



Spatial Variation of Soil Organic Carbon from Bamen Bay Mangrove in Southern China

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Investigation and Soil Sampling
2.3. Soil Analysis and Data Analysis
3. Results
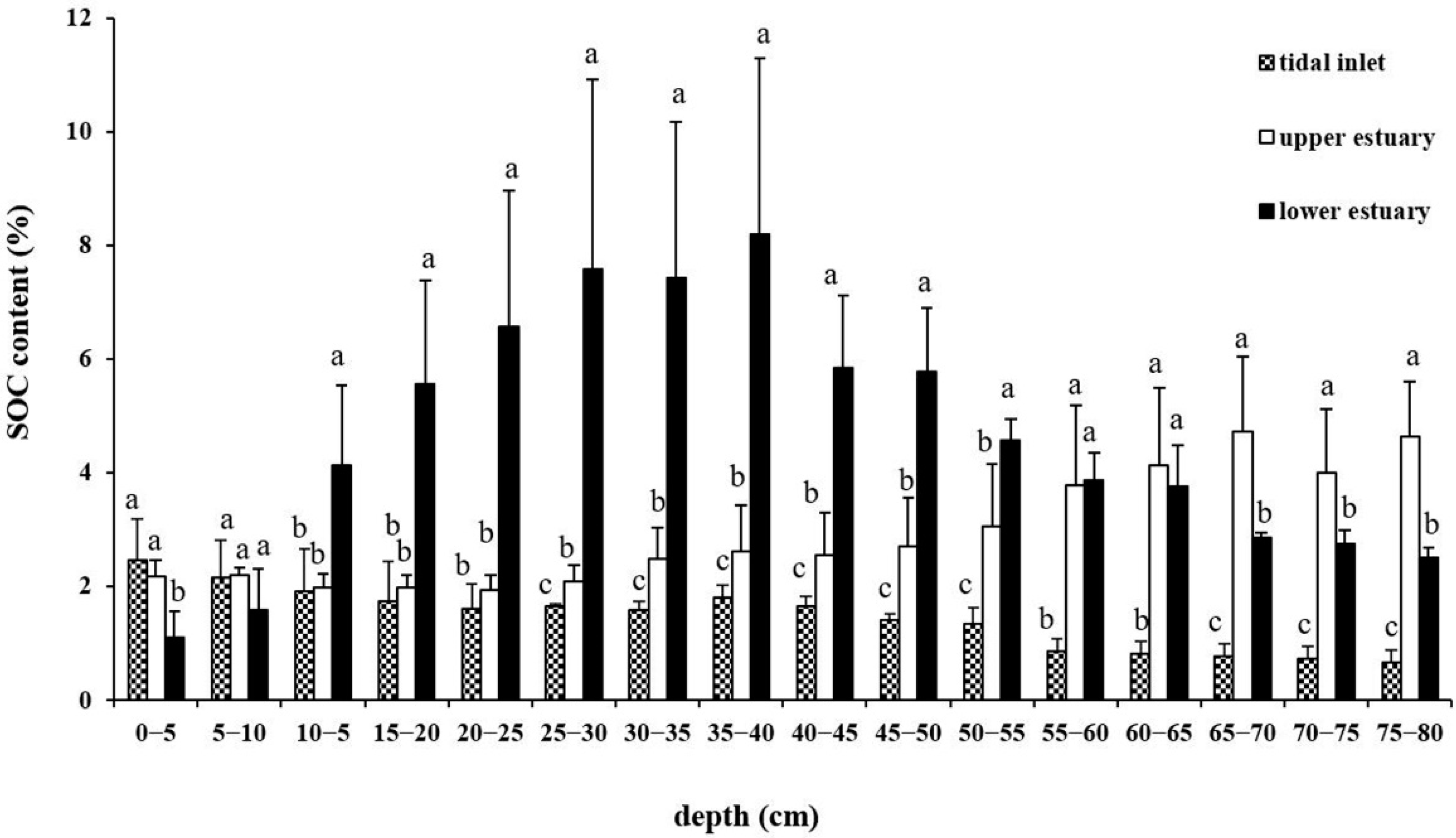
3.1. SOC Content of Three Sites
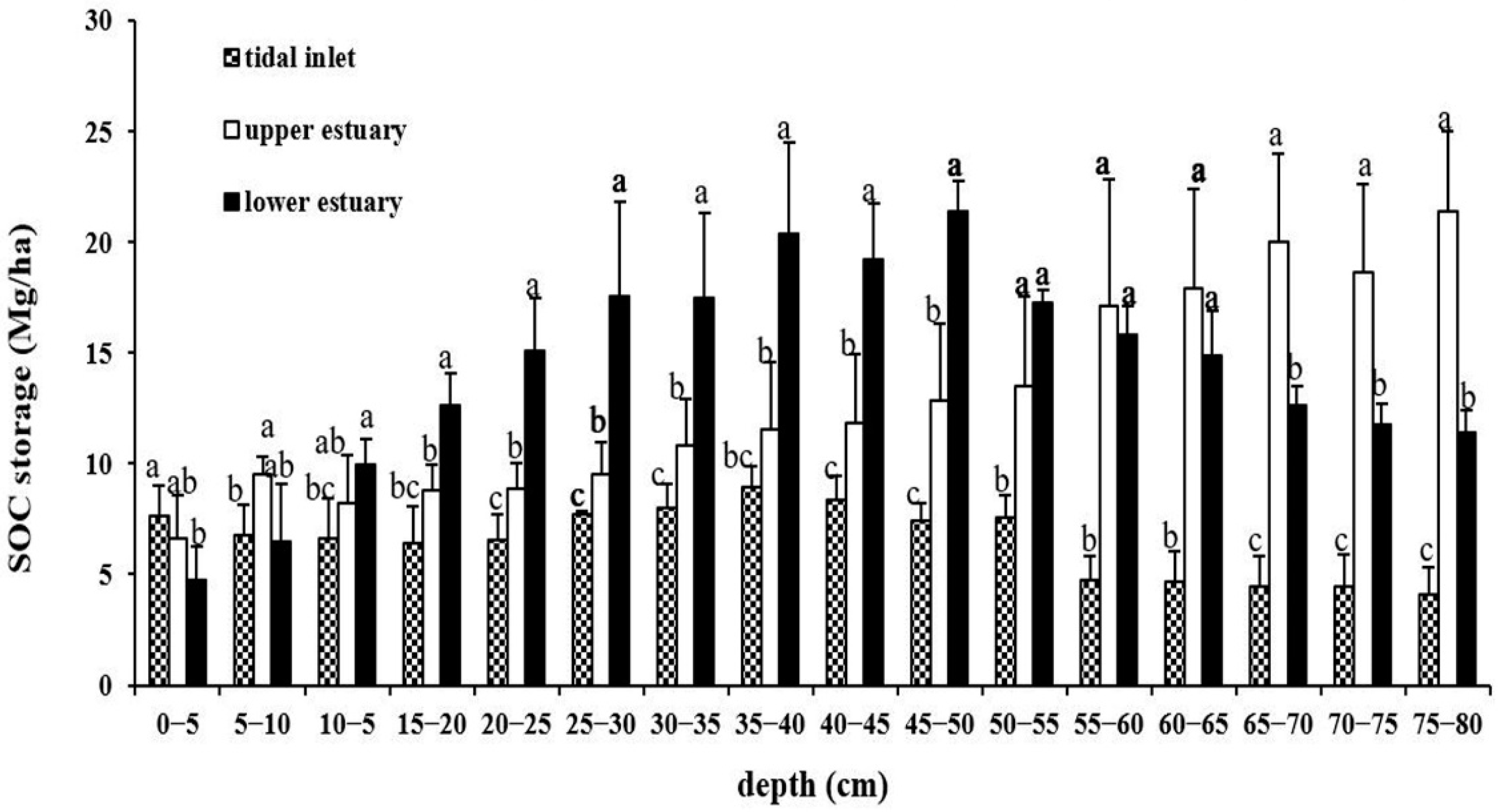
3.2. SOC Storage in Three Geomorphologic Settings
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alongi, D.M. Carbon cycling and storage in mangrove forests. Ann. Rev. Mar. Sci. 2014, 6, 195–219. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zeng, X.; Tam, N.F.Y. Comparing carbon sequestration and stand structure of monoculture and mixed mangrove plantations of Sonneratia caseolaris and S, Apetala in Southern China. For. Ecol. Manag. 2012, 284, 222–229. [Google Scholar] [CrossRef]
- Nong, S.Q.; Yang, X.B.; Li, D.H. Vegetation composition in the mangrove forest nature protection area of Qinglan, China. Plant Sci. J. 2011, 29, 459–466, (In Chinese with English Abstract). [Google Scholar]
- Kauffman, J.B.; Heider, C.; Cole, T.G. Ecosystem carbon stocks of Micronesian mangrove forests. Wetlands 2011, 31, 343–352. [Google Scholar] [CrossRef]
- Sitoe, A.A.; Mandlate, L.J.; Guedes, B.S. Biomass and carbon stocks of Sofala Bay mangrove forests. Forests 2014, 5, 1967–1981. [Google Scholar] [CrossRef]
- Donato, D.C.; Kauffman, J.B.; Murdiyarso, D. Mangroves among the most carbon rich forests in the tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- Giri, C.E.; Ochieng, L.L.; Tieszen, Z. Status and distribution of mangrove forests of the world using earth observation satellite data. Glob. Ecol. Biogeogr. 2011, 20, 154–159. [Google Scholar] [CrossRef]
- Pendleton, L.; Donato, D.C.; Murray, B.C. Estimating global “blue carbon” emissions from conversion and degradation of vegetated coastal ecosystems. PLoS ONE 2012, 7, e43542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atwood, T.B.; Connolly, R.M.; Almahasheer, H.; Carnell, P.E.; Lewis, C.J.E. Global patterns in mangrove soil carbon stocks and losses. Nat. Clim. Chang. 2017, 7, 523–528. [Google Scholar] [CrossRef] [Green Version]
- Rozainah, M.Z.; Nazri, M.N.; Sofawi, A.B.; Hemati, Z.; Juliana, W.A. Estimation of carbon pool in soil, above and below ground vegetation at different types of mangrove forests in Peninsular Malaysia. Mar. Pollut. Bull. 2018, 137, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Alongi, D.M.; Mukhopadhyay, S.K. Contribution of mangroves to coastal carbon cycling in low latitude seas. Agric. For. Meteorol. 2015, 213, 266–272. [Google Scholar] [CrossRef]
- Trevor, G.J.; Harifidy, R.R.; Lalao, R. Ecological variability and carbon stock estimates of mangrove ecosystems in northwestern Madagascar. Forests 2014, 5, 177–205. [Google Scholar]
- Cameron, C.; Kennedy, B.; Tuiwawa, S.; Goldwater, N.; Soapi, K.; Lovelock, C.E. High variance in community structure and ecosystem carbon stocks of Fijian mangroves driven by differences in geomorphology and climate. Environ. Res. 2021, 192, 110213. [Google Scholar] [CrossRef] [PubMed]
- Alongi, D.M. Global significance of mangrove blue carbon in climate change mitigation. Science 2020, 2, 67. [Google Scholar] [CrossRef]
- Bouillon, S.; Borges, A.V.; Castaneda-Moya, E. Mangrove production and carbon sinks, a revision of global budget estimates. Glob. Biogeochem. Cycles 2008, 22, 1–12. [Google Scholar] [CrossRef] [Green Version]
- ShyleshChandran, M.S.; Ravi, A.; John, S.M.; Sivan, S.; Asha, M.S.; Mammen, P.C.; Kumar, K.J.S.; Sruthi, S.N. Ecosystem Carbon Stock of Selected Mangrove Forests of Vypin—Cochin Region, Southwest Coast of India. Wetlands 2020, 40, 2263–2273. [Google Scholar] [CrossRef]
- Bosire, J.O.; Dahdouh-Guebas, F.; Kairo, J.G.; Kazungu, J.M.; Dehairs, F.; Koedam, N. Litter degradation and CN dynamics in reforested mangrove plantations at Gazi Bay, Kenya. Biol. Conserv. 2005, 126, 287–295. [Google Scholar] [CrossRef] [Green Version]
- Werry, J.; Lee, S.Y. Grapsid crabs mediate link between mangrove litter production and estuarine planktonic food chains. Mar. Ecol. Prog. Ser. 2005, 293, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Krauss, K.W.; Doyle, T.W.; Twilley, R.R.; Rivera-Monroy, V.H.; Sullivan, J.K. Evaluating the relative contributions of hydroperiod and soil fertility on growth of south Florida mangroves. Hydrobiologia 2006, 569, 311–324. [Google Scholar] [CrossRef]
- Lugo, A.E.; Medina, E.; McGinley, K. Issues and challenges of mangrove conservation in the Anthropocene. Madera Y Bosques. 2014, 20, 11–38. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Wang, X.; Li, X.; Xin, K.; Yan, Z.; Sun, Y.; Bellerby, R. Distribution Pattern and Influencing Factors for Soil Organic Carbon (SOC) in Mangrove Communities at Dongzhaigang, China. J. Coast. Res. 2018, 34, 434–442. [Google Scholar] [CrossRef]
- Li, S.; Chen, P.; Huang, J.; Hsueh, M.; Hsieh, L.; Lee, C. Factors regulating carbon sinks in mangrove systems. Glob. Chang. Biol. 2018, 24, 4195–4210. [Google Scholar] [CrossRef]
- Wigand, C.; Eagle, M.; Branoff, B.L.; Balogh, S. Recent Carbon Storage and Burial Exceed Historic Rates in the San Juan Bay Estuary Peri-Urban Mangrove Forests (Puerto Rico, United States). Front. For. Glob. Chang. 2021, 4, 676–691. [Google Scholar] [CrossRef] [PubMed]
- Rovai, A.S.; Twilley, R.R.; Riul, P.; Cifuentes-Jara, M.; Manrow-Villalobos, M.; Horta, P.A.; Simonassi, J.C. Global controls on carbon storage in mangrove soils. Nat. Clim. Chang. 2018, 6, 534–538. [Google Scholar] [CrossRef]
- Breithaupt, J.L.; Smoak, J.M.; Smith III, T.J. Organic carbon burial rates in mangrove sediments: Strengthening the global budget. Glob. Biogeochem. Cycles 2012, 26, GB3011. [Google Scholar] [CrossRef]
- Taillardat, P.; Friess, D.A.; Lupascu, M. Mangrove blue carbonstrategies for climate change mitigation are most effective at the national scale. Biol. Lett. 2018, 14, 20180251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adame, M.F.; Neil, D.; Wright, S.F.; Lovelock, C.E. Sedimentation within and among mangrove forests along a gradient of geomorphological settings, Estuar. Coast. Shelf. Sci. 2010, 86, 21–30. [Google Scholar] [CrossRef]
- Adame, M.F.; Santini, N.S.; Tovilla, C.; Castro, L.; Guevara, M. Carbon stocks and soil sequestration rates of tropical riverine wetlands. Biogeosciences 2015, 12, 3805–3818. [Google Scholar] [CrossRef] [Green Version]
- Mcleod, E.; Chmura, G.L.; Bouillon, S.; Salm, R.; Duarte, C.M.; Lovelock, C.E. A blueprint for blue carbon: Toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2. Front. Ecol. Environ. 2011, 9, 552–560. [Google Scholar] [CrossRef] [Green Version]
- Huxham, M.; Langat, J.; Tamooh, F. Decomposition of mangrove roots, effects of location, nutrients, species identity and mix in a Kenyan forest. Estuar. Coast. Shelf. Sci. 2010, 88, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Adame, M.F.; Lovelock, C.E. Carbon and nutrient exchange of mangrove forests with the coastal ocean. Hydrobiologia 2011, 663, 23–50. [Google Scholar] [CrossRef]
- Salmo III, S.G.; Lovelock, C.E.; Duke, N.C. Vegetation and soil characteristics as indicators of restoration trajectories in restored mangroves. Hydrobiologia 2013, 720, 1–18. [Google Scholar] [CrossRef]
- Woodroffe, C.D.; Rogers, K.; Mckee, K.L.; Lovelock, C.E.; Mendelssohn, I.A.; Saintilan, N. Mangrove sedimentation and response to relative sea-level rise. Ann. Rev. Mar. Sci. 2016, 8, 243–266. [Google Scholar] [CrossRef] [Green Version]
- Twilley, R.R.; Chen, R.H.; Hargis, T. Carbon sinks in mangroves and their implications to carbon budget of tropical coastal ecosystems. Water Air Soil Pollut. 1992, 64, 265–288. [Google Scholar] [CrossRef]
- Harris, P.T.; Heap, A.D.; Bryce, S.M. Classification of Australian clastic coastal depositional environments based upon a quantitative analysis of wave, tidal, and river power. J. Sediment. Res. 2002, 72, 858–870. [Google Scholar] [CrossRef]
- Yang, J.; Gao, J.; Liu, B.; Zhang, W. Sediment deposits and organic carbon sequestration along mangrove coasts of the Leizhou peninsula, southern China. Estuar. Coast. Shelf. S. 2014, 136, 3–10. [Google Scholar] [CrossRef]
- Gleason, S.M.; Ewel, K.C. Organic matter dynamics on the forest floor of a Micronesia mangrove forest, an investigation of species composition shifts. Biotropica 2002, 34, 190–198. [Google Scholar] [CrossRef]
- Cerón-Bretón, J.G.; Cerón-Bretón, R.M.; Rangel-MAarrón, M. Determination of carbon sequestration rate in soil of a mangrove forest in Campeche, Mexico. WSEAS Trans. Environ. Dev. 2011, 7, 55–64. [Google Scholar]
- Sherman, R.E.; Fahey, T.J.; Martinez, P. Spatial patterns of biomass and above-ground net primary productivity in a mangrove ecosystem in the Dominican Republic. Ecosystems 2003, 6, 384–398. [Google Scholar] [CrossRef]
- Ren, H.; Chen, H.; Li, A.Z.; Han, W.D. Biomass accumulation and carbon storage of four different aged Sonneratia apetala plantation in Southern China. Plant Soil 2010, 327, 279–291. [Google Scholar] [CrossRef]
- Sun, L. Study on Carbon Storage of Subtropical Broad-Leaved Forest Ecosystem in the Pearl River Delta; Sun Yat-Sen University: Guangzhou, Chinas, 2011. [Google Scholar]
- Zhang, H.; Guan, D.S.; Song, M.W. Biomass and carbon storage of Eucalyptus and Acacia plantations in the Pearl River Delta, South China. For. Ecol. Manag. 2012, 277, 90–97. [Google Scholar] [CrossRef]
- Wang, G.; Guan, D.; Peart, M.R.; Chen, Y.; Peng, Y. Ecosystem carbon stocks of mangrove forest in Yingluo Bay, Guangdong province of South China. For. Ecol. Manag. 2013, 310, 539–546. [Google Scholar] [CrossRef]
- Sebastian, R.; Chacko, J. Distribution of organic carbon in tropical mangrove sediments (Cochin, India). Int. J. Environ. Stud. 2006, 63, 303–311. [Google Scholar] [CrossRef]
- Zhu, Y.; Bourgeois, C.; Lin, G.; Wu, X.; Guo, Z. Spatial characteristic of grain size of surface sediments in mangrove wetlands in Gaoqiao of Zhanjiang, Guangdong Province of South China. Chin. J. Appl. Ecol. 2012, 23, 2114–2120. [Google Scholar]
- Chen, S.; Chen, B.; Chen, G.; Ji, J.; Yu, W.; Liao, J. Higher soil organic carbon sequestration potential at a rehabilitated mangrove comprised of Aegiceras corniculatum compared to Kandelia obovata. Sci. Total Environ. 2021, 752, 142279. [Google Scholar] [CrossRef]
- Ouyang, X.; Lee, S.Y. Improved estimates on global carbon stock and carbon pools in tidal wetlands. Nat. Commun. 2020, 11, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Murdiyarso, D.; Purbopuspito, J.; Kauffman, J.B.; Warren, M.W.; Sasmito, S.D.; Donato, D.C.; Manuri, S.; Krisnawati, H. The potential of Indonesian mangrove forests for global climate change mitigation. Nat. Clim. Chang. 2015. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Bernardino, A.F.; Ferreira, T.O.; Giovannoni, L.R.; Giovannoni, L.R.; Ruiz, F. Carbon stocks of mangroves and salt marshes of the Amazon region, Brazil. Biol. Lett. 2018, 14, 20180208. [Google Scholar] [CrossRef]
- Donato, D.C.; Kauffman, J.B.; Mackenzie, R.A. Whole-island carbon stocks in the tropical Pacific: Implications for mangrove conservation and upland restoration. J. Environ. Manag. 2012, 97, 89–96. [Google Scholar] [CrossRef]
- Eid, E.M.; Arshad, M. Effect of the conversion of mangroves into shrimp farms on carbon stock in the sediment along the southern Red Sea coast, Saudi Arabia. Environ. Res. 2019, 176, 108536. [Google Scholar] [CrossRef]
- Adame, M.F.; Kauffman, J.B.; Medina, I.; Gamboa, J.N.; Torres, O.; Caamal, J.P.; Reza, M.; Jorge, A. Carbon Stocks of Tropical Coastal Wetlands within the Karstic Landscape of the Mexican Caribbean. PLoS ONE 2013, 8, e56569. [Google Scholar] [CrossRef] [Green Version]
- Almahasheer, H.; Serrano, O.; Duarte, C.M.; Arias-Ortiz, A. Low Carbon sink capacity of Red Sea mangroves. Sci. Rep. 2017, 7, 9700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alongi, D.M. Carbon payments for mangrove conservation: Ecosystem constraints and uncertainties of sequestration potential. Environ. Sci. Policy 2011, 14, 462–470. [Google Scholar] [CrossRef]
- Stringer, C.E.; Trettin, C.C.; Zarnoch, S.J.; Tang, W. Carbon stocks of mangroves within the Zambezi River Delta, Mozambique. For. Ecol. Manag. 2015, 354, 139–148. [Google Scholar] [CrossRef]
- Jacotot, A.; Marchand, C.; Rosenheim, B.E.; Domack, E.W. Mangrove sediment carbon stocks along an elevation gradient: Influence of the late Holocene marine regression (New Caledonia). Mar. Geol. 2018, 404, 60–70. [Google Scholar] [CrossRef]
- Schile, L.M.; Kauffman, J.B.; Crooks, S.; Fourqurean, J.W.; Glavan, J. Limits on carbon sequestration in arid blue carbon ecosystems. Ecol. Appl. 2018, 27, 859–874. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Site Name | Longitude and Latitude | Features | Dominant Species |
|---|---|---|---|
| Upper estuary (UE) | 110°47′56.25″, 19°36′59.47″ | More Fresh water, far from the estuary mouth | Sonneratia caseolaris, Lumnitzera racemosa |
| Lower estuary (LE) | 110°47′52.52″, 19°36′7.98″ | Fresh water and tidal water, near to the estuary mouth | Bruguiera sexangula, Ceriops tagal |
| Tidal inlet (TI) | 110°50′20.58″, 19°33′30.90″ | More tidal water, near to the sea | Sonneratia alba |
| Sample Depth (cm) | SOC Storage (Mg/ha) | ||
|---|---|---|---|
| TI | UE | LE | |
| 0–10 | 14.4 A ± 2.8 | 16.2 A ± 2.7 | 11.2 A ± 4.1 |
| 0–20 | 27.4 A ± 6.1 | 33.2 A ± 5.9 | 33.8 A ± 1.9 |
| 0–30 | 41.7 B ± 6.8 | 51.6 AB ± 8.4 | 66.5 A ± 5.0 |
| 0–40 | 58.6 B ± 8.1 | 73.9 B ± 14.0 | 104.4 A ± 12.4 |
| 0–50 | 74.5 B ± 9.8 | 98.6 B ± 19.9 | 145.0 A ± 16.0 |
| 0–60 | 86.8 BC ± 11.3 | 129.2 AB ± 29.7 | 178.1 A ± 17.4 |
| 0–70 | 95.9 BC ± 13.9 | 167.1 AB ± 37.9 | 205.6 A ± 19.0 |
| 0–80 | 104.4 B ± 16.6 | 207.1 A ± 44.8 | 228.8 A ± 19.4 |
| Location | Latitude | C Storage (Mg /ha) | Soil Layer (cm) | Reference |
|---|---|---|---|---|
| Indonesian Archipelago | 00°40′–07°43′ S | 466.3 ± 10.9 | 0–100 | Murdiyarso et al. [48] |
| Mangroves of east of the mouth of Amazon river, Brazil | 00°38′–00°59′ S | 340.0 ± N | >100 | Kauffman et al. [49] |
| Yap and Babeldoab island, Micronesia | 07°21′–09°35′ N | 415.6 ± 23.5 | 0–100 | Kauffman et al. [4] |
| Two island groups of Micronesia (Yap and Palau) | 07°35′–09°33′ N | 692.5± N | 0–100 | Donato et al. [50] |
| The Encrucijada Biosphere Reserve, in Chiapas, Mexico | 14°43′ N | 505.9 ± 72.6 | <100 | Adame et al. [31] |
| southern Red Sea coast of Saudi Arabia | 17°10′–17°48′ N | 292.0 ± 4.0 | 0–100 | Eid et al. [51] |
| Mangrove Nature Reserve of Bamen Bay at Qinglan harbor, China | 19°22′–19°35′ N | 180.1 ± 35.2 | 0–80 | This study |
| Sian Ka’an Biosphere Reserve, Quintana Roo State in the Yucatan Peninsula, Mexico | 19°28′–20°05′ | 489.0 ± 105.5 | 0–100 | Adame et al. [52] |
| Saudi coast, Central Red Sea | 22°21′–23°00′ N | 42.5 ± 5.3 | 0–100 | Almahasheer et al. [53] |
| Dampier, Port Hedland, Mangrove Bay, Bay of Rest, Northern coast of Western Australia | 20°20′–22°19′ | 338.0 ± N | 0–100 | Alongi et al. [54] |
| Zhanjiang Nature Reserve in Yingluo Bay, South China | 20°14′–21°35′ N | 229.1 ± N | 0–100 | Wang et al. [43] |
| The Zambezi River Delta, Mozambique | about 21° | 158.9 ± 27.8 | 0–110 | Stringer et al. [55] |
| Amboa Swamp, New Caledonia | 21°44′ | 283.2± N | 0–100 | Jacotot et al. [56] |
| Aribian Gulf, United Arab Emirates | 24°20′–25°15′ | 143.5 ± N | 0–100 | Schile et al. [57] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, W.; Zhu, Y.; Guo, J.; Pan, X.; Li, J.; Guo, Y.; Guo, J.; Wu, G.; Wang, Y.; Gong, M. Spatial Variation of Soil Organic Carbon from Bamen Bay Mangrove in Southern China. Water 2022, 14, 3278. https://doi.org/10.3390/w14203278
Wen W, Zhu Y, Guo J, Pan X, Li J, Guo Y, Guo J, Wu G, Wang Y, Gong M. Spatial Variation of Soil Organic Carbon from Bamen Bay Mangrove in Southern China. Water. 2022; 14(20):3278. https://doi.org/10.3390/w14203278
Chicago/Turabian StyleWen, Wanyu, Yaojun Zhu, Jia Guo, Xu Pan, Jing Li, Yanru Guo, Julan Guo, Gaojie Wu, Yuhang Wang, and Minghao Gong. 2022. "Spatial Variation of Soil Organic Carbon from Bamen Bay Mangrove in Southern China" Water 14, no. 20: 3278. https://doi.org/10.3390/w14203278
APA StyleWen, W., Zhu, Y., Guo, J., Pan, X., Li, J., Guo, Y., Guo, J., Wu, G., Wang, Y., & Gong, M. (2022). Spatial Variation of Soil Organic Carbon from Bamen Bay Mangrove in Southern China. Water, 14(20), 3278. https://doi.org/10.3390/w14203278