Materials and Methods
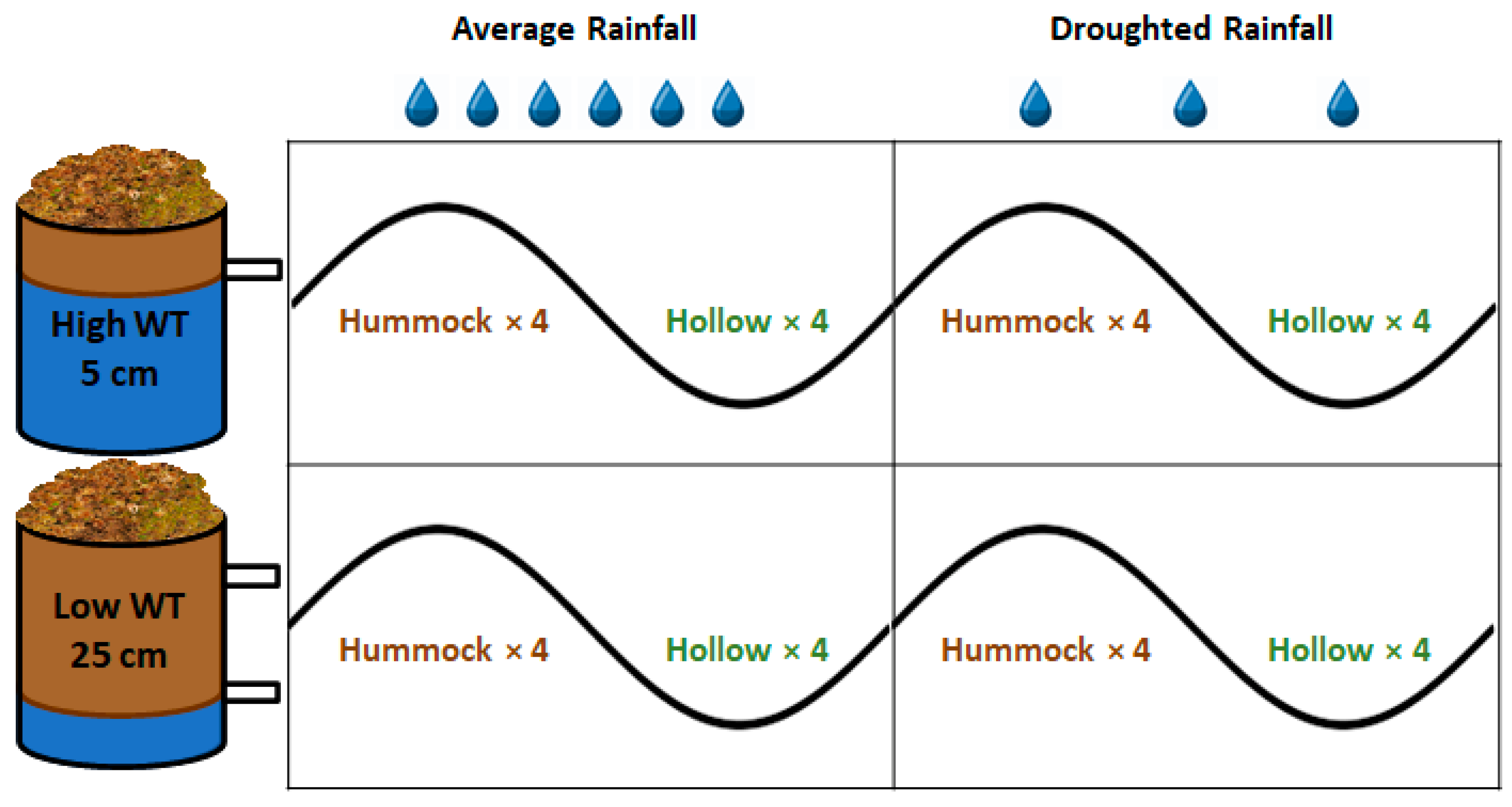
2.1. Peat Harvest and Water Table Treatments
2.2. Flux Measurements
2.3. Spectral Reflectance Measurements
2.4. Sphagnum Characteristics
2.5. Data Analysis
3. Results
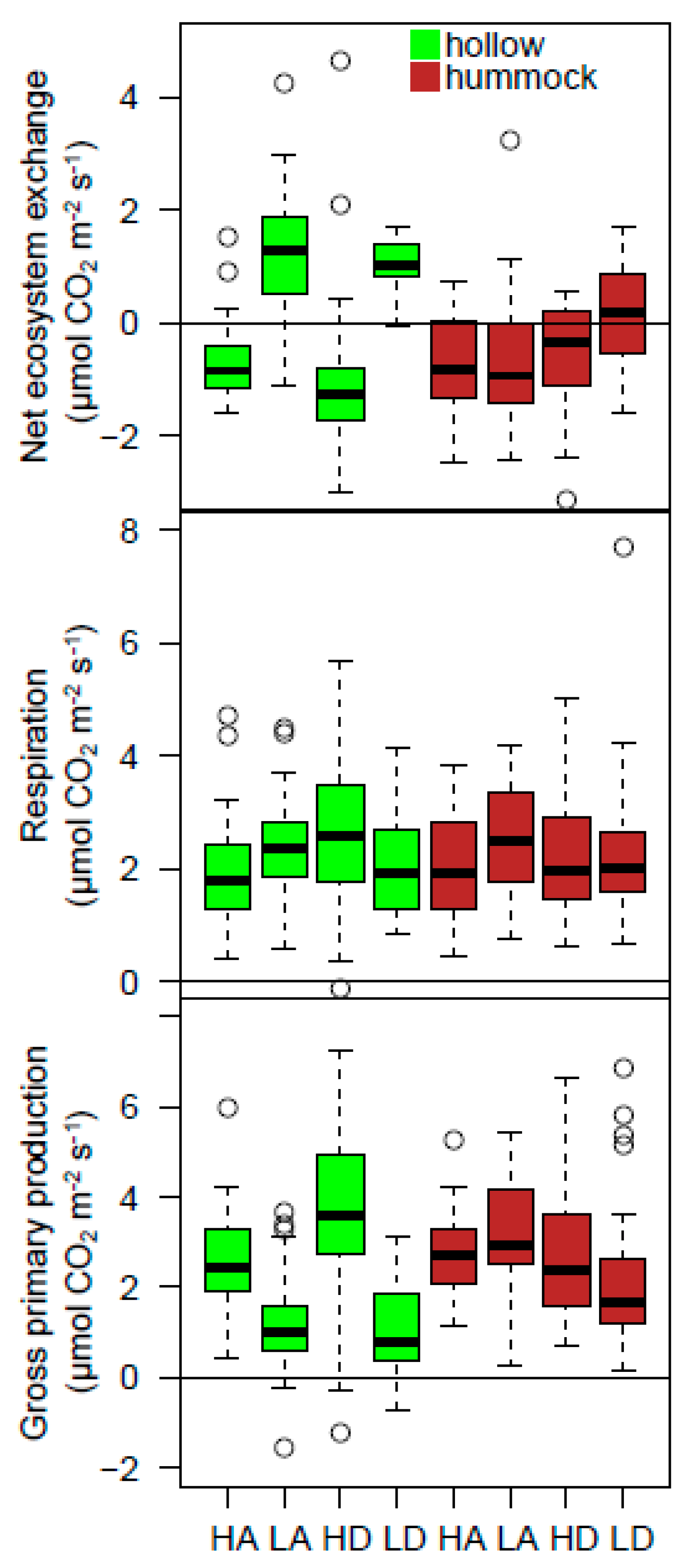
3.1. CO2 Fluxes in Response to Treatments
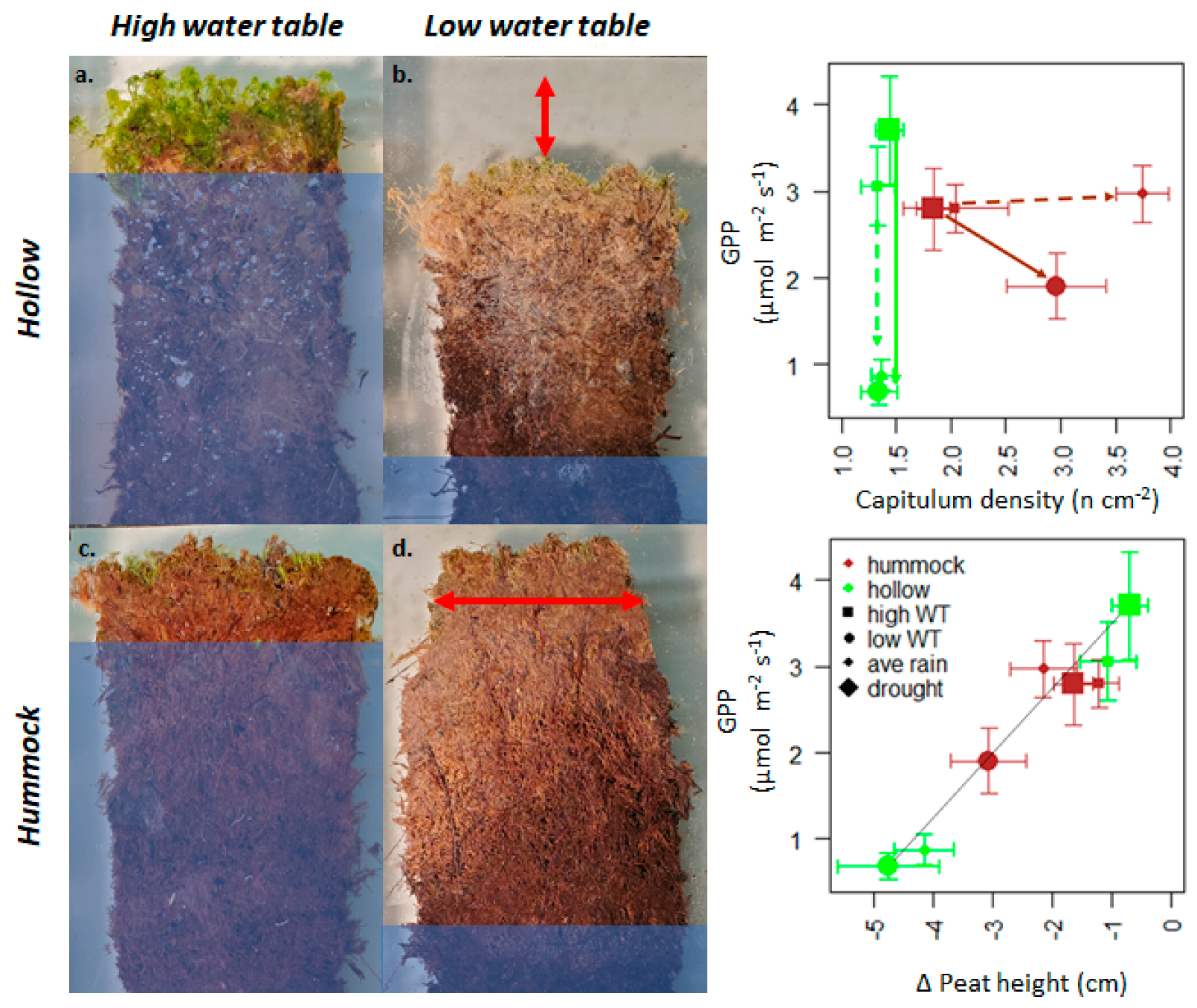
3.2. Sphagnum and Peat Physical Structure in Response to Treatments
4. Discussion
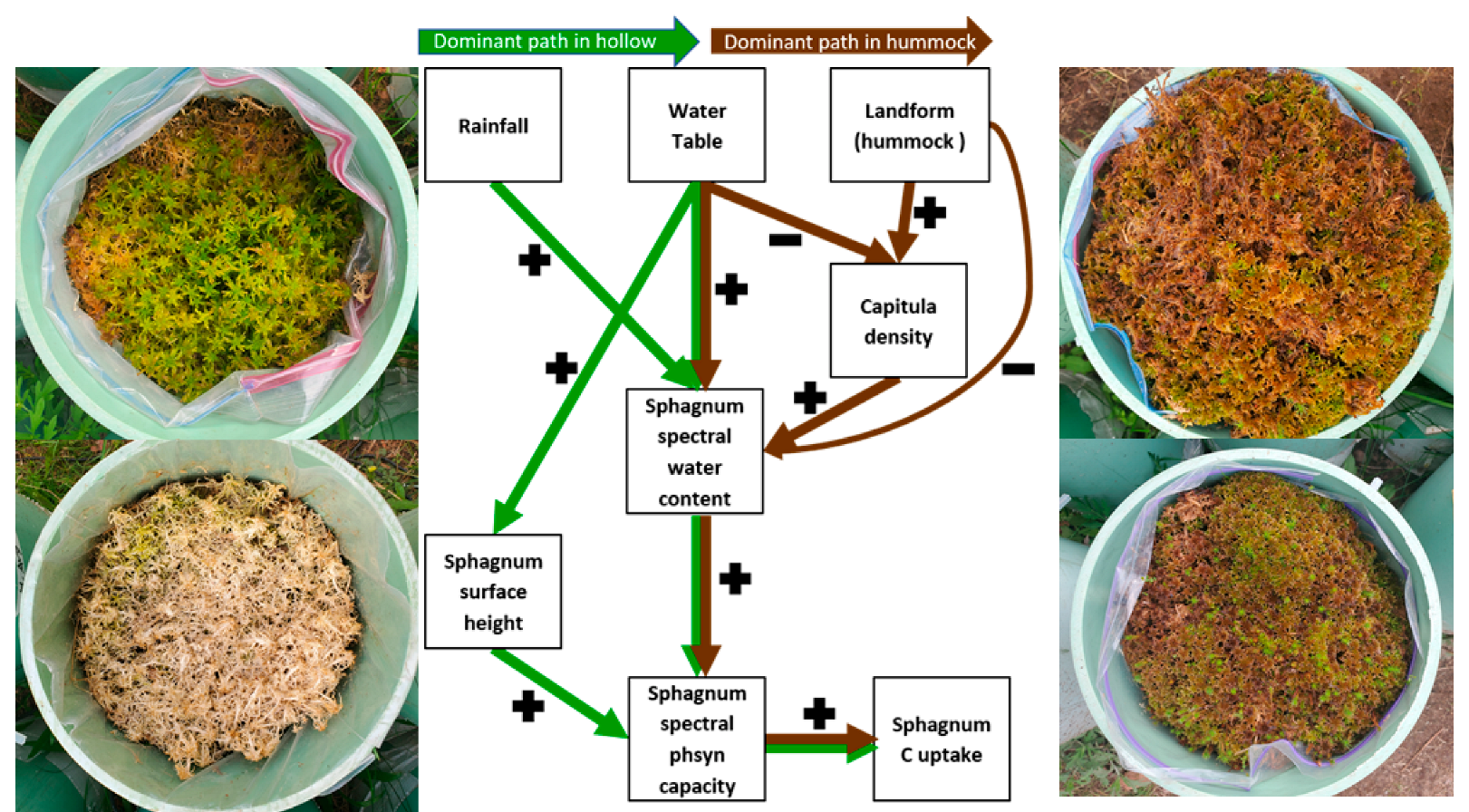
4.1. Sphagnum Trait Mechanisms and Consequences
4.2. Mire-Breathing
4.3. Implications for Peatland Ecosystem Carbon and Water Functions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Loisel, J.; Gallego-Sala, A.V.; Amesbury, M.J.; Magnan, G.; Anshari, G.; Beilman, D.W.; Benavides, J.C.; Blewett, J.; Camill, P.; Charman, D.J.; et al. Expert Assessment of Future Vulnerability of the Global Peatland Carbon Sink. Nat. Clim. Chang. 2021, 11, 70–77. [Google Scholar] [CrossRef]
- Moomaw, W.R.; Chmura, G.L.; Davies, G.T.; Finlayson, C.M.; Middleton, B.A.; Natali, S.M.; Perry, J.E.; Roulet, N.; Sutton-Grier, A.E. Wetlands in a Changing Climate: Science, Policy and Management. Wetlands 2018, 38, 183–205. [Google Scholar] [CrossRef] [Green Version]
- Nichols, J.E.; Peteet, D.M. Rapid Expansion of Northern Peatlands and Doubled Estimate of Carbon Storage. Nat. Geosci. 2019, 12, 917–921. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A Large and Persistent Carbon Sink in the World’s Forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Morris, P.J.; Liu, J.; Holden, J. PEATMAP: Refining Estimates of Global Peatland Distribution Based on a Meta-Analysis. Catena 2018, 160, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Strack, M.; Waddington, J.M.; Tuittila, E.S. Effect of Water Table Drawdown on Northern Peatland Methane Dynamics: Implications for Climate Change. Glob. Biogeochem. Cycles 2004, 18. [Google Scholar] [CrossRef] [Green Version]
- Fenner, N.; Freeman, C. Drought-Induced Carbon Loss in Peatlands. Nat. Geosci. 2011, 4, 895–900. [Google Scholar] [CrossRef]
- Bragazza, L.; Buttler, A.; Robroek, B.J.M.; Albrecht, R.; Zaccone, C.; Jassey, V.E.J.; Signarbieux, C. Persistent High Temperature and Low Precipitation Reduce Peat Carbon Accumulation. Glob. Chang. Biol. 2016, 22, 4114–4123. [Google Scholar] [CrossRef]
- Chivers, M.R.; Turetsky, M.R.; Waddington, J.M.; Harden, J.W.; McGuire, A.D. Effects of Experimental Water Table and Temperature Manipulations on Ecosystem CO2 Fluxes in an Alaskan Rich Fen. Ecosystems 2009, 12, 1329–1342. [Google Scholar] [CrossRef]
- Krause, L.; McCullough, K.J.; Kane, E.S.; Kolka, R.K.; Chimner, R.A.; Lilleskov, E.A. Impacts of Historical Ditching on Peat Volume and Carbon in Northern Minnesota USA Peatlands. J. Environ. Manag. 2021, 296, 113090. [Google Scholar] [CrossRef]
- Laing, C.G.; Granath, G.; Belyea, L.R.; Allton, K.E.; Rydin, H. Tradeoffs and Scaling of Functional Traits in Sphagnum as Drivers of Carbon Cycling in Peatlands. Oikos 2014, 123, 817–828. [Google Scholar] [CrossRef]
- Clymo, R.S.; Hayward, P.M. The Ecology of Sphagnum. In Bryophyte Ecology; Smith, A.J.E., Ed.; Springer: Dordrecht, The Netherlands, 1982. [Google Scholar]
- Thompson, D.K.; Waddington, J.M. Sphagnum under Pressure: Towards an Ecohydrological Approach to Examining Sphagnum Productivity. Ecohydrology 2008, 1, 299–308. [Google Scholar] [CrossRef]
- Dai, A. Drought under Global Warming: A Review. Wiley Interdiscip. Rev. Clim. Chang. 2011, 2, 45–65. [Google Scholar] [CrossRef] [Green Version]
- Rahmstorf, S.; Coumou, D. Increase of Extreme Events in a Warming World. Proc. Natl. Acad. Sci. USA 2011, 108, 17905–17909. [Google Scholar] [CrossRef] [Green Version]
- Mccarter, C.P.R.; Price, J.S. Ecohydrology of Sphagnum Moss Hummocks: Mechanisms of Capitula Water Supply and Simulated Effects of Evaporation. Ecohydrology 2014, 7, 33–44. [Google Scholar] [CrossRef]
- Håkan, R.; Jeglum, J.K. The Biology of Peatlands, 2nd ed.; Håkan, R., Jeglum, J.K., Eds.; Oxford University Press: Oxford, UK, 2013. [Google Scholar]
- Piatkowski, B.T.; Yavitt, J.B.; Turetsky, M.R.; Shaw, A.J. Natural Selection on a Carbon Cycling Trait Drives Ecosystem Engineering by Sphagnum (Peat Moss). Proc. R. Soc. B Biol. Sci. 2021, 88, 20210609. [Google Scholar] [CrossRef]
- Mazziotta, A.; Granath, G.; Rydin, H.; Bengtsson, F.; Norberg, J. Scaling Functional Traits to Ecosystem Processes: Towards a Mechanistic Understanding in Peat Mosses. J. Ecol. 2019, 107, 843–859. [Google Scholar] [CrossRef] [Green Version]
- Robroek, B.J.M.; Schouten, M.G.C.; Limpens, J.; Berendse, F.; Poorter, H. Interactive Effects of Water Table and Precipitation on Net CO2 Assimilation of Three Co-Occurring Sphagnum Mosses Differing in Distribution above the Water Table. Glob. Chang. Biol. 2009, 15, 680–691. [Google Scholar] [CrossRef]
- Almendinger, J.C.; Almendinger, J.E.; Glaser, P.H. Topographic Fluctuations Across a Spring Fen and Raised Bog in the Lost River Peatland, Northern Minnesota. J. Ecol. 1986, 74, 393–401. [Google Scholar] [CrossRef]
- Whittington, P.; Strack, M.; van Haarlem, R. The Influence of Peat Volume Change and Vegetation on the Hydrology of a Kettle-Hole Wetland in Southern Ontario, Canada. Mires Peat 2007, 2, 1–14. [Google Scholar]
- Morris, P.J.; Waddington, J.M.; Benscoter, B.W.; Turetsky, M.R. Conceptual Frameworks in Peatland Ecohydrology: Looking beyond the Two-Layered (Acrotelm-Catotelm) Model. Ecohydrology 2011, 4, 1–14. [Google Scholar] [CrossRef]
- Waddington, J.M.; Morris, P.J.; Kettridge, N.; Granath, G.; Thompson, D.K.; Moore, P.A. Hydrological Feedbacks in Northern Peatlands. Ecohydrology 2015, 8, 113–127. [Google Scholar] [CrossRef]
- Howie, S.A.; Hebda, R.J. Bog Surface Oscillation (Mire Breathing): A Useful Measure in Raised Bog Restoration. Hydrol. Process. 2018, 32, 1518–1530. [Google Scholar] [CrossRef]
- Marshall, C.; Sterk, H.P.; Gilbert, P.J.; Andersen, R.; Bradley, A.V.; Sowter, A.; Marsh, S.; Large, D.J. Multiscale Variability and the Comparison of Ground and Satellite Radar Based Measures of Peatland Surface Motion for Peatland Monitoring. Remote Sens. 2022, 14, 336. [Google Scholar] [CrossRef]
- Kellner, E.; Halldin, S. Water Budget Surface-Layer Water Storage in a Sphagnum Bog in Central Sweden. Hydrol. Process. 2002, 16, 87–103. [Google Scholar] [CrossRef]
- Gallego-Sala, A.V.; Charman, D.J.; Brewer, S.; Page, S.E.; Prentice, I.C.; Friedlingstein, P.; Moreton, S.; Amesbury, M.J.; Beilman, D.W.; Björck, S.; et al. Latitudinal Limits to the Predicted Increase of the Peatland Carbon Sink with Warming. Nat. Clim. Chang. 2018, 8, 907–913. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Ricciuto, D.M.; Thornton, P.E.; Xu, X.; Yuan, F.; Norby, R.J.; Walker, A.P.; Warren, J.M.; Mao, J.; Hanson, P.J.; et al. Extending a Land-Surface Model with Sphagnum Moss to Simulate Responses of a Northern Temperate Bog to Whole Ecosystem Warming and Elevated CO2. Biogeosciences 2021, 18, 467–486. [Google Scholar] [CrossRef]
- Fan, Z.; David Mcguire, A.; Turetsky, M.R.; Harden, J.W.; Michael Waddington, J.; Kane, E.S. The Response of Soil Organic Carbon of a Rich Fen Peatland in Interior Alaska to Projected Climate Change. Glob. Chang. Biol. 2013, 19, 604–620. [Google Scholar] [CrossRef]
- Meingast, K.M.; Falkowski, M.J.; Kane, E.S.; Potvin, L.R.; Benscoter, B.W.; Smith, A.M.S.; Bourgeau-Chavez, L.L.; Miller, M.E. Spectral Detection of Near-Surface Moisture Content and Water-Table Position in Northern Peatland Ecosystems. Remote Sens. Environ. 2014, 152, 536–546. [Google Scholar] [CrossRef]
- Waddington, J.M.; Kellner, E.; Strack, M.; Price, J.S. Differential Peat Deformation, Compressibility, and Water Storage between Peatland Microforms: Implications for Ecosystem Function and Development. Water Resour. Res. 2010, 46. [Google Scholar] [CrossRef]
- Potvin, L.R.; Kane, E.S.; Chimner, R.A.; Kolka, R.K.; Lilleskov, E.A. Effects of Water Table Position and Plant Functional Group on Plant Community, Aboveground Production, and Peat Properties in a Peatland Mesocosm Experiment (PEATcosm). Plant Soil 2015, 387, 277–294. [Google Scholar] [CrossRef]
- Harris, A. Spectral Reflectance and Photosynthetic Properties of Sphagnum Mosses Exposed to Progressive Drought. Ecohydrology 2008, 1, 35–42. [Google Scholar] [CrossRef]
- Lees, K.J.; Lees, K.J.; Artz, R.R.E.; Khomik, M.; Clark, J.M.; Ritson, J.; Hancock, M.H.; Cowie, N.R.; Quaife, T. Using Spectral Indices to Estimate Water Content and GPP in Sphagnum Moss and Other Peatland Vegetation. IEEE Trans. Geosci. Remote Sens. 2020. [Google Scholar] [CrossRef] [Green Version]
- Tucker, C.; O’Neill, A.; Meingast, K.; Bourgeau-Chavez, L.; Lilleskov, E.; Kane, E.S. Spectral Indices of Vegetation Condition and Soil Water Content Reflect Controls on CH4 and CO2 Exchange in Sphagnum-Dominated Northern Peatlands. J. Geophys. Res. Biogeosciences 2022, 127, e2021JG006486. [Google Scholar] [CrossRef]
- Bonham, C.W.; Ahmed, J. Measurements for Terrestrial Vegetation; Wiley: New York, NY, USA, 1989; 338p, ISBN 0471048801. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. ; R Core Team. Nlme: Linear and Nonlinear Mixed Effects Models. R-Project; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Rosseel, Y. Lavaan: An R Package for Structural Equation Modeling. J. Stat. Softw. 2012, 48, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Bradley, A.V.; Andersen, R.; Marshall, C.; Sowter, A.; Large, D.J. Identification of Typical Ecohydrological Behaviours Using InSAR Allows Landscape-Scale Mapping of Peatland Condition. Earth Surf. Dyn. 2022, 10, 261–277. [Google Scholar] [CrossRef]
- Golubev, V.; Whittington, P. Effects of Volume Change on the Unsaturated Hydraulic Conductivity of Sphagnum Moss. J. Hydrol. 2018, 559, 884–894. [Google Scholar] [CrossRef]
- Marshall, C.; Gilbert, P.; Sterk, H.P.; Bradley, A.; Andersen, R.; Parry, L.; Sowter, A.; Marsh, S.; Large, D. Peat Surface Response to the 2018 European Drought Event. Evidence from InSAR and Levelling, Proceedings of the 21st EGU General Assembly, EGU2019, Vienna, Asutria, 7–12 April 2019; Copernicus Publications: Göttingen, Germania, 2019. [Google Scholar]
- Nungesser, M.K. Modelling Microtopography in Boreal Peatlands: Hummocks and Hollows. Ecol. Modell. 2003, 165, 175–207. [Google Scholar] [CrossRef]
- Jassey, V.E.J.; Signarbieux, C. Effects of Climate Warming on Sphagnum Photosynthesis in Peatlands Depend on Peat Moisture and Species-Specific Anatomical Traits. Glob. Chang. Biol. 2019, 25, 3859–3870. [Google Scholar] [CrossRef]
- Jassey, V.E.J.; Reczuga, M.K.; Zielińska, M.; Słowińska, S.; Robroek, B.J.M.; Mariotte, P.; Seppey, C.V.W.; Lara, E.; Barabach, J.; Słowiński, M.; et al. Tipping Point in Plant–Fungal Interactions under Severe Drought Causes Abrupt Rise in Peatland Ecosystem Respiration. Glob. Chang. Biol. 2018, 24, 972–986. [Google Scholar] [CrossRef]
- Laine, A.M.; Mäkiranta, P.; Laiho, R.; Mehtätalo, L.; Penttilä, T.; Korrensalo, A.; Minkkinen, K.; Fritze, H.; Tuittila, E.S. Warming Impacts on Boreal Fen CO2 Exchange under Wet and Dry Conditions. Glob. Chang. Biol. 2019, 25, 1995–2008. [Google Scholar] [CrossRef]
- Wang, H.; Richardson, C.J.; Ho, M. Dual Controls on Carbon Loss during Drought in Peatlands. Nat. Clim. Chang. 2015, 5, 584–587. [Google Scholar] [CrossRef]
- McNeil, P.; Waddington, J.M. Moisture Controls on Sphagnum Growth and CO2 Exchange on a Cutover Bog. J. Appl. Ecol. 2003, 40, 354–367. [Google Scholar] [CrossRef]
- Strack, M.; Price, J.S. Moisture Controls on Carbon Dioxide Dynamics of Peat-Sphagnum Monoliths. Ecohydrology 2009, 2, 34–41. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Response | Factors | Standardized Effect | Z-Value | P > |z|) |
| Capitulum density | Water table | 0.275 | 2.673 | 0.008 |
| Rainfall | 0.036 | 0.354 | 0.723 | |
| Landform | 0.513 | 4.994 | 0 | |
| Surface height | Water table | 0.67 | 7.279 | 0 |
| Rainfall | −0.053 | −0.576 | 0.565 | |
| Landform | −0.113 | −1.225 | 0.221 | |
| Wetness index (WI) | Water table | −0.653 | −5.939 | 0 |
| Rainfall | 0.193 | 2.596 | 0.009 | |
| Capitulum density | 0.257 | 2.727 | 0.006 | |
| Surface height | −0.174 | −1.652 | 0.098 | |
| Landform | −0.387 | −4.411 | 0 | |
| Normalized difference vegetation index (NDVI) | Water table | −0.013 | −0.159 | 0.874 |
| Rainfall | −0.027 | −0.557 | 0.577 | |
| WI | 0.682 | 8.809 | 0 | |
| Capitulum density | 0.118 | 1.933 | 0.053 | |
| Surface height | −0.307 | −4.656 | 0 | |
| Landform | 0.295 | 4.783 | 0 | |
| Gross primary productivity (GPP) | WI | −0.189 | −0.846 | 0.398 |
| NDVI | 0.709 | 3.399 | 0.001 | |
| Landform | −0.021 | −0.162 | 0.871 | |
| Water table | −0.181 | −1.335 | 0.182 | |
| Rainfall | −0.094 | −0.976 | 0.329 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
O’Neill, A.; Tucker, C.; Kane, E.S. Fresh Air for the Mire-Breathing Hypothesis: Sphagnum Moss and Peat Structure Regulate the Response of CO2 Exchange to Altered Hydrology in a Northern Peatland Ecosystem. Water 2022, 14, 3239. https://doi.org/10.3390/w14203239
O’Neill A, Tucker C, Kane ES. Fresh Air for the Mire-Breathing Hypothesis: Sphagnum Moss and Peat Structure Regulate the Response of CO2 Exchange to Altered Hydrology in a Northern Peatland Ecosystem. Water. 2022; 14(20):3239. https://doi.org/10.3390/w14203239
Chicago/Turabian StyleO’Neill, Ally, Colin Tucker, and Evan S. Kane. 2022. "Fresh Air for the Mire-Breathing Hypothesis: Sphagnum Moss and Peat Structure Regulate the Response of CO2 Exchange to Altered Hydrology in a Northern Peatland Ecosystem" Water 14, no. 20: 3239. https://doi.org/10.3390/w14203239
APA StyleO’Neill, A., Tucker, C., & Kane, E. S. (2022). Fresh Air for the Mire-Breathing Hypothesis: Sphagnum Moss and Peat Structure Regulate the Response of CO2 Exchange to Altered Hydrology in a Northern Peatland Ecosystem. Water, 14(20), 3239. https://doi.org/10.3390/w14203239