
A Review of the Ecological and Biogeographic Differences of Amazonian Floodplain Forests

Abstract
:1. Introduction
2. The Physical Setting of Amazonian Floodplains
3. Floristic Differences between Várzea and Igapó
3.1. Methods Applied in the Floristic Comparison
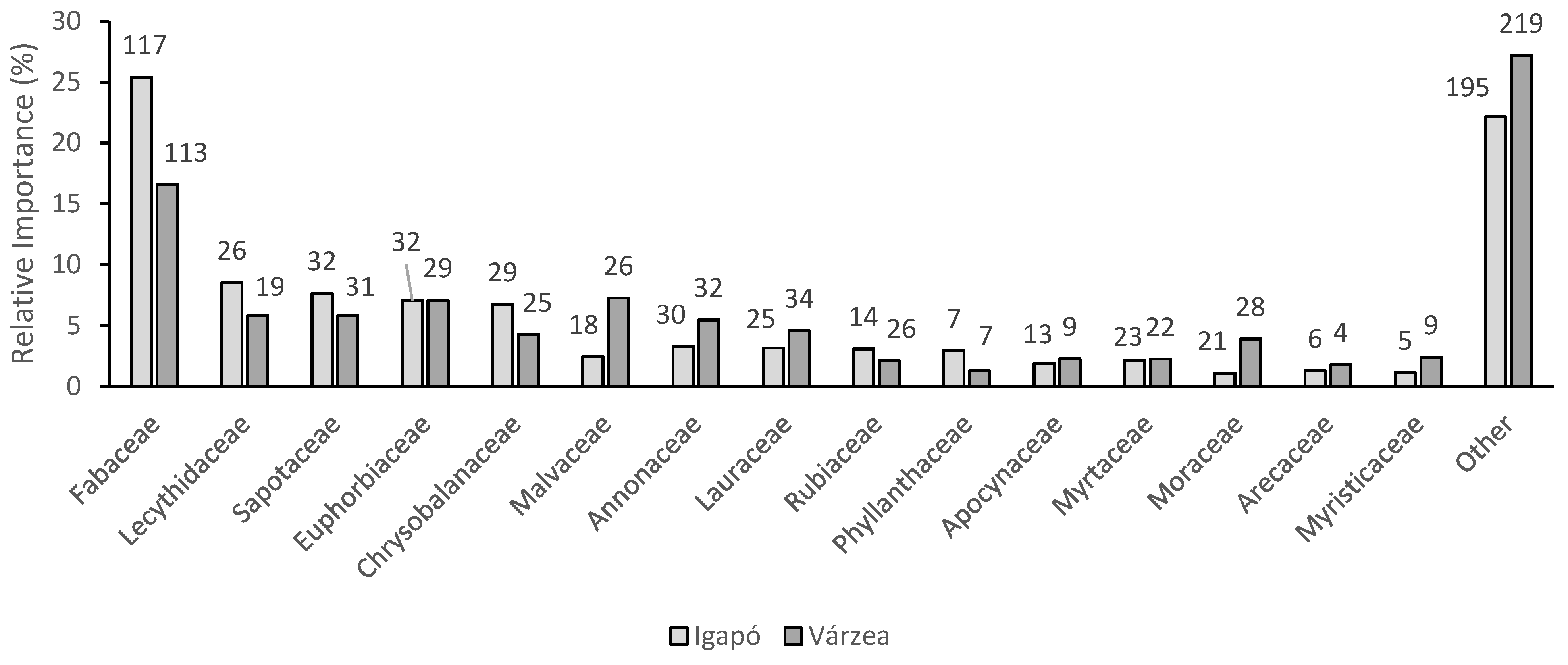
3.2. Results of the Floristic Comparison
4. Ecosystem Specificity of Várzea and Igapó
4.1. The Role of Flooding
4.2. The Role of Nutrients

{kind=link}
| Igapó | Várzea | |
|---|---|---|
| Area 1 (km2) | 302,000 | 456,000 |
| River origin 2,3 | Guiana and Central Brazilian Shields, pre-Cambrian and Paleozoic formations | Andes and Andean foothills |
| Ecosystem characteristics | ||
| Habitat stability | Stable during centuries to millennia 4 | Instable through sedimentation and erosion processes and channel migration 5 |
| Habitat diversity 5 | Low, mostly defined by flood-levels | High through small-scale hydrogeomorphic landforms and different flood levels |
| Fertility 6 | Low (intermediate in clear-water rivers) | High |
| Soil type 7 | Mostly arenosols with C content ~50% | Mostly different grained (sand-clay) Fluvisols and Inceptisols with C content ~40% |
| Risk of edaphic drought 8 | High, and very high at the highest flood levels | Low, but intermediate at the highest flood levels |
| Risk of fire 9 | High at the highest flood levels | Low |
| Life-history traits | ||
| Strategy 10 | Resource conservation | Resource acquisition |
| Leaves 11 | Small, hard, sclerophyllic, protected against herbivores | Soft, big, deciduous |
| Wood density 12 | Higher | Lower |
| Roots 13 | Surface or top 40 cm | 40–70 cm |
| Seeds 14 | Big and heavy, attached to seedlings | Small and light |
| Biogeographic connections | ||
| Guiana and Central Brazilian Shields, Amazonian white-sand forests and savannas (campinaranas), Cerrado 15 | Western Amazon, Andes, Orinoco floodplains, Central American rainforest 16,17 | |
4.3. The Role of Geomorphic Disturbance and Habitat Stability
4.4. The Role of Drought
4.5. The Role of Fire
4.6. The Role of Microclimates
5. Biogeographic Patterns
5.1. Endemism and Habitat Specialization in Amazonian Floodplains
5.2. Floristic Links of Floodplain Floras within Amazonia and Beyond
5.3. Floodplain Ecosystems through Time
5.4. Floodplain Refugia
6. Conservation Implications
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Wittmann, F.; Schöngart, J.; Montero, J.C.; Motzer, T.; Junk, W.J.; Piedade, M.T.F.; Queiroz, H.L.; Worbes, M. Tree species composition and diversity gradients in white-water forests across the Amazon basin. J. Biogeogr. 2006, 33, 1334–1347. [Google Scholar] [CrossRef]
- Malhi, Y.; Wood, D.; Baker, T.R.; Wright, J.; Phillips, O.L.; Cochrane, T.; Meir, P.; Chave, J.; Almeida, S.; Arroyo, L.; et al. The regional variation of aboveground live biomass in old-growth Amazonian forests. Glob. Change Biol. 2006, 12, 1107–1138. [Google Scholar] [CrossRef]
- Schöngart, J.; Wittmann, F.; Worbes, M. Biomass and Net Primary Production of Central Amazonian Floodplain Forests. In Amazonian Floodplain Forests: Ecophysiology, Biodiversity and Sustainable Management. Ecological Studies 210; Junk, W.J., Piedade, M.T.F., Wittmann, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 347–388. [Google Scholar]
- Junk, W.J.; Piedade, M.T.F.; Schöngart, J.; Cohn-Haft, M.; Adeney, J.M.; Wittmann, F. A classification of major naturally-occurring Amazonian lowland wetlands. Wetlands 2011, 31, 623–640. [Google Scholar] [CrossRef]
- Wittmann, F.; Householder, E.; Piedade, M.T.F.; de Assis, R.L.; Schöngart, J.; Parolin, P.; Junk, W.J. Habitat specificity, endemism and the neotropical distribution of Amazonian white-water floodplain trees. Ecography 2013, 36, 690–707. [Google Scholar] [CrossRef]
- Sculthorpe, C.D. The Biology of Aquatic Vascular Plants; Koeltz Scientific Books: Königstein, Germany, 1985. [Google Scholar]
- Greb, F.G.; DiMichele, W.A.; Gastaldo, R.A. Evolution and importance of wetlands in earth history. In WetlandsThrough Time: Geological Society of America Special Paper 399; Greb, F.G., DiMichele, R.A., Eds.; Geological Society of America: Boulder, CO, USA, 2006; pp. 1–40. [Google Scholar]
- Sedell, J.R.; Reeves, G.H.; Hauer, F.R.; Stanford, J.A.; Hawkins, C.P. Role of refugia in recovery from disturbances: Modern fragmented and disconnected river systems. Environ. Manag. 1990, 14, 711–724. [Google Scholar] [CrossRef]
- Sioli, H. Beiträge zur regionalen Limnologie des Amazonasgebietes. Arch. Hydrobiol. 1954, 45, 267–283. [Google Scholar]
- Prance, G.T. Notes on the vegetation of Amazonia. III. Terminology of Amazonian forest types subjected to inundation. Brittonia 1979, 31, 26–38. [Google Scholar] [CrossRef]
- Furch, K.; Junk, W.J. Physicochemical conditions in floodplains. In The Central Amazon Floodplain: Ecology of a Pulsing System. Ecological Studies 126; Junk, W.J., Ed.; Springer: Berlin/Heidelberg, Germany, 1997; pp. 69–108. [Google Scholar]
- Kubitzki, K. The ecogeographical differentiation of Amazonian inundation forests. Plant Syst. Evol. 1989, 162, 285–304. [Google Scholar] [CrossRef]
- Melack, J.M.; Hess, L.L. Remote sensing of the distribution and extent of wetlands in the Amazon basin. In Amazonian Floodplain Forest: Ecophysiology, Biodiversity and Sustainable Management. Ecological Studies 210; Junk, W.J., Piedade, M.T.F., Wittmann, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 27–42. [Google Scholar]
- Junk, W.J.; Bayley, P.B.; Sparks, R.E. The Flood Pulse Concept in River Floodplain Systems. In Proceedings of the International Large River Symposium, Honey Harbour, ON, Canada, 14–21 September 1986; Dodge, D.P., Ed.; Canadian Special Publication of Fisheries Aquatic Sciences: Ottawa, ON, Canada, 1989; pp. 110–127. [Google Scholar]
- Fassoni-Andrade, A.C.; Fleischmann, A.S.; Papa, F.; de Paiva, R.C.D.; Wongchuig, S.; Melack, J.M.; Moreira, A.A.; Paris, A.; Ruhoff, A.; Barbosa, C.; et al. Amazon hydrology from space: Advances and future challenges. Rev. Geophys. 2021, 59, e2020RG000728. [Google Scholar] [CrossRef]
- Wittmann, F.; Junk, W.J. The Amazon River basin. In The Wetland Book II: Distribution, Description and Conservation; Finlayson, C.M., Milton, G.R., Prentice, R.C., Davidson, N.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 1–20. [Google Scholar]
- Welcomme, R.L. Fisheries Ecology of Floodplain Rivers; Longman: London, UK, 1979. [Google Scholar]
- Goulding, M. The Fishes and the Forest: Explorations in the Amazonian Natural History; University of California Press: Berkeley, CA, USA, 1980. [Google Scholar]
- Lowe-McConnell, R.H. Ecological Studies in Tropical Fish Communities; Cambridge University Press: Cambridge, UK, 1987. [Google Scholar]
- Adis, J.; Junk, W.J. Terrestrial invertebrates inhabiting lowland river floodplains of Central Amazonian and Central Europe: A review. Freshw. Biol. 2022, 47, 711–731. [Google Scholar] [CrossRef]
- Correa, S.B.; Costa-Pereira, R.; Fleming, T.; Goulding, M.; Anderson, J.T. Neotropical fish-fruit interactions: Eco-evolutionary dynamics and conservation. Biol. Rev. 2015, 90, 1263–1278. [Google Scholar] [CrossRef] [PubMed]
- Junk, W.J.; da Silva, V.M.F. Mammals, reptiles and amphibians. In The Central Amazon Floodplain: Ecology of a Pulsing System. Ecological Studies 126; Junk, W.J., Ed.; Springer: Berlin/Heidelberg, Germany, 1997; pp. 409–417. [Google Scholar]
- Tomas, W.M.; Cáceres, N.C.; Nunes, A.P.; Fischer, E.; Mourão, G.; Campos, Z. Mammals in the Pantanal wetland, Brazil. In The Pantanal: Ecology, Biodiversity and Sustainable Management of a Large Neotropical Seasonal Wetland; Junk, W.J., da Silva, C.J., Eds.; Pensoft: Sofia, Bulgaria, 2011; pp. 565–597. [Google Scholar]
- Ramalho, E.E.; Main, M.B.; Alvarenga, G.C.; Oliveira-Santos, L.G.R. Walking on water: The unexpected evolution of arboreal lifestyle in a large top predator in the Amazon flooded forests. Ecology 2021, 102, e03286. [Google Scholar] [CrossRef] [PubMed]
- Junk, W.J. Flood tolerance and tree distribution in Central Amazonian floodplains. In Tropical Forests: Botanical Dynamics, Speciation and Diversity; Nielsen, L.B., Nielsen, I.C., Balslev, H., Eds.; Academic Press: London, UK, 1989; pp. 47–64. [Google Scholar]
- Parolin, P.; De Simone, O.; Haase, K.; Waldhoff, D.; Rottenberger, S.; Kuhn, U.; Kesselmeier, J.; Kleiss, B.; Schmidt, W.; Piedade, M.T.F.; et al. Central Amazon floodplain forests: Tree survival in a pulsing system. Bot. Rev. 2004, 70, 357–380. [Google Scholar] [CrossRef]
- Parolin, P. Submerged in darkness: Adaptations to prolonged submergence by woody species of the Amazonian floodplains. Ann. Bot. 2009, 103, 359–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rull, V. Palaeofloristics and palaeovegetacional changes across the Paleocene-Eocene boundary in northern South America. Rev. Palaeobot. Palynol. 1999, 107, 83–95. [Google Scholar] [CrossRef]
- Burnham, R.J.; Graham, A. The history of neotropical vegetation: New developments and status. Ann. Mo. Bot. Gard. 1999, 86, 546–589. [Google Scholar] [CrossRef]
- Burnham, R.J.; Johnson, K.R. South-American palaeobotany and the origins of neotropical rainforests. Philos. Trans. R. Soc. Lond. B 2004, 359, 1595–1610. [Google Scholar] [CrossRef] [Green Version]
- Shephard, G.E.; Müller, R.D.; Liu, L.; Gurnis, M. Miocene drainage reversal of the Amazon River driven by plate-mantle interaction. Nat. Geosci. 2010, 3, 870–875. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, J.; Hoorn, C.; van der Veen, P.; Soares, E. Late Miocene onset of the Amazon River and the Amazon deep-sea fan: Evidence from the Foz do Amazonas Basin. Geology 2010, 37, 619–622. [Google Scholar] [CrossRef]
- Hoorn, C.; Wesselingh, F.P.; Ter Steege, H.; Bermudez, M.A.; Mora, A.; Sevink, J.; Sanmartín, I.; Sanchez-Messeguer, A.; Anderson, C.L.; Figueiredo, J.P.; et al. Amazonia through time: Andean uplift, climate change, landscape evolution and biodiversity. Science 2010, 330, 927–931. [Google Scholar] [CrossRef] [Green Version]
- Wittmann, F.; Householder, J.E. Why rivers make the difference: A review on the phytogeography of forested floodplains in the Amazon basin. In Forest Structure, Functions and Dynamics in Western Amazonia; Myster, R.W., Ed.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2017; pp. 125–144. [Google Scholar]
- Gibbs, R.J. Amazon River: Environmental factors that control its dissolved and suspended load. Science 1967, 156, 1734–1737. [Google Scholar] [CrossRef] [PubMed]
- Junk, W.J.; Wittmann, F.; Schöngart, J.; Piedade, M.T.F. A classification of the major habitats of Amazonian black-water river floodplains and a comparison with their white-water counterparts. Wetl. Ecol. Manag. 2015, 23, 677–693. [Google Scholar] [CrossRef]
- Ter Steege, H.; Pitman, N.; Sabatier, D.; Baraloto, C.; Salomão, R.P.; Guevara, J.E.; Phillips, O.L.; Castilho, C.V.; Magnusson, W.E.; Molino, J.-F.; et al. Hyper-dominance in the Amazonian tree flora. Science 2013, 342, 325–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terborgh, J.; Andresen, E. The composition of Amazonian forests: Patterns at local and regional scales. J. Trop. Ecol. 1998, 14, 645–664. [Google Scholar] [CrossRef]
- Ríos-Villamizar, E.A.; Adeney, J.M.; Piedade, M.T.F.; Junk, W.J. New insights on the classification of major Amazonian river water types. Sustain. Water Resour. Manag. 2020, 6, 83. [Google Scholar] [CrossRef]
- Assis RL de Haugaasen, T.; Schöngart, J.; Montero, J.C.; Piedade, M.T.F.; Wittmann, F. Patterns of tree diversity and composition in Amazonian floodplain paleo-várzea forest. J. Veg. Sci. 2015, 26, 312–322. [Google Scholar] [CrossRef]
- Assis RL de Wittmann, F.; Luize, B.G.; Haugaasen, T. Patterns of floristic diversity and composition in floodplain forests across four Southern Amazon River tributaries, Brazil. Flora 2017, 229, 124–140. [Google Scholar] [CrossRef] [Green Version]
- Wittmann, F.; Schöngart, J.; Junk, W.J. Phytogeography, species diversity, community structure and dynamics of Amazonian floodplain forests. In Amazonian Floodplain Forests: Ecophysiology, Biodiversity and Sustainable Management. Ecological Studies 210; Junk, W.J., Piedade, M.T.F., Wittmann, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 61–104. [Google Scholar]
- Householder, J.E.; Schöngart, J.; Piedade, M.T.F.; Junk, W.J.; Ter Steege, H.; Montero, J.C.; de Assis, R.L.; de Aguiar, D.P.P.; Pombo, M.M.; Quaresma, A.C.; et al. Modelling the ecological responses of tree species to the flood pulse of the Amazon Negro River floodplains. Front. Ecol. Evol. 2021, 9, 628606. [Google Scholar] [CrossRef]
- Lobo, G.S.; Wittmann, F.; Piedade, M.T.F. Response of blackwater floodplain (igapo) forests to flood pulse regulation in a dammed Amazonian river. For. Ecol. Manag. 2019, 434, 110–118. [Google Scholar] [CrossRef]
- Targhetta, N.; Kesselmeier, J.; Wittmann, F. Effects of the hydroedaphic gradient on tree species composition and aboveground wood biomass of oligotrophic forest ecosystems in the central Amazon basin. Folia Geobot. 2015, 50, 185–205. [Google Scholar] [CrossRef]
- Montero, J.C.; Piedade, M.T.F.; Wittmann, F. Floristic variation across 600 km of inundation forests (Igapó) along the Negro River, central Amazonia. Hydrobiologia 2014, 729, 229–246. [Google Scholar] [CrossRef]
- Wittmann, F.; Anhuf, D.; Junk, W.J. Tree species distribution and community structure of central Amazonian várzea forests by remote-sensing techniques. J. Trop. Ecol. 2002, 18, 805–820. [Google Scholar] [CrossRef]
- Schöngart, J. Dendrochronologische Untersuchungen in Überschwemmungswäldern der Várzea Zentralamazoniens; Böhnel, H., Tiessen, H., Weidelt, H.J., Eds.; Göttinger Beiträge zur Land- und Forstwirtschaft in den Tropen und Subtropen, Erich Goltze Göttingen: Göttingen, Germany, 2003; Volume 149, pp. 1–257. [Google Scholar]
- Curtis, J.T.; McIntosh, R.P. An upland forest continuum in the prairie-forest border region of Wisconsin. Ecology 1951, 32, 476–496. [Google Scholar] [CrossRef]
- Sørensen, T. A method of establishing groups of equal amplitude in plant sociology based on similarity of species content and its application to analyses of the vegetation on Danish commons. Det K. Dan. Vidensk. Selsk. Biol. Skr. 1948, 5, 1–34. [Google Scholar]
- Ferreira, L.V. Effects of the duration of flooding on species richness and floristic composition in three hectares in the Jaú National Park in floodplain forests in central Amazonia. Biodivers. Conserv. 1997, 6, 1353–1363. [Google Scholar] [CrossRef]
- Piedade, M.T.F.; Junk, W.J.; Adis, J.; Parolin, P. Ecologia, zonação e colonização da vegetação arbórea das Ilhas Anavilhanas. Pesqui. Bot. 2005, 56, 17–144. [Google Scholar]
- Wittmann, F.; Junk, W.J.; Piedade, M.T.F. The várzea forests in Amazonia: Flooding and the highly dynamic geomorphology interact with natural forest succession. For. Ecol. Manag. 2004, 196, 199–212. [Google Scholar] [CrossRef]
- Junk, W.J.; Furch, K. The physical and chemical properties of Amazonian waters and their relationship with the biota. In Amazonia: Key Environments; Prance, G.T., Lovejoy, T., Eds.; Pergamon Press: Oxford, UK, 1985; pp. 3–17. [Google Scholar]
- Crawford, R.M.M. The anaerobic retreat in Studies in Plant Survival: Ecological case histories of plant adaptation to adversity. In Studies in Ecology 11; Crawfoed, R.M.M., Ed.; Blackwell Scientific Publications: Oxford, UK, 1989; pp. 105–129. [Google Scholar]
- Crawford, R.M.M. Oxygen availability as an ecological limit to plant distribution. Adv. Ecol. Res. 1992, 223, 93–185. [Google Scholar]
- Armstrong, W.; Brändle, R.A.; Jackson, M.B. Mechanisms of flood tolerance in plants. Acta Bot. Neerl. 1994, 43, 307–358. [Google Scholar] [CrossRef]
- Visser, E.J.W.; Voesenek, L.A.C.J.; Vatapetian, B.B.; Jackson, M.B. Flooding and plant growth. Ann. Bot. 2003, 91, 107–109. [Google Scholar] [CrossRef] [Green Version]
- Joly, C.A.; Crawford, R.M.M. Variation in tolerance and metabolic responses to flooding in some tropical trees. J. Exp. Bot. 1982, 33, 799–809. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Pallardy, S.G. Acclimation and adaptive responses of woody plants to environmental stresses. Bot. Rev. 2002, 68, 270–334. [Google Scholar] [CrossRef]
- Parolin, P.; Lucas, C.; Piedade, M.T.F.; Wittmann, F. Drought responses of flood-tolerant trees in Amazonian floodplains. Ann. Bot. 2010, 105, 129–139. [Google Scholar] [CrossRef] [Green Version]
- Jackson, M.B.; Colmer, T.D. Response and adaptation by plants to flooding stress. Ann. Bot. 2005, 96, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Schlüter, U.-B.; Furch, B.; Joly, C.A. Physiological and anatomical adaptations by joung Astrocaryum jauari Mart. (Arecaceae) in periodically inundated biotopes of Central Amazonia. Biotropica 1993, 25, 384–396. [Google Scholar] [CrossRef]
- Worbes, M. The forest ecosystem of the floodplains. In The Central Amazon Floodplain: Ecology of a Pulsing System. Ecological Studies 126; Junk, W.J., Ed.; Springer: Berlin/Heidelberg, Germany, 1997; pp. 223–266. [Google Scholar]
- Larcher, W. Ökophysiologie der Pflanzen: Leben, Leistung und Streßbewältigung der Pflanzen in Ihrer Umwelt; 6. Aufl.; Ulmer, UTB für Wissenschaft: Stuttgart, Germany, 2001. [Google Scholar]
- De Simone, O.; Haase, K.; Müller, E.; Junk, W.J.; Gonsior, G.; Schmidt, W. Impact of root morphology on metabolism and oxygen distribution in roots and rhizosphere from two Central Amazon floodplain tree species. Funct. Plant Biol. 2002, 29, 1025–1035. [Google Scholar] [CrossRef]
- Parolin, P.; Oliveira, A.C.; Piedade, M.T.F.; Wittmann, F.; Junk, W.J. Pioneer trees in Amazonian floodplains: Key-species form monospecific stands in different habitats. Folia Geobot. 2002, 37, 225–238. [Google Scholar] [CrossRef]
- Wittmann, F.; Parolin, P. Aboveground roots in Amazonian floodplain trees. Biotropica 2005, 37, 609–619. [Google Scholar] [CrossRef]
- Piedade, M.T.F.; Ferreira, C.S.; Oliveira Wittmann, A.; Buckeridge, M.; Parolin, P. Biochemistry of Amazonian Floodplain Trees. In Amazonian Floodplain Forests: Ecophysiology, Biodiversity and Sustainable Management. Ecological Studies 210; Junk, W.J., Piedade, M.T.F., Wittmann, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 123–134. [Google Scholar]
- Oliveira Wittmann, A.; Lopes, A.; Conserva, A.S.; Wittmann, F.; Piedade, M.T.F. Seed germination and seedling establishment of Amazonian floodplain trees. In Amazonian Floodplain Forests: Ecophysiology, Biodiversity and Sustainable Management. Ecological Studies 210; Junk, W.J., Piedade, M.T.F., Wittmann, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 259–280. [Google Scholar]
- Parolin, P. Submergence tolerance versus escape from submergence: Two strategies of seedling establishment in Amazonian floodplains. Environ. Exp. Bot. 2002, 48, 177–186. [Google Scholar] [CrossRef]
- Voesenek, L.A.C.J.; Benschop, J.J.; Bou, J.; Cox, M.C.H.; Groeneveld, H.W.; Millenaar, F.F.; Vreeburg, R.A.M.; Peeters, A.J.M. Interactions between plant hormones regulate submergence-induced shoot elongation in the flooding-tolerant dicot Rumex palustris. Ann. Bot. 2003, 91, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Kurzatkowski, D.; Leuschner, C.; Homeier, J. Effects of flooding on trees in the semi-deciduous transition forests of the Araguaia floodplain, Brazil. Acta Oecol. 2015, 69, 21–30. [Google Scholar] [CrossRef]
- Worbes, M.; Klinge, H.; Revilla, J.D.; Martius, C. On the dynamics, floristic subdivision and geographical distribution of varzea forests in Central Amazonia. J. Veg. Sci. 1992, 3, 553–564. [Google Scholar] [CrossRef]
- Resende, A.F.; Piedade, M.T.F.; Feitosa, Y.O.; Andrade, V.H.F.; Trumbore, S.E.; Durgante, F.M.; Macedo, M.O.; Schöngart, J. Flood-pulse disturbances as a threat for long-living Amazonian trees. New Phytol. 2020, 227, 1790–1803. [Google Scholar] [CrossRef] [PubMed]
- Marinho, T.A.S.; Piedade, M.T.F.; Wittmann, F. Distribution and population structure of four Central Amazonian high-várzea timber species. Wetl. Ecol. Manag. 2010, 18, 665–677. [Google Scholar] [CrossRef]
- Schöngart, J.; Piedade, M.T.F.; Wittmann, F.; Junk, W.J.; Worbes, M. Wood growth patterns of Macrolobium acaciifolium (Benth.) Benth. (Fabaceae) in Amazonian black-water and white-water floodplain forests. Oecologia 2005, 145, 454–461. [Google Scholar] [CrossRef]
- Parolin, P. Phenology and CO2-assimilation of trees in Central Amazonian floodplains. J. Trop. Ecol. 2000, 16, 465–473. [Google Scholar] [CrossRef]
- Oliveira-Wittmann, A.; Piedade, M.T.F.; Wittmann, F.; Parolin, P. Germination in four low-várzea tree species of Central Amazonia. Aquat. Bot. 2007, 86, 197–203. [Google Scholar] [CrossRef]
- Kubitzki, K.; Ziburski, A. Seed dispersal of floodplain forest in Amazonia. Biotropica 1994, 26, 30–43. [Google Scholar] [CrossRef]
- Wittmann, F.; Parolin, P. Phenology of six tree species from Central Amazonian várzea. Ecotropica 1999, 5, 51–57. [Google Scholar]
- Schöngart, J.; Piedade, M.T.F.; Ludwigshausen, S.; Horna, V.; Worbes, M. Phenology and stem-growth periodicity of tree species in Amazonian floodplain forests. J. Trop. Ecol. 2002, 18, 581–597. [Google Scholar] [CrossRef] [Green Version]
- Piedade, M.T.F.; Parolin, P.; Junk, W.J. Phenology, fruit production and seed dispersal of Astrocaryum jauari (Arecaceae) in Amazonian black water floodplains. Rev. Biol. Trop. 2006, 54, 1171–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrade, A.C.S.; Ramos, F.N.; Souza, A.F.; Loureiro, M.B.; Bastos, R. Flooding effects in seedlings of Cytharexyllum myrianthum Cham. and Genipa americana L.: Responses of two neotropical lowland tree species. Rev. Bras. Bot. 1999, 22, 281–285. [Google Scholar] [CrossRef] [Green Version]
- Kubitzki, K. The dispersal of forest plants. In Amazonia–Key Environments; Prance, G.T., Lovejoy, T.E., Eds.; Pergamon: Oxford, UK, 1985; pp. 129–163. [Google Scholar]
- Waldhoff, D.; Saint-Paul, U.; Furch, B. Value of fruits and seeds from the floodplain forests of Central Amazonia as food resource for fish. Ecotropica 1996, 2, 143–156. [Google Scholar]
- Ziburski, A. Dissemination, Keimung und Etablierung einiger Baumarten der Über-schwemmungswälder Amazoniens. In Tropische und Subtropische Pflanzenwelt; Rauh, W., Ed.; Akademie der Wissenschaften und der Literatur: Mainz, Germany, 1991; Volume 77, pp. 1–96. [Google Scholar]
- Gottsberger, G. Seed dispersal by fish in the inundated region of Humaitá, Amazonia. Biotropica 1978, 10, 170–183. [Google Scholar] [CrossRef]
- Weiss, B.; Zuanon, J.A.S.; Piedade, M.T.F. Viability of seeds consumed by fishes in a lowland forest in the Brazilian Central Amazon. Trop. Conserv. Sci. 2016, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Correa, S.B.; Winemiller, K.O.; López-Fernández, H.; Galeti, M. Evolutionary perspectives on seed consumption and dispersal by fishes. BioScience 2007, 57, 748–756. [Google Scholar] [CrossRef]
- Mannheimer, S.; Bevilacqua, G.; Caramaschi, E.P.; Scarano, F.R. Evidence for seed dispersal by the catfish Auchenipterrichthys longimanus in an Amazonian lake. J. Trop. Ecol. 2003, 19, 215–218. [Google Scholar] [CrossRef]
- Ferreira, C.S.; Piedade, M.T.F.; Franco, A.; Gonçalves, J.F.C.; Junk, W.J. Adaptive strategies to tolerate prolonged flooding in seedlings of floodplain and upland populations of Himatanthus sucuuba, a Central Amazon tree. Aquat. Bot. 2009, 90, 246–252. [Google Scholar] [CrossRef]
- Maurenza, D.; Marenco, R.A.; Parolin, P.; Piedade, M.T.F. Physiological responses to flooding and light in two tree species native to the Amazonian floodplains. Aquat. Bot. 2012, 96, 7–13. [Google Scholar] [CrossRef]
- Fontes, C.G.; Fine, P.V.A.; Wittmann, F.; Bittencourt, P.R.L.; Piedade, M.T.F.; Higuchi, N.; Chambers, J.Q.; Dawson, T.E. Convergent evolution of tree hydraulic traits in Amazonian habitats: Implications for community assemblage and vulnerability to drought. New Phytol. 2020, 228, 106–120. [Google Scholar] [CrossRef]
- Sperry, J.S.; Hacke, U.G.; Pittermann, J. Size and function in conifer tracheids and angiosperm vessels. Am. J. Bot. 2006, 93, 1490–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gleason, S.M.; Westoby, M.; Jansen, S.; Choat, B.; Hacke, U.G.; Pratt, R.B.; Bhaskar, R.; Brodripp, T.J.; Bucci, S.J.; Cao, K.-F.; et al. Weak tradeoff between xylem safety and xylem-specific hydraulic efficiency across the world’s woody plant species. New Phytol. 2016, 209, 123–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hacke, U.G.; Spicer, R.; Schreiber, S.G.; Plavcova, L. An ecophysiological and developmental perspective on variation in vessel diameter. Plant Cell Environ. 2017, 40, 831–845. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S. From tropics to tundra: Global convergence in plant functioning. Proc. Natl. Acad. Sci. USA 1997, 94, 13730–13734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westoby, M. A leaf-height-seed (LHS) plant ecology strategy scheme. Plant Soil 1998, 199, 213–227. [Google Scholar] [CrossRef]
- Fortunel, C.; Ruelle, J.; Beauchene, J.; Fine, P.V.A.; Baraloto, C. Wood specific gravity and anatomy of branches and roots in 113 Amazonian rainforest tree species across environmental gradients. New Phytol. 2013, 202, 79–94. [Google Scholar] [CrossRef]
- Kunstler, G.; Falster, D.; Coomes, D.A.; Hui, F.; Kooyman, R.M.; Laughlin, D.C.; Poorter, L.; Vanderwel, M.; Vieilledent, G.; Wright, S.J.; et al. Plant functional traits have globally consistent effects on competition. Nature 2016, 529, 204–207. [Google Scholar] [CrossRef] [Green Version]
- Mori, S. A familia da Castanha-do-Pará: Símbolo do Rio Negro. In Florestas do Rio Negro; Oliveira, A.A., Daly, D.C., Eds.; UNIP, NYBG e companhia das Letras: São Paulo, Brazil, 2001; pp. 119–142. [Google Scholar]
- Irion, G.; Müller, J.; De Mello, J.; Junk, W.J. Quaternary geology of the Amazon lowland. Geo-Mar. Lett. 1995, 15, 172–177. [Google Scholar] [CrossRef]
- Piedade, M.T.F.; Junk, W.J.; Long, S.P. The productivity of the C4 grass Echinochloa polystachia on the Amazon Floodplain. Ecology 1991, 72, 1456–1463. [Google Scholar] [CrossRef]
- Fonseca Júnior, S.F.; Piedade, M.T.F.; Schöngart, J. Wood growth of Tabebuia barbata (E. Mey.) Sandwith (Bignoniaceae) and Vatairea guianensis Aubl. (Fabaceae) in Central Amazonian black-water (igapó) and white-water (várzea) floodplain forests. Trees 2009, 23, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Malhi, Y.; Baker, T.R.; Phillips, O.L.; Almeida, S.; Alvarez, E.; Arroyo, L.; Chave, J.; Czimczik, C.I.; Di Fiore, A.; Higuchi, N.; et al. The above-ground coarse wood productivity of 104 neotropical forest plots. Glob. Change Biol. 2004, 10, 563–591. [Google Scholar] [CrossRef] [Green Version]
- Mori, G.B.; Schietti, J.; Poorter, L.; Piedade, M.T.F. Trait divergence and habitat specialization in tropical floodplain forest trees. PLoS ONE 2019, 14, e0212232. [Google Scholar] [CrossRef] [PubMed]
- Prance, G.T. Estudos sobre a vegetação das campinas Amazônicas. I: Introdução a uma série de publicações sobre a vegetação das campinas Amazônicas. Acta Amaz. 1975, 5, 207–209. [Google Scholar] [CrossRef] [Green Version]
- Medina, E.; Sobrado, M.; Herrera, R. Significance of leaf orientation for leaf temperature in an Amazonian sclerophyll vegetation. Radiat. Environ. Biophys. 1978, 15, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Coomes, D.A. Nutrient status of Amazonian caatinga forests in a seasonally dry area: Nutrient fluxes in litter fall and analyses of soils. Can. J. For. Res. 1997, 27, 831–839. [Google Scholar] [CrossRef]
- Medina, E.; García, V.; Cuevas, E. Sclerophylly and oligotrophic environments: Relationships between leaf structure, mineral nutrient content, and drought resistance in tropical rainforest of the upper Negro region. Biotropica 1990, 22, 51–64. [Google Scholar] [CrossRef]
- Parolin, P. Seed germination and early establishment of 12 tree species from nutrient-rich and nutrient-poor Central Amazonian floodplains. Aquat. Bot. 2001, 70, 89–103. [Google Scholar] [CrossRef]
- Fine, P.V.A.; Mesones, I.; Coley, P.D. Herbivores promote habitat specialization by trees in Amazonian forests. Science 2004, 305, 663–665. [Google Scholar] [CrossRef] [Green Version]
- Furch, K. Chemistry of várzea and igapó soils and nutrient inventory of their floodplain forests. In The Central Amazon Floodplain: Ecology of a Pulsing System. Ecological Studies 126; Junk, W.J., Ed.; Springer: Berlin/Heidelberg, Germany, 1997; pp. 47–68. [Google Scholar]
- Klinge, H.; Medina, E. Rio Negro caatingas and campinas, Amazonas States of Venezuela and Brazil. In Heatlands and Related Shrublands. Ecosystems of the World; Specht, R.L., Ed.; Elsevier: Amsterdam, The Netherlands, 1979; Volume 9, pp. 483–488. [Google Scholar]
- Santos, A.R.; Nelson, B.W. Leaf decomposition and fine fuels in floodplain forests of the Rio Negro in the Brazilian Amazon. J. Trop. Ecol. 2013, 29, 455–458. [Google Scholar] [CrossRef]
- Meyer, U.; Junk, W.J.; Linck, C. Fine root systems and mycorrhizal associations in two central Amazonian inundation forests: Igapó and várzea. In Amazonian Floodplain Forest: Ecophysiology, Biodiversity and Sustainable Management. Ecological Studies 210; Junk, W.J., Piedade, M.T.F., Wittmann, F., Eds.; Springer: Berlin, Germany, 2010; pp. 163–178. [Google Scholar]
- Parolin, P.; Wittmann, F.; Ferreira, L.V. Fruit and seed dispersal in Amazonian floodplain trees—A review. Ecotropica 2013, 19, 15–32. [Google Scholar]
- Kalliola, R.; Salo, J.; Puhakka, M.; Rajasilta, M. New site formation and colonizing vegetation in primary succession on the western Amazon floodplains. J. Ecol. 1991, 79, 877–901. [Google Scholar] [CrossRef]
- Keel, S.H.K.; Prance, G.T. Studies of the vegetation of a white-sand blackwater igapó (Rio Negro, Brazil). Acta Amaz. 1979, 9, 645–655. [Google Scholar] [CrossRef]
- Almeida, D.R.A.; Nelson, B.W.; Schietti, J.; Gorgens, E.B.; Resende, A.F.; Stark, S.C.; Valbuena, R. Contrasting fire damage and fire susceptibility between seasonally flooded forest and upland forest in the Central Amazon using portable profiling LiDAR. Remote Sens. Environ. 2016, 184, 153–160. [Google Scholar] [CrossRef]
- Godoy, J.R.; Petts, G.; Salo, J. Riparian flooded forests of the Orinoco and Amazon basins: A comparative review. Biodivers. Conserv. 1999, 8, 551–586. [Google Scholar] [CrossRef]
- Campbell, D.G.; Stone, J.L.; Rosas, A. A comparison of the phytosociology and dynamics of three floodplain (várzea) forests of known ages, Rio Juruá, western Brazilian Amazon. Bot. J. Linn. Soc. 1992, 108, 213–237. [Google Scholar] [CrossRef]
- Mertes, L.A.K.; Daniel, D.L.; Melack, J.M.; Nelson, B.; Martinelli, L.A.; Forsberg, B.R. Spatial patterns of hydrology, geomorphology, and vegetation on the floodplain of the Amazon River in Brazil from a remote sensing perspective. Geomorphology 1995, 13, 215–232. [Google Scholar] [CrossRef]
- Peixoto, J.M.A.; Nelson, B.W.; Wittmann, F. Spatial and temporal dynamics of alluvial geomorphology and vegetation in central Amazonian white-water floodplains by remote-sensing techniques. Remote Sens. Environ. 2009, 113, 2258–2266. [Google Scholar] [CrossRef]
- Kalliola, R.; Salo, J.; Puhakka, M.; Rajasilta, M.; Häme, T.; Neller, R.J.; Räsänen, M.E.; Arias, W.A.D. Upper Amazon channel migration: Implications for vegetation disturbance and succession using bitemporal Landsat MSS images. Naturwissenschaften 1992, 79, 75–79. [Google Scholar] [CrossRef]
- Latrubesse, W.M.; Franzinelli, E. The late Quaternary evolution of the Negro River, Amazon, Brazil: Implications for island and floodplain formation in large anabranching tropical systems. Geomorphology 2005, 70, 372–397. [Google Scholar] [CrossRef]
- Irion, G. Sedimentation and sediments of Amazonian rivers and evolution of the Amazonian landscape since Pliocene times. In The Amazon: Limnology and Landscape Ecology of a Mighty Tropical River and Its Basin; Sioli, H., Ed.; Dr. W. Junk Publications: Dordrecht, The Netherlands, 1984; pp. 201–214. [Google Scholar]
- Swaine, M.D.; Whitmore, T.C. On the definition of ecological species groups in tropical rain forests. Vegetatio 1988, 75, 81–86. [Google Scholar] [CrossRef]
- Wittmann, F.; Junk, W.J. Sapling communities in Amazonian white-water forests. J. Biogeogr. 2003, 30, 1577–1588. [Google Scholar] [CrossRef]
- Wittmann, F.; Schöngart, J.; Piedade, M.T.F.; Junk, W.J. Tropical large-river wetlands. In Encyclopedia of Inland Waters, 2nd ed.; Mehner, T., Tockner, K., Eds.; Elsevier: Oxford, UK, 2022; Volume 3, pp. 90–104. [Google Scholar]
- Chambers, J.Q.; Robertson, A.L.; Carneiro, V.M.C.; Lima, A.J.N.; Smith, M.-L.; Plourde, L.C.; Higuchi, N. Hyperspectral remote detection of niche partitioning among canopy trees driven by blowdown gap disturbances in the Central Amazon. Oecologia 2009, 160, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Veloso, H.P.; Rangel Filho, A.R.L.; Lima, J.C.A. Classificação da Vegetação Brasileira, Adaptada a um Sistema Universal; Fundação Instituto Brasileiro de Geografia e Estatística: Rio de Janeiro, Brazil, 1991. [Google Scholar]
- Householder, J.E.; Wittmann, F.; Tobler, M.W.; Janovec, J.P. Montane bias in lowland Amazonian peatlands: Plant assembly on heterogeneous landscapes and potential significance to palynological inference. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 423, 138–148. [Google Scholar] [CrossRef]
- Ferreira, C.S.; Piedade, M.T.F.; Oliveira Wittmann, A.; Franco, A. C Plant reproduction in the Central Amazonian floodplains: Challenges and adaptations. AOB Plants 2010, 2010, plq009. [Google Scholar] [CrossRef] [Green Version]
- Puhakka, M.; Kalliola, R. La vegetación en áreas de inundación en la selva baja de la Amazonia Peruana. In Amazonia Peruana: Vegetación Húmeda Tropical em el Llano Subandino; Kalliola, R., Puhakka, M., Danjoy, W., Eds.; Proyecto Amazonia: Turku, Finland, 1993; pp. 113–138. [Google Scholar]
- Connell, J.H.; Slatyer, R.O. Mechanisms of succession in natural communities in their role in community stability and organization. Am. Nat. 1977, 111, 1119–1144. [Google Scholar] [CrossRef]
- Salo, J.; Kalliola, R.; Häkkinen, L.; Mäkinen, Y.; Niemelä, P.; Puhakka, M.; Coley, P.D. River dynamics and the diversity of the Amazon lowland forest. Nature 1986, 322, 254–258. [Google Scholar] [CrossRef]
- Latrubesse, E.M. Patterns of anabranching channels: The ultimate end-member adjustment of mega rivers. Geomorphology 2008, 101, 130–145. [Google Scholar] [CrossRef]
- Sombroek, W. Spatial and temporal patterns of Amazonian rainfall: Consequences for the planning of agricultural occupation and the protection of primary forests. Ambio 2001, 30, 388–396. [Google Scholar] [CrossRef]
- Schöngart, J.; Junk, W.J.; Piedade, M.T.F.; Ayres, J.M.; Hüttermann, A.; Worbes, M. Teleconnection between tree growth in the Amazonian floodplains and the El Niño-Southern Oscillation Effect. Glob. Change Biol. 2004, 10, 683–692. [Google Scholar] [CrossRef]
- Schöngart, J.; Junk, W.J. Forecasting the flood pulse in Central Amazonia by ENSO-indices. J. Hydrol. 2007, 335, 124–132. [Google Scholar] [CrossRef]
- Stroh, C.L.; De Steven, D.; Guntenspergen, G.R. Effect of climate fluctuations on long-term vegetation dynamics in Carolina Bay wetlands. Wetlands 2008, 28, 17–27. [Google Scholar] [CrossRef]
- Medina, E. Adaptations of tropical trees to moisture stress. In Ecosystems of the World: Tropical Rain Forest Ecosystems; Golley, F.B., Ed.; Elsevier Scientific Publishing Co.: Amsterdam, The Netherlands, 1983; pp. 225–237. [Google Scholar]
- Waldhoff, D.; Furch, B. Leaf morphology and anatomy in eleven tree species from Central Amazonian floodplains (Brazil). Amazoniana 2002, 17, 79–94. [Google Scholar]
- Waldhoff, D.; Junk, W.J.; Furch, B. Responses of three Amazonian tree species to drought and flooding under controlled conditions. Int. J. Ecol. Environ. Sci. 1998, 24, 237–252. [Google Scholar]
- Piedade, M.T.F.; Junk, W.J.; Parolin, P. The flood pulse and photosynthetic response of trees in a white water floodplain (várzea) of the Central Amazon, Brazil. Proc. Int. Assoc. Theor. Appl. Limnol. 2000, 27, 1734–1739. [Google Scholar]
- Armbrüster, N.; Müller, E.; Parolin, P. Contrasting responses of two Amazonian floodplain trees to hydrological changes. Ecotropica 2004, 10, 73–84. [Google Scholar]
- Schlüter, U.-B. Morphologische, Anatomische und Physiologische Untersuchungen zur Überflutungstoleranz Zweier Charakteristischer Baumarten des Weiß- und Schwarzwasser Überschwemmungswaldes bei Manaus. Ein Beitrag zur Ökosystemanalyse von Várzea und Igapó Zentralamazoniens. Ph.D. Thesis, Universität Kiel, Kiel, Germany, 1989. [Google Scholar]
- Worbes, M. Lebensbedingungen und Holzwachstum in zentralamazonischen Überschwemmungswäldern. Scr. Geobot. 1986, 17, 1–112. [Google Scholar]
- Resende, A.F.; Nelson, B.W.; Flores, B.M.; Almeida, D.R.A. Fire damage in seasonally flooded and upland forests of the Central Amazon. Biotropica 2014, 46, 643–646. [Google Scholar] [CrossRef]
- Waldhoff, D.; Parolin, P. Morphology and anatomy of leafs. In Central Amazonian Floodplain Forests: Ecophysiology, Biodiversity and Sustainable Management. Ecological Studies 210; Junk, W.J., Piedade, M.T.F., Wittmann, F., Eds.; Springer: Berlin, Germany, 2010; pp. 179–202. [Google Scholar]
- Davidson, E.A.; Araújo, A.C.; Artaxo, P.; Balch, J.K.; Brown, I.F.; Bustamante, M.M.C.; Coe, M.T.; DeFries, R.S.; Keller, M.; Longo, M.; et al. The Amazon basin in transition. Nature 2012, 481, 321–328. [Google Scholar] [CrossRef]
- Aragão, L.E.O.C.; Anderson, L.O.; Fonseca, M.G.; Rosan, T.M.; Vedovato, L.B.; Wagner, F.H.; Silva, C.V.J.; Silva Junior, C.H.L.; Arai, E.; Aguiar, A.P.; et al. 21st century drought-related fires counteract the decline of Amazon deforestation carbon emissions. Nat. Commun. 2018, 9, 536. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, T.C.; Wittmann, F.; Piedade, M.T.F.; Resende, A.F.; Silva, T.S.F.; Schöngart, J. Fires in Amazonian blackwater floodplain forests: Causes, human dimension, and implications for conservation. Front. For. Glob. Chang. 2021, 4, 755441. [Google Scholar] [CrossRef]
- Flores, B.M.; Fagoaga, R.; Nelson, B.W.; Holmgren, M. Repeated fires trap Amazonian blackwater floodplains in an open vegetation state. J. Appl. Ecol. 2016, 53, 1597–1603. [Google Scholar] [CrossRef]
- Flores, B.M.; Holmgren, M. White-sand savannas expand at the core of the Amazon after forest wildfires. Ecosystems 2021, 24, 1624–1637. [Google Scholar] [CrossRef]
- Marengo, J.A.; Souza, C.A.J.; Thonicke, K.; Burton, C.; Halladay, K.; Betts, R.A.; Alves, L.M.; Soares, W.R. Changes in climate and land use over the Amazon region: Current and future variability and trends. Front. Earth Sci. 2018, 6, 228. [Google Scholar] [CrossRef]
- Brunke, M.; Gonser, T. The ecological significance of exchange processes between rivers and groundwater. Freshw. Biol. 1997, 37, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Naiman, R.J.; Décamps, H.; Pollock, M. The role of riparian corridors in maintaining regional biodiversity. Ecol. Appl. 1993, 3, 209–212. [Google Scholar] [CrossRef]
- Salati, E.; Vose, P.B. Amazon basin: A system in equilibrium. Science 1984, 225, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Togersen, C.E.; Faux, R.N.; McIntosh, B.A.; Poage, N.J.; Norton, D.J. Airborne thermal remote sensing for water temperature assessment in rivers and streams. Remote Sens. Environ. 2001, 76, 386–398. [Google Scholar] [CrossRef]
- Tonolla, D.; Acuña, V.; Uehlinger, U.; Frank, T.; Tockner, K. Thermal heterogeneity in river floodplains. Ecosystems 2010, 13, 727–740. [Google Scholar] [CrossRef] [Green Version]
- Oren, R.; Zimmermann, R.; Terborgh, J. Transpiration in upper Amazonian floodplain and upland forests in response to drought-braking rains. Ecology 1996, 77, 968–973. [Google Scholar] [CrossRef]
- Cermák, J.; Prax, A. Transpiration and soil water supply in floodplain forests. Ekológia 2009, 28, 248–254. [Google Scholar] [CrossRef]
- Ribeiro, M.N.G.; Adis, J. Local rainfall variability: A potential bias for bioecological studies in the Central Amazon. Acta Amaz. 1984, 14, 159–174. [Google Scholar] [CrossRef] [Green Version]
- Meave, J.; Kellman, M.; Mac Dougall, D.; Rosales, J. Riparian habitats as tropical forest refugia. Glob. Ecol. Biogeogr. Lett. 1991, 1, 69–76. [Google Scholar] [CrossRef]
- Oliveira-Filho, A.T.; Ratter, J.A. A study of the origin of Central Brazilian forests by the analysis of plant species distribution patterns. Edinb. J. Bot. 1995, 52, 141–194. [Google Scholar] [CrossRef]
- Irion, G.; Kalliola, R. Long-term landscape development processes in Amazonia. In Amazonia, Landscape and Species Evolution: A Look into the Past; Hoorn, C., Wesselingh, F.P., Eds.; Blackwell Publications: Oxford, UK, 2010; pp. 185–197. [Google Scholar]
- Goldberg, S.L.; Schmidt, M.J.; Perron, J.T. Fast response of Amazon rivers to Quaternary climate cycles. J. Geophys. Res. Earth Surf. 2021, 126, e2021JF006416. [Google Scholar] [CrossRef]
- Junk, W.J.; Cunha, C.N.; Wantzen, K.M.; Petermann, P.; Strüssmann, C.; Marques, M.I.; Adis, J. Biodiversity and its conservation in the Pantanal of Mato Grosso, Brazil. Aquat. Sci. 2006, 68, 279–309. [Google Scholar] [CrossRef]
- Montes, R.; San José, J.J. Vegetation and soil analyses of topo-sequences in the Orinoco llanos. Flora 1995, 190, 1–33. [Google Scholar] [CrossRef]
- Veneklaas, E.J.; Fajardo, A.; Obregon, S.; Lozano, J. Gallery forest types and their environmental correlates in a Colombian Savanna landscape. Ecography 2005, 28, 236–252. [Google Scholar] [CrossRef]
- Wittmann, F.; Marques, M.C.M.; Damasceno Júnior, C.; Budke, J.C.; Piedade, M.T.F.; Oliveira Wittmann, A.; Montero, J.C.; de Assis, R.L.; Targhetta, N.; Parolin, P.; et al. The Brazilian freshwater wetscape: Changes in tree community diversity and composition on climatic and geographic gradients. PLoS ONE 2017, 12, e0175003. [Google Scholar] [CrossRef] [Green Version]
- Gentry, A.H. Changes in plant community diversity and floristic composition on environmental and geographical gradients. Ann. Mo. Bot. Gard. 1988, 75, 1–34. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, C.S.; Piedade, M.T.F.; Junk, W.J.; Parolin, P. Floodplain and upland populations of Amazonian Himatanthus sucuuba: Effects of flooding on germination, seedling growth, and mortality. Environ. Exp. Bot. 2007, 60, 477–483. [Google Scholar] [CrossRef]
- Ferreira, C.S.; Figueira, A.V.O.; Gribel, R.; Wittmann, F.; Piedade, M.T.F. Genetic variability, divergence, and speciation in trees of periodically flooded forests of the Amazon: A case study of Himatanthus sucuuba (Spruce) Woodson. In Amazonian Floodplain forests: Ecophysiology, Biodiversity and Sustainable Management. Ecological Studies 210; Junk, W.J., Piedade, M.T.F., Wittmann, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 301–312. [Google Scholar]
- Donoghue, M.J. A phylogenetic perspective in the distribution of plant diversity. Proc. Natl. Acad. Sci. USA 2008, 105, 11549–11555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prance, G.T.; Schubart, F.O.R. Nota preliminar sobre a origem das campinas abertas de areia branca do baixo Rio Negro. Acta Amaz. 1978, 7, 567–569. [Google Scholar] [CrossRef]
- Jordan, C.F. Soils of the Amazon rainforest. In Key Environments: Amazonia; Prance, G.T., Lovejoy, T.E., Eds.; Pergamon Press: Oxford, UK, 1985; pp. 83–105. [Google Scholar]
- Steyermark, J.A. Speciation and endemism in the flora of the Venezuelan tepuis. In High Altitude Tropical Biogeography; Vuilleumier, F., Monasterio, M., Eds.; Oxford University Press: Oxford, UK, 1986; pp. 317–373. [Google Scholar]
- Huber, O. Guyana highlands versus Guyana lowlands, a reappraisal. Taxon 1984, 37, 595–614. [Google Scholar] [CrossRef]
- Prance, G.T. Islands in Amazonia. Philos. Trans. R. Soc. B Biol. Sci. 1996, 351, 823–833. [Google Scholar]
- Vicentini, A. A vegetacão ao longo de um gradient edáfico no Parque Nacional do Jaú. In Janelas para a Biodiversidade no Parque Nacional do Jaú: Uma Estratégia para o Estudo da Biodiversidade na Amazônia; Fundacão Vitória, Amazonica; Borges, S.H., Iwanaga, S., Durigan, C.C., Pinheiro, R.R., Eds.; WWF, IBAMA: Manaus, Brazil, 2004; pp. 117–143. [Google Scholar]
- Fine, P.V.A.; García-Villacorta, R.; Pitman, N.C.A.; Mesones, I.; Kembel, S.W. A floristic study of the white-sand forests of Peru. Ann. Mo. Bot. Gard. 2010, 97, 283–305. [Google Scholar] [CrossRef]
- Damasco, G.; Vicentini, A.; Castilho, C.V.; Pimentel, T.P.; Nascimento, H.E.M. Disentangling the role of edaphic variability, flooding regime and topography of Amazonian white-sand vegetation. J. Veg. Sci. 2013, 24, 384–394. [Google Scholar] [CrossRef] [Green Version]
- Guevara, J.E.; Damasco, G.; Baraloto, C.; Fine, P.V.A.; Peñuela, M.C.; Castilho, C.; Vicentini, A.; Cárdenas, D.; Wittmann, F.; Targhetta, N.; et al. Low phylogenetic beta diversity and geographic neo-endemism in Amazonian white sand forests. Biotropica 2016, 48, 34–46. [Google Scholar] [CrossRef]
- Demarchi, L.O.; Klein, V.P.; Aguiar, D.P.P.; Marinho, L.C.; Ferreira, M.J.; Lopes, A.; Da Cruz, J.; Quaresma, A.C.; Schöngart, J.; Wittmann, F.; et al. The specialized white-sand flora of the Uatumã Sustainable Development Reserve, central Amazon, Brazil. Checklist 2022, 18, 187–217. [Google Scholar] [CrossRef]
- Bongers, F.; Engelen, D.; Klinge, H. Phytomass structure of natural plant communities on spodosols in southern Venezuela: The Bana woodland. Vegetatio 1985, 63, 13–34. [Google Scholar] [CrossRef]
- García-Villacorta, R.; Dexter, K.G.; Pennington, T. Amazonian white-sand forests show strong floristic links with surrounding oligotrophic habitats and the Guiana Shield. Biotropica 2016, 48, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Gentry, A.H. Neotropical floristic diversity: Phytogeographical connections between Central and South America, Pleistocene climatic fluctuations, or an accident of the Andean orogeny? Ann. Mo. Bot. Gard. 1982, 69, 557–593. [Google Scholar] [CrossRef]
- Baker, T.R.; Phillips, O.L.; Malhi, Y.; Almeida, S.; Arroyo, L.; Di Fiore, A.; Killeen, T.J.; Laurance, S.G.; Laurance, W.F.; Lewis, S.L.; et al. Variation in wood density determines spatial patterns in Amazonian forest biomass. Glob. Change Biol. 2004, 10, 545–562. [Google Scholar] [CrossRef]
- Saatchi, S.S.; Houghton, R.A.; Dos Santos Alvalá, R.C.; Soares, J.V.; Yu, Y. Distribution of aboveground wood biomass in the Amazon basin. Glob. Change Biol. 2007, 13, 816–837. [Google Scholar] [CrossRef]
- Hoorn, C. Marine incursions and the influence of Andean tectonics on the Miocene depositional history of northwestern Amazonia: Results of a palynostratigraphic study. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1993, 105, 267–309. [Google Scholar] [CrossRef]
- Fine, P.V.A.; Daly, D.C.; Cameron, C.N. The contribution of edaphic heterogeneity to the evolution and diversity of Burseraceae trees in the Western Amazon. Evolution 2005, 59, 1464–1478. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, A.; Nylander, A.A.; Persson, C.; Sanmartín, L. Tracing the impact of the Andean uplift on Neotropical plant evolution. Proc. Natl. Acad. Sci. USA 2009, 106, 9749–9754. [Google Scholar] [CrossRef] [Green Version]
- Antonelli, A.; Sanmartíın, I. Why are there so many plant species in the Neotropics? Taxon 2011, 60, 403–414. [Google Scholar] [CrossRef]
- Roncal, J.; Khan, F.; Millan, B.; Couvreur, T.L.P.; Pintaud, J.-C. Cenozoic colonization and diversification patterns of tropical American palms: Evidence from Astrocaryum (Arecaceae). Bot. J. Linn. Soc. 2013, 171, 120–139. [Google Scholar] [CrossRef] [Green Version]
- Ter Steege, H.; Pitman, N.; Sabatier, D.; Castellanos, H.; Van der Hout, P.; Daly, D.C.; Silveira, M.; Phillips, O.; Vasquez, R.; Van Andel, T.; et al. A spatial model of tree alpha-diversity and beta-density for the Amazon Region. Biodivers. Conserv. 2003, 12, 2255–2277. [Google Scholar] [CrossRef]
- Baker, T.R.; Pennington, R.T.; Magallon, S.; Gloor, E.; Laurance, W.F.; Alexiades, M.; Alvarez, E.; Araujo, A.; Arets, E.J.M.M.; Aymard, G.; et al. Fast demographic traits promote high diversification rates of Amazonian trees. Ecol. Lett. 2014, 17, 527–536. [Google Scholar] [CrossRef] [Green Version]
- Kellman, M.; Meave, J. Fire in the tropical gallery forests of Belize. J. Biogeogr. 1997, 24, 23–34. [Google Scholar] [CrossRef]
- McLaughlin, D.L.; Cohen, M.J. Realizing ecosystem services: Wetland hydrologic function along a gradient of ecosystem condition. Ecol. Appl. 2013, 23, 1619–1631. [Google Scholar] [CrossRef]
- Wallace, A.R. On the monkeys of the Amazon. Proc. Zool. Soc. Lond. 1852, 20, 107–110. [Google Scholar] [CrossRef]
- Boubli, J.P.; Ribas, C.; Alfaro, J.W.L.; Alfaro, M.E.; Da Silva, M.N.F.; Pinho, G.M.; Farias, I.P. Spatial and temporal patterns of diversification on the Amazon: A test of the riverine hypothesis for all diurnal primates of Rio Negro and Rio Branco in Brazil. Mol. Phylogenet. Evol. 2015, 82, 400–412. [Google Scholar] [CrossRef] [PubMed]
- Haffer, J.; Prance, G.T. Climatic forcing of evolution in Amazonia during the Cenozoic: On the refuge theory of biotic differentiation. Amazoniana 2001, 16, 579–607. [Google Scholar]
- Ribas, C.C.; Aleixo, A.; Nogueira, A.C.R.; Miyaki, C.Y.; Cracraft, J. A paleobiogeographic model for biotic diversification within Amazonia over the past three million years. Proc. R. Soc. B Biol. Sci. 2012, 279, 681–689. [Google Scholar] [CrossRef]
- Ávila Pires, T.C.S. Lizards of Brazilian Amazonia. Zool. Verh. Leiden 1995, 299, 1–706. [Google Scholar]
- Hall, J.P.W.; Harvey, D. The phylogeography of Amazonia revisited: New evidence from riodinid butterflies. Evolution 2002, 56, 1489–1497. [Google Scholar] [CrossRef]
- Dambros, C.; Zuquim, G.; Moulatlet, G.M.; Costa, F.R.C.; Tuomisto, H.; Ribas, C.C.; Azevedo, R.; Baccaro, F.; Bobrowiec, P.E.D.; Dias, M.S.; et al. The role of environmental filtering, geographic distance and dispersal barriers in shaping the turnover of plant and animal species in Amazonia. Biodivers. Conserv. 2020, 29, 3609–3634. [Google Scholar] [CrossRef]
- Silva, J.M.C.; Rylands, A.B.; Fonseca, G.A.B. The fate of the Amazon areas of endemism. Conserv. Biol. 2005, 19, 689–694. [Google Scholar] [CrossRef]
- Nepstad, D.; McGrath, D.; Stickler, C.; Alencar, A.; Azevedo, A.; Swette, B.; Bezerra, T.; Digiano, M.; Shimada, J.; Da Motta, R.S.; et al. Slowing Amazon deforestation through public policy and interventions in beef and soy supply chains. Science 2014, 344, 1118–1123. [Google Scholar] [CrossRef] [PubMed]
- Castello, L.; McGrath, D.G.; Hess, L.L.; Coe, M.T.; Lefebvre, P.A.; Petry, P.; Macedo, M.N.; Renó, V.F.; Arantes, C.C. The vulnerability of Amazon freshwater ecosystems. Conserv. Lett. 2013, 6, 217–229. [Google Scholar] [CrossRef]
- McGrath, D.G.; Cardoso, A.; Almeida, O.T.; Pezzuti, J. Constructing a policy and institutional framework for an ecosystem-based approach to managing the Lower Amazon floodplain. Environ. Dev. Sustain. 2008, 10, 677–695. [Google Scholar] [CrossRef]
- Renó, V.F.; Novo, E.M.L.M.; Suemitsu, C.; Rennó, C.D.; Silva, T.S.F. Assessment of deforestation in the Lower Amazon floodplain using historical Landsat MSS/TM imagery. Remote Sens. Environ. 2011, 115, 3446–3456. [Google Scholar] [CrossRef]
- Junk, W.J.; Piedade, M.T.F.; Wittmann, F.; Schöngart, J. The role of floodplain forests in an integrated sustainable management concept of the natural resources of the Central Amazonian várzea. In Amazonian Floodplain Forests: Ecophysiology, Biodiversity and Sustainable Management. Ecological Studies 210; Junk, W.J., Piedade, M.T.F., Wittmann, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 485–510. [Google Scholar]
- Timpe, K.; Kaplan, D. The changing hydrology of a dammed Amazon. Sci. Adv. 2017, 3, e1700611. [Google Scholar] [CrossRef]
- Assahira, C.; Piedade, M.T.F.; Trumbore, S.E.; Wittmann, F.; Cintra, B.B.L.; Batista, E.S.; Resende, A.F.; Schöngart, J. Tree mortality of a flood-adapted species in response of hydrographic changes caused by an Amazonian river dam. For. Ecol. Manag. 2017, 396, 113–123. [Google Scholar] [CrossRef]
- Resende, A.F.; Schöngart, J.; Streher, A.S.; Ferreira-Ferreira, J.; Piedade, M.T.F.; Silva, T.S.F. Massive tree mortality from flood-pulse disturbances in Amazonian floodplain forests: The collateral effects of hydropower production. Sci. Total Environ. 2019, 659, 587–598. [Google Scholar] [CrossRef]
- Schöngart, J.; Wittmann, F.; Resende, A.F.; Assahira, C.; Lobo, G.S.; Neves, J.R.D.; Rocha, M.; Mori, G.B.; Quaresma, A.C.; Demarchi, L.O.; et al. The shadow of Balbina dam: A synthesys of over 35 years of downstream impacts on floodplain forests in Central Amazonia. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 1117–1135. [Google Scholar] [CrossRef]
- Wittmann, F.; Damm, C.; Schöngart, J. Der Sandwich-Effekt: Einengung von Habitaten durch Staudämme gefährdet die größten und artenreichsten Flussauen der Erde. Auenmagazin 2019, 15, 49–53. [Google Scholar]
- Latrubesse, E.M.; Arima, E.Y.; Dunne, T.; Park, E.; Baker, V.R.; d’Horta, F.M.; Wight, C.; Wittmann, F.; Zuanon, J.; Baker, P.A.; et al. Damming the rivers of the Amazon basin. Nature 2017, 546, 363–369. [Google Scholar] [CrossRef] [Green Version]
- Latrubesse, E.M.; d’Horta, F.M.; Ribas, C.C.; Wittmann, F.; Zuanon, J.; Park, E.; Dunne, T.; Arima, E.Y.; Baker, P.A. Vulnerability of the biota in riverine and seasonally flooded habitats to damming of Amazonian Rivers. Aquat. Conserv. Mar. Freshw. Ecosyst. 2020, 31, 1136–1149. [Google Scholar] [CrossRef]
- Agostinho, A.A.; Tomaz, S.M.; Gomes, L.C. Threats for biodiversity in the floodplain of the Upper Paraná River: Effects of hydrological regulation by dams. Ecohydrol. Hydrobiol. 2004, 4, 255–268. [Google Scholar]
- Junk, W.J.; Piedade, M.T.F.; Cunha, C.N.; Wittmann, F.; Schöngart, J. Macrohabitat studies in large Brazilian floodplains to support sustainable development in the face of climate change. Ecohydrol. Hydrobiol. 2018, 18, 334–344. [Google Scholar] [CrossRef]
- McGrath, D.C.; Almeida, O.T.; Crossa, M.; Cardoso, A.; Cunha, M. Working towards community-based ecosystem management of the lower Amazon floodplain. PLEC News Views 2005, 6, 3–10. [Google Scholar]
- Lewis, S.L.; Brando, P.M.; Phillips, O.L.; Van der Heijden, G.M.F.; Nepstad, D. The 2010 Amazon drought. Science 2011, 331, 554. [Google Scholar] [CrossRef] [PubMed]
- Condit, R.; Engelbrecht, B.M.J.; Pino, D.; Pérez, R.; Turner, B.L. Species distributions in response to individual soil nutrients and seasonal drought across a community of tropical trees. Proc. Natl. Acad. Sci. USA 2013, 110, 5064–5068. [Google Scholar] [CrossRef]

| Habitat | Region/River | Lat./Long. | Size (ha) | Mean Flood Period (Days Year−1) | No. ind. | No. Species | Fisher’s Alpha | Author |
|---|---|---|---|---|---|---|---|---|
| IG | CA, Abacate River | −02.10, −58.43 | 1 | 70 | 747 | 59 | 15.02 | [44] |
| IG | CA, Abacate River | −02.09, −58.43 | 1 | 200 | 624 | 97 | 32.17 | [44] |
| IG | NA, Aracá River | −00.12, −63.29 | 1 | 52 | 1981 | 61 | 11.91 | [43] |
| IG | NA, Aracá River | −00.12, −63.29 | 0.5 | 68 | 865 | 51 | 11.85 | [43] |
| IG | CA, Cuyuni River | −00.42, −63.08 | 1 | 228 | 713 | 76 | 21.53 | [43] |
| IG | CA, Cuyuni River | −00.45, −63.10 | 0.5 | 122 | 982 | 36 | 7.34 | [43] |
| IG | CA, Jaú River | −01.83, −61.62 | 1 | 264 | 758 | 24 | 4.72 | [43] |
| IG | CA, Jaú River | −01.83, −61.61 | 1 | 216 | 609 | 46 | 11.55 | [43] |
| IG | CA, Jaú River | −01.83, −61.62 | 1 | 182 | 971 | 27 | 5.15 | [43] |
| IG | CA, Jaú River | −01.86, −61.59 | 1 | 191 | 758 | 55 | 13.63 | [43] |
| IG | CA, Jaú River | −01.86, −61.59 | 1 | 173 | 939 | 53 | 12.16 | [43] |
| IG | CA, Jaú River | −01.87, −61.59 | 1 | 124 | 986 | 54 | 12.28 | [43] |
| IG | CA, Jaú River | −01.84, −61.59 | 1 | 101 | 450 | 72 | 24.2 | [43] |
| IG | CA, Jaú River | −01.90, −61.46 | 1 | 67 | 726 | 63 | 16.56 | [43] |
| IG | CA, Jaú River | −01.94, −61.44 | 1 | 53 | 674 | 69 | 19.25 | [43] |
| IG | CA, Cuieiras River | −02.36, −60.19 | 0.5 | 188 | 462 | 33 | 8.13 | [43] |
| IG | CA, Cuieiras River | −02.37, −60.19 | 0.5 | 75 | 525 | 29 | 6.6 | [43] |
| IG | CA, Uatumã River | −02.13, 59.03 | 1 | 128 | 459 | 110 | 45.85 | [45] |
| IG | CA, Uatumã River | −02.15, 59.03 | 1 | 270 | 548 | 28 | 6.24 | [45] |
| IG | CA, Negro River | −02.76, −60.76 | 1 | 92 | 468 | 78 | 26.72 | [46] |
| IG | CA, Negro River | −02.69, −60.78 | 1 | 92 | 398 | 51 | 15.54 | [46] |
| IG | NA, Negro River | −00.69, −63.16 | 1 | 85 | 722 | 62 | 16.24 | [46] |
| IG | NA, Negro River | −00.63, −63.26 | 1 | 86 | 815 | 53 | 12.68 | [46] |
| IG | NA, Jufaris River | −00.92, −62.29 | 1 | 91 | 593 | 57 | 15.54 | [46] |
| IG | NA, Jufaris River | −01.09, −62.05 | 0.75 | 52 | 851 | 65 | 16.37 | [46] |
| IG | NA, Padauari River | −00.15, −64.04 | 0.5 | 227 | 644 | 23 | 4.66 | [43] |
| IG | NA, Padauari River | −00.15, −64.05 | 0.5 | 112 | 270 | 69 | 29.94 | [43] |
| IG | NA, Padauari River | −00.15, −64.04 | 0.5 | 100 | 309 | 72 | 29.51 | [43] |
| IG | NA, Negro River | −00.24, −64.24 | 0.5 | 101 | 302 | 64 | 24.83 | [43] |
| IG | NA, Negro River | −00.24, −64.24 | 0.5 | 99 | 269 | 50 | 18.09 | [43] |
| IG | NA, Negro River | −00.21, −64.25 | 0.5 | 39 | 328 | 57 | 19.93 | [43] |
| IG | NA, Negro River | −00.35, −63.91 | 1 | 90 | 504 | 64 | 19.43 | [46] |
| IG | NA, Negro River | −00.45, −64.78 | 1 | 79 | 500 | 66 | 20.37 | [46] |
| IG | NA, Negro River | −00.35, −64.31 | 1 | 74 | 573 | 59 | 16.45 | [46] |
| VZ | CA, Solimões River | −02.56, −64.70 | 1 | 93 | 566 | 94 | 32.16 | Unpublished |
| VZ | CA, Solimões River | −02.56, −64.69 | 1 | 160 | 507 | 31 | 7.28 | Unpublished |
| VZ | CA, Solimões River | −02.55, −64.69 | 1 | 114 | 462 | 49 | 13.86 | Unpublished |
| VZ | CA, Solimões River | −03.34, −60.11 | 0.875 | 115 | 421 | 64 | 21 | [47] |
| VZ | WA, Juruá River | −03.23, −66.05 | 0.5 | 161 | 354 | 73 | 27.9 | [40] |
| VZ | WA, Juruá River | −03.21, −66.00 | 0.5 | 112 | 285 | 51 | 18.09 | [40] |
| VZ | WA, Juruá River | −03.44, −66.04 | 0.5 | 156 | 358 | 60 | 20.62 | [40] |
| VZ | WA, Juruá River | −03.20, −65.99 | 0.5 | 115 | 300 | 78 | 34.23 | [40] |
| VZ | WA, Juruá River | −03.20, −66.01 | 0.5 | 132 | 348 | 74 | 28.77 | [40] |
| VZ | WA, Juruá River | −03.35, −66.02 | 0.5 | 170 | 409 | 47 | 13.71 | [40] |
| VZ * | WA, Jutaí River | −03.32, −67.44 | 0.5 | 170 | 297 | 29 | 7.95 | [40] |
| VZ * | WA, Jutaí River | −03.36, −67.49 | 0.5 | 131 | 413 | 59 | 18.83 | [40] |
| VZ * | WA, Jutaí River | −03.39, −67.49 | 0.5 | 110 | 368 | 78 | 30.27 | [40] |
| VZ * | WA, Jutaí River | −03.37, −67.50 | 0.5 | 200 | 261 | 40 | 13.18 | [40] |
| VZ * | WA, Jutaí River | −03.33, −67.44 | 0.5 | 169 | 695 | 58 | 15.05 | [40] |
| VZ * | WA, Jutaí River | −03.37, −67.48 | 0.5 | 82 | 356 | 84 | 34.69 | [40] |
| VZ | CA, Solimões River | −03.25, −59.97 | 1 | 190 | 486 | 38 | 9.65 | [47] |
| VZ | WA, Japurá River | −02.85, −64.91 | 1 | 162 | 662 | 37 | 8.46 | [47] |
| VZ | WA, Japurá River | −02.89, −64.88 | 1 | 129 | 841 | 36 | 7.64 | [48] |
| VZ | WA, Japurá River | −02.90, −64.88 | 1 | 152 | 487 | 48 | 13.21 | [48] |
| VZ | WA, Japurá River | −02.85, −64.91 | 1 | 139 | 461 | 89 | 32.83 | [47] |
| VZ | WA, Japurá River | −02.85, −64.92 | 1 | 139 | 462 | 108 | 44.35 | [48] |
| VZ | WA, Japurá River | −02.79, −65.06 | 1 | 141 | 504 | 86 | 29.81 | [48] |
| VZ | WA, Japurá River | −02.83, −65.04 | 1 | 87 | 444 | 149 | 78.69 | [47] |
| VZ | CA, Purus River | −04.13, −61.88 | 1 | 70 | 542 | 103 | 37.68 | [40] |
| VZ | CA, Purus River | −04.28, −61.85 | 1 | 90 | 603 | 76 | 23 | [40] |
| VZ | CA, Purus River | −04.36, −61.91 | 1 | 160 | 731 | 56 | 14.12 | [40] |
| VZ | CA, Purus River | −04.25, −61.75 | 1 | 90 | 457 | 85 | 30.76 | [40] |
| VZ | WA, Japurá River | −02.36, −65.47 | 1 | 227 | 486 | 78 | 26.25 | Unpublished |
| VZ | WA, Japurá River | −02.37, −65.47 | 1 | 166 | 607 | 71 | 20.85 | Unpublished |
| VZ | WA, Japurá River | −01.76, −65.76 | 1 | 49 | 618 | 71 | 20.71 | Unpublished |
| VZ | WA, Japurá River | −01.77, −65.76 | 1 | 140 | 481 | 63 | 19.38 | Unpublished |
| VZ * | WA, Tefé River | −04.13, −65.97 | 0.5 | 40 | 239 | 73 | 35.83 | [40] |
| VZ * | WA, Tefé River | −03.98, −65.01 | 0.5 | 31 | 320 | 69 | 27.03 | [40] |
| VZ * | WA, Tefé River | −04.12, −65.08 | 0.5 | 168 | 349 | 70 | 26.35 | [40] |
| VZ * | WA, Tefé River | −03.98, 65.01 | 0.5 | 187 | 303 | 59 | 21.86 | [40] |
| VZ * | WA, Tefé River | −04.17, −65.12 | 0.5 | 100 | 329 | 74 | 29.7 | [40] |
| VZ * | WA, Tefé River | −04.16, −65.01 | 0.5 | 118 | 281 | 62 | 24.61 | [40] |
| Igapó | Várzea | Total | |
|---|---|---|---|
| No. of inventoried plots | 34 | 38 | 72 |
| Inventoried area (ha) | 28.75 | 28.875 | 57.625 |
| Mean flood duration (days year−1) | 123 ± 64 | 130 ± 65 | |
| No. of individuals (total) | 22,323 | 17,093 | 39,416 |
| No. individuals ha−1 (mean, SD) | 656.5 ± 310 | 449.8 ± 139 | |
| No. of species ha−1 (mean, SD) | 56.9 ± 19.1 | 67.7 ± 23.2 | |
| Identified individuals (%, mean) | 77.79 | 87.1 | |
| No. of identified species (total) | 464 | 494 | 761 |
| Fisher’s alpha (mean, SD) | 16.84 ± 8.83 | 24.22 ± 12.88 | |
| The 12 most important species account for (%, OIV) | 20.03 | 18.34 |
| Igapó | ||||||
|---|---|---|---|---|---|---|
| Rank | Species | Abund. (Total) | Abund. (Rel.) | Dom. (Rel.) | Frequ. (Total) | Importance (Rel.) |
| 1 | Pouteria elegans (A. DC.) Baehni | 883 | 3.955 | 3.278 | 23 | 2.817 |
| 2 | Tachigali venusta Dwyer | 554 | 2.481 | 3.326 | 10 | 2.112 |
| 3 | Amanoa oblongifolia Müll. Arg. | 584 | 2.616 | 2.499 | 8 | 1.846 |
| 4 | Leptobalanus apetalus (E. Mey.) Sothers and Prance | 437 | 1.957 | 1.563 | 29 | 1.685 |
| 5 | Hevea spruceana (Benth.) Müll. Arg. | 445 | 1.993 | 2.378 | 12 | 1.669 |
| 6 | Macrolobium acaciifolium (Benth.) Benth. | 350 | 1.567 | 2.523 | 17 | 1.664 |
| 7 | Hymenopus heteromorphus (Benth.) Sothers and Prance | 463 | 2.074 | 1.688 | 20 | 1.607 |
| 8 | Gustavia augusta L. | 502 | 2.248 | 1.815 | 12 | 1.566 |
| 9 | Duroia velutina (Spruce ex Benth. and Hook f.) J. D. Hook. ex Schumann | 407 | 1.823 | 1.487 | 13 | 1.333 |
| 10 | Swartzia polyphylla DC. | 290 | 1.299 | 1.709 | 14 | 1.25 |
| 11 | Swartzia racemosa Benth. | 344 | 1.541 | 1.609 | 11 | 1.244 |
| 12 | Aldina heterophylla Spruce ex Benth. | 212 | 0.949 | 2.123 | 12 | 1.236 |
| Σ 13-593 | 16,852 | 75.491 | 73.996 | 79.967 | ||
| Várzea | ||||||
| 1 | Pseudobombax munguba (Mart.) Dugand | 577 | 3.375 | 4.823 | 22 | 3.027 |
| 2 | Luehea cymulosa Spruce ex Benth. | 514 | 3.007 | 3.241 | 16 | 2.296 |
| 3 | Eschweilera albiflora (DC.) Miers | 363 | 2.123 | 2.438 | 23 | 1.828 |
| 4 | Pouteria elegans (A. DC.) Baehni | 390 | 2.281 | 1.732 | 19 | 1.592 |
| 5 | Virola surinamensis (Rol. ex Rottb.) Warb. | 272 | 1.591 | 2.101 | 18 | 1.472 |
| 6 | Cecropia latiloba Miq. | 341 | 1.995 | 1.716 | 15 | 1.437 |
| 7 | Handroanthus barbatus (E. Mey.) Mattos | 267 | 1.562 | 1.678 | 13 | 1.254 |
| 8 | Mabea nitida Spruce ex Benth. | 263 | 1.539 | 1.286 | 16 | 1.155 |
| 9 | Pterocarpus rohrii Vahl | 243 | 1.421 | 1.215 | 19 | 1.133 |
| 10 | Hevea spruceana (Benth.) Müll. Arg. | 206 | 1.205 | 1.354 | 18 | 1.093 |
| 11 | Hevea brasiliensis (Willd. ex A. Juss.) Müll. Arg. | 213 | 1.246 | 1.245 | 17 | 1.057 |
| 12 | Eschweilera ovalifolia (DC.) Nied. | 173 | 1.012 | 1.333 | 17 | 1.009 |
| Σ 13-633 | 13,271 | 77.64 | 75.834 | 81.646 | ||
| Pioneer Stages | Early Secondary Stages | Late Secondary Stages | Late Stages | |
|---|---|---|---|---|
| Environmental factors | ||||
| Shear stress by hydraulic force | very high | high | intermediate | low |
| Mean inundation height (m) | 8–12 | 5–7 (230–270) | 3–5 (50–100) | <3 |
| Mean inundation duration (days year−1) | >300 | 230–270 | 50–100 | <50 |
| Sedimentation rates | dm–m | cm–dm | mm–cm | mm |
| Substrate grain size 1 | sand | fine sand | silt | clay |
| Rel. PAR (%) 2 | 70–100 | 30–70 | 5–30 | <5 |
| Water retention capacity | low | very low | intermediate | high |
| Biotic factors | ||||
| Vegetation type | macrophytes (i.e., semiaquatic grasses) | open shrub and tree formations | forest | forest |
| Tree density ha−1 (> 10 cm dbh) 1 | - | 100–200 | 800–1000 | 400–600 |
| Mean tree diameters (cm) 3 | - | 10–15 | 15–30 | >30 |
| Tree heights in upper canopy (m) 1 | - | 8–10 | 15–20 | >20 |
| Stratification | - | single | double | double or more |
| Max tree ages (years) 3 | 10–15 | 35–100 | >100 | |
| Individual crown area (m2) 4 | - | 30–60 | 60–200 | 200–800 |
| Aboveground root type 5 | stilt roots | stilt and tabular roots | tabular roots | |
| Wood density (g cm−3) 3 | - | <0.4 | intermediate | >0.5 |
| Biomass (Mg ha−1) | 70–100 6 | 18 ± 3 3 | 117 ± 9 3 | 239 ± 11 3 |
| NPP (Mg ha−1 year−1) 3,6 | 30–99 6 | 11.25 3 | 14.34 3 | 6.46 3 |
| Species richness (ha−1) | 1–5 herb species | 1–3 shrub or tree species | 10–25 tree species | > 80 tree species |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wittmann, F.; Householder, J.E.; Piedade, M.T.F.; Schöngart, J.; Demarchi, L.O.; Quaresma, A.C.; Junk, W.J. A Review of the Ecological and Biogeographic Differences of Amazonian Floodplain Forests. Water 2022, 14, 3360. https://doi.org/10.3390/w14213360
Wittmann F, Householder JE, Piedade MTF, Schöngart J, Demarchi LO, Quaresma AC, Junk WJ. A Review of the Ecological and Biogeographic Differences of Amazonian Floodplain Forests. Water. 2022; 14(21):3360. https://doi.org/10.3390/w14213360
Chicago/Turabian StyleWittmann, Florian, John Ethan Householder, Maria Teresa Fernandez Piedade, Jochen Schöngart, Layon Oreste Demarchi, Adriano Costa Quaresma, and Wolfgang J. Junk. 2022. "A Review of the Ecological and Biogeographic Differences of Amazonian Floodplain Forests" Water 14, no. 21: 3360. https://doi.org/10.3390/w14213360
APA StyleWittmann, F., Householder, J. E., Piedade, M. T. F., Schöngart, J., Demarchi, L. O., Quaresma, A. C., & Junk, W. J. (2022). A Review of the Ecological and Biogeographic Differences of Amazonian Floodplain Forests. Water, 14(21), 3360. https://doi.org/10.3390/w14213360