
Influence of Spatial Extent on Contemporary and Future Threat Evaluation for Imperiled Fluvial Fishes and Mussels



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area, Spatial Framework, and Climate Data
2.2. Habitat Suitability Models and Climate Change Projections
2.3. Contemporary Human Landscape Disturbance
2.4. Contemporary Stream Fragmentation
2.5. Combining Contemporary and Future Threats
3. Results
3.1. Contemporary Human Landscape Disturbance
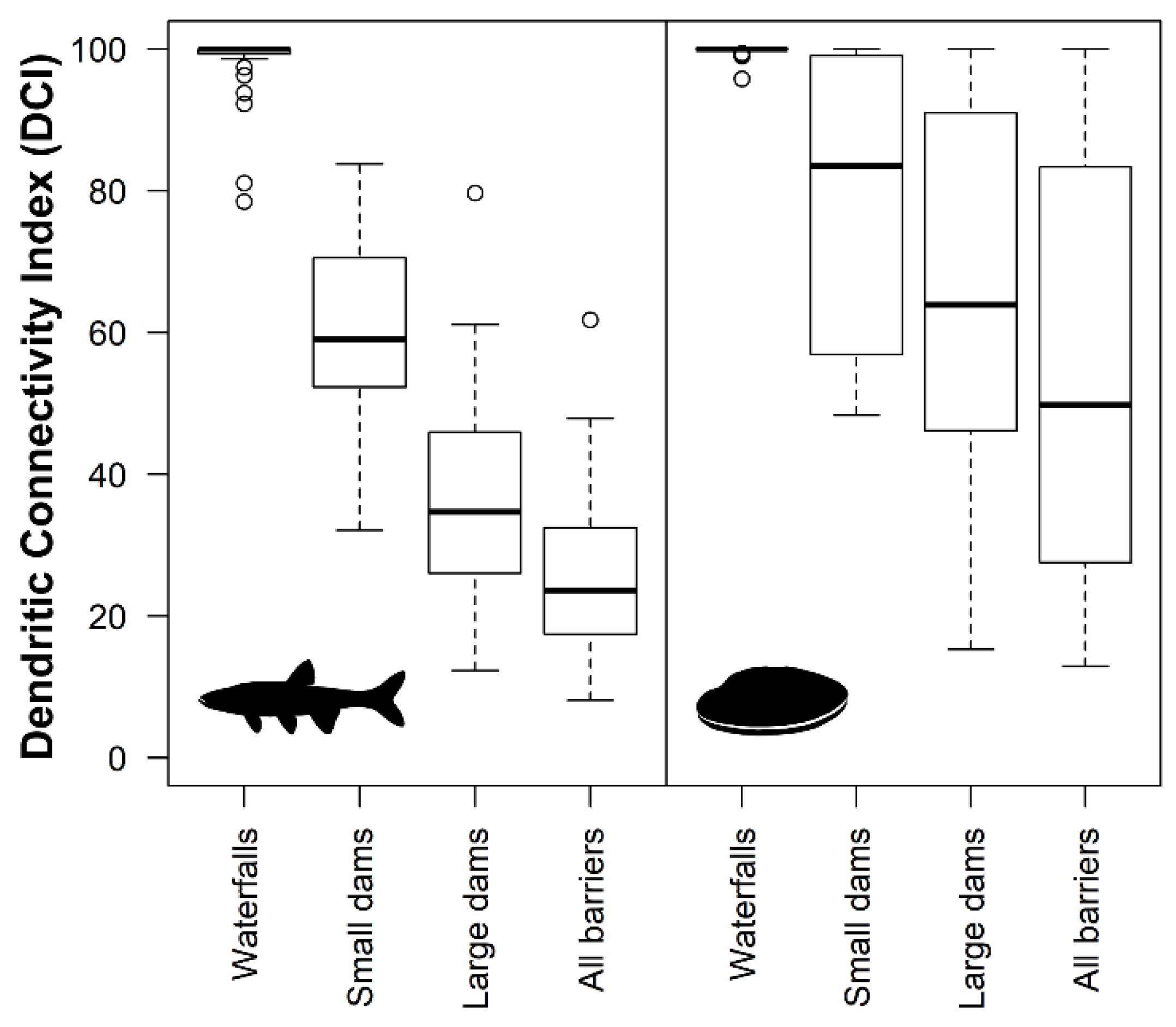
3.2. Contemporary Stream Habitat Fragmentation
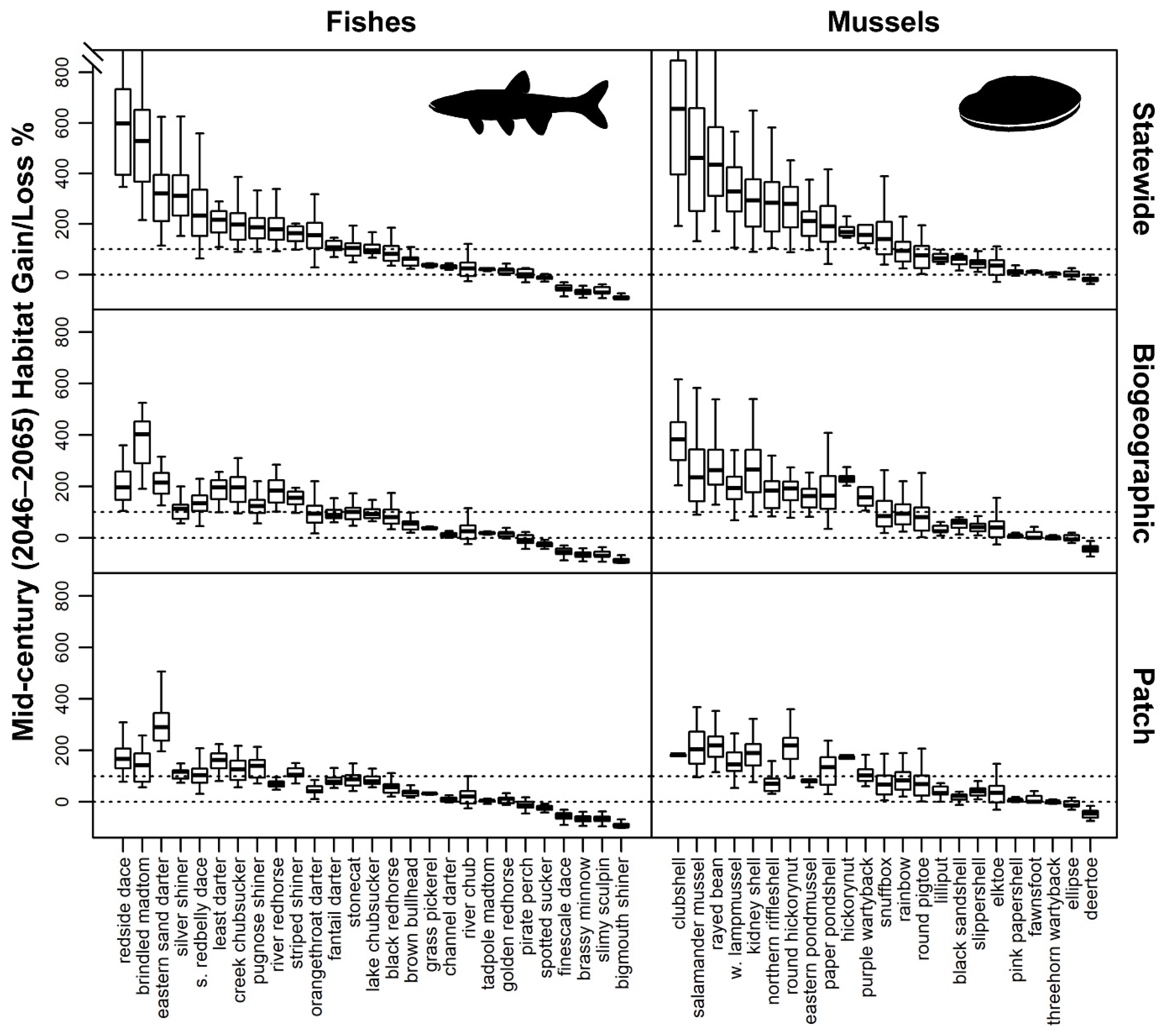
3.3. Future Habitat Suitability under Climate Change
3.4. Combining Contemporary and Future Threats
4. Discussion
4.1. Incorporating Multiple Taxa in Threat Assessments vs. Use of a Surrogate Taxon
4.2. Assessing Multiple Contemporary and Future Threats
4.3. Influence of Spatial Extent on Threat Assessments for Imperiled Fluvial Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.H.; Soto, D.; Stiassny, M.L.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Su, G.; Logez, M.; Xu, J.; Tao, S.; Villéger, S.; Brosse, S. Human impacts on global freshwater fish biodiversity. Science 2021, 371, 835–838. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Zarfl, C.; Bremerich, V.; David, J.N.; Hogan, Z.; Kalinkat, G.; Tockner, K.; Jähnig, S.C. The global decline of freshwater megafauna. Glob. Chang. Biol. 2019, 25, 3883–3892. [Google Scholar] [CrossRef]
- Lydeard, C.; Cowie, R.H.; Ponder, W.F.; Bogan, A.E.; Bouchet, P.; Clark, S.A.; Cummings, K.S.; Frest, T.J.; Gargominy, O.; Herbert, D.G.; et al. The global decline of nonmarine mollusks. BioScience 2004, 54, 321–330. [Google Scholar] [CrossRef]
- Lopes-Lima, M.; Burlakova, L.E.; Karatayev, A.Y.; Mehler, K.; Seddon, M.; Sousa, R. Conservation of freshwater bivalves at the global scale: Diversity, threats and research needs. Hydrobiologia 2018, 810, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Ferreira-Rodríguez, N.; Akiyama, Y.B.; Aksenova, O.V.; Araujo, R.; Barnhart, M.C.; Bespalaya, Y.V.; Bogan, A.E.; Bolotov, I.N.; Budha, P.B.; Clavijo, C.; et al. Research priorities for freshwater mussel conservation assessment. Biol. Conserv. 2019, 231, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Jelks, H.L.; Walsh, S.J.; Burkhead, N.M.; Contreras-Balderas, S.; Diaz-Pardo, E.; Hendrickson, D.A.; Lyons, J.; Mandrak, N.E.; McCormick, F.; Nelson, J.S.; et al. Conservation status of imperiled North American freshwater and diadromous fishes. Fisheries 2008, 33, 372–407. [Google Scholar] [CrossRef]
- Wilcove, D.S.; Rothstein, D.; Dubow, J.; Phillips, A.; Losos, E. Quantifying threats to imperiled species in the United States. BioScience 1998, 48, 607–615. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.D.; Warren, M.L., Jr.; Cummings, K.S.; Harris, J.L.; Neves, R.J. Conservation status of freshwater mussels of the United States and Canada. Fisheries 1993, 18, 6–22. [Google Scholar] [CrossRef]
- Strayer, D.L.; Downing, J.A.; Haag, W.R.; King, T.L.; Layzer, J.B.; Newton, T.J.; Nichols, J.S. Changing perspectives on pearly mussels, North America’s most imperiled animals. BioScience 2004, 54, 429–439. [Google Scholar] [CrossRef]
- Lopes-Lima, M.; Sousa, R.; Geist, J.; Aldridge, D.C.; Araujo, R.; Bergengren, J.; Bespalaya, Y.; Bódis, E.; Burlakova, L.; Van Damme, D.; et al. Conservation status of freshwater mussels in Europe: State of the art and future challenges. Biol. Rev. 2017, 92, 572–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelicice, F.M.; Azevedo-Santos, V.M.; Vitule, J.R.; Orsi, M.L.; Lima Junior, D.P.; Magalhães, A.L.; Pompeu, P.S.; Petrere, M., Jr.; Agostinho, A.A. Neotropical freshwater fishes imperilled by unsustainable policies. Fish Fish. 2017, 18, 1119–1133. [Google Scholar] [CrossRef]
- Zieritz, A.; Bogan, A.E.; Froufe, E.; Klishko, O.; Kondo, T.; Kovitvadhi, U.; Kovitvadhi, S.; Lee, J.H.; Lopes-Lima, M.; Pfeiffer, J.M.; et al. Diversity, biogeography and conservation of freshwater mussels (Bivalvia: Unionida) in East and Southeast Asia. Hydrobiologia 2018, 810, 29–44. [Google Scholar] [CrossRef]
- Palmer, M.A.; Lettenmaier, D.P.; Poff, N.L.; Postel, S.L.; Richter, B.; Warner, R. Climate change and river ecosystems: Protection and adaptation options. Environ. Manag. 2009, 44, 1053–1068. [Google Scholar] [CrossRef]
- Woodward, G.; Perkins, D.M.; Brown, L.E. Climate change and freshwater ecosystems: Impacts across multiple levels of organization. Philos. T. R. Soc. B 2010, 365, 2093–2106. [Google Scholar] [CrossRef] [Green Version]
- Strayer, D.L.; Dudgeon, D. Freshwater biodiversity conservation: Recent progress and future challenges. J. N. Am. Benthol. Soc. 2010, 29, 344–358. [Google Scholar] [CrossRef] [Green Version]
- Ormerod, S.J.; Dobson, M.; Hildrew, A.G.; Townsend, C. Multiple stressors in freshwater ecosystems. Freshw. Biol. 2010, 55, 1–4. [Google Scholar] [CrossRef]
- Titeux, N.; Henle, K.; Mihoub, J.B.; Regos, A.; Geijzendorffer, I.R.; Cramer, W.; Verburg, P.H.; Brotons, L. Biodiversity scenarios neglect future land-use changes. Glob. Chang. Biol. 2016, 22, 2505–2515. [Google Scholar] [CrossRef] [Green Version]
- Nelson, K.C.; Palmer, M.A.; Pizzuto, J.E.; Moglen, G.E.; Angermeier, P.L.; Hilderbrand, R.H.; Dettinger, M.; Hayhoe, K. Forecasting the combined effects of urbanization and climate change on stream ecosystems: From impacts to management options. J. Appl. Ecol. 2009, 46, 154–163. [Google Scholar] [CrossRef] [Green Version]
- Mantyka-Pringle, C.S.; Martin, T.G.; Moffatt, D.B.; Udy, J.; Olley, J.; Saxton, N.; Sheldon, F.; Bunn, S.E.; Rhodes, J.R. Prioritizing management actions for the conservation of freshwater biodiversity under changing climate and land-cover. Biol. Conserv. 2016, 197, 80–89. [Google Scholar] [CrossRef]
- Radinger, J.; Hölker, F.; Horký, P.; Slavík, O.; Dendoncker, N.; Wolter, C. Synergistic and antagonistic interactions of future land use and climate change on river fish assemblages. Glob. Chang. Biol. 2016, 22, 1505–1522. [Google Scholar] [CrossRef]
- Kennedy, C.M.; Oakleaf, J.R.; Theobald, D.M.; Baruch-Mordo, S.; Kiesecker, J. Managing the middle: A shift in conservation priorities based on the global human modification gradient. Glob. Chang. Biol. 2019, 25, 811–826. [Google Scholar] [CrossRef] [PubMed]
- Rojas, I.M.; Jennings, M.K.; Conlisk, E.; Syphard, A.D.; Mikesell, J.; Kinoshita, A.M.; West, K.; Stow, D.; Storey, E.; De Guzman, M.E.; et al. A landscape-scale framework to identify refugia from multiple stressors. Conserv. Biol. 2022, 36, e13834. [Google Scholar] [CrossRef] [PubMed]
- Geist, J. Integrative freshwater ecology and biodiversity conservation. Ecol. Indic. 2011, 11, 1507–1516. [Google Scholar] [CrossRef]
- Olden, J.D.; Kennard, M.J.; Leprieur, F.; Tedesco, P.A.; Winemiller, K.O.; García-Berthou, E. Conservation biogeography of freshwater fishes: Recent progress and future challenges. Divers. Distrib. 2010, 16, 496–513. [Google Scholar] [CrossRef]
- Strayer, D.L. Challenges for freshwater invertebrate conservation. J. N. Am. Benthol. Soc. 2006, 25, 271–287. [Google Scholar] [CrossRef]
- Collier, K.J.; Probert, P.K.; Jeffries, M. Conservation of aquatic invertebrates: Concerns, challenges and conundrums. Aquat. Conserv. 2016, 26, 817–837. [Google Scholar] [CrossRef]
- Infante, D.M.; David Allan, J.; Linke, S.; Norris, R.H. Relationship of fish and macroinvertebrate assemblages to environmental factors: Implications for community concordance. Hydrobiologia 2009, 623, 87–103. [Google Scholar] [CrossRef] [Green Version]
- Heino, J.; Virkkala, R.; Toivonen, H. Climate change and freshwater biodiversity: Detected patterns, future trends and adaptations in northern regions. Biol. Rev. 2009, 84, 39–54. [Google Scholar] [CrossRef]
- Oikonomou, A.; Stefanidis, K. α-and β-diversity patterns of macrophytes and freshwater fishes are driven by different factors and processes in lakes of the unexplored southern Balkan biodiversity hotspot. Water 2020, 12, 1984. [Google Scholar] [CrossRef]
- Stewart, D.R.; Underwood, Z.E.; Rahel, F.J.; Walters, A.W. The effectiveness of surrogate taxa to conserve freshwater biodiversity. Conserv. Biol. 2018, 32, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Linke, S.; Turak, E.; Nel, J. Freshwater conservation planning: The case for systematic approaches. Freshw. Biol. 2011, 56, 6–20. [Google Scholar] [CrossRef]
- Du Toit, J.T. Considerations of scale in biodiversity conservation. Anim. Conserv. 2010, 13, 229–236. [Google Scholar] [CrossRef]
- Razgour, O.; Hanmer, J.; Jones, G. Using multi-scale modelling to predict habitat suitability for species of conservation concern: The grey long-eared bat as a case study. Biol. Conserv. 2011, 144, 2922–2930. [Google Scholar] [CrossRef] [Green Version]
- Cheok, J.; Weeks, R.; Pressey, R.L. Identifying the strengths and weaknesses of conservation planning at different scales: The Coral Triangle as a case study. Ecol. Soc. 2019, 24, 24. [Google Scholar] [CrossRef] [Green Version]
- Hermoso, V.; Kennard, M.J.; Linke, S. Integrating multidirectional connectivity requirements in systematic conservation planning for freshwater systems. Divers. Distrib. 2012, 18, 448–458. [Google Scholar] [CrossRef] [Green Version]
- Game, E.T.; Lipsett-Moore, G.; Saxon, E.; Peterson, N.; Sheppard, S. Incorporating climate change adaptation into national conservation assessments. Glob. Chang. Biol. 2011, 17, 3150–3160. [Google Scholar] [CrossRef]
- Schwenk, W.S.; Donovan, T.M. A multispecies framework for landscape conservation planning. Conserv. Biol. 2011, 25, 1010–1021. [Google Scholar] [CrossRef]
- Poiani, K.A.; Richter, B.D.; Anderson, M.G.; Richter, H.E. Biodiversity conservation at multiple scales: Functional sites, landscapes, and networks. BioScience 2000, 50, 133–146. [Google Scholar] [CrossRef] [Green Version]
- Quist, M.C.; Rahel, F.J.; Hubert, W.A. Hierarchical faunal filters: An approach to assessing effects of habitat and nonnative species on native fishes. Ecol. Freshw. Fish 2005, 14, 24–39. [Google Scholar] [CrossRef]
- Zorn, T.G.; Seelbach, P.W.; Wiley, M.J. Distributions of stream fishes and their relationship to stream size and hydrology in Michigan’s Lower Peninsula. Trans. Am. Fish. Soc. 2002, 131, 70–85. [Google Scholar] [CrossRef]
- Wehrly, K.E.; Wiley, M.J.; Seelbach, P.W. Classifying regional variation in thermal regime based on stream fish community patterns. Trans. Am. Fish. Soc. 2003, 132, 18–38. [Google Scholar] [CrossRef]
- Daniel, W.M.; Cooper, A.R.; Badra, P.J.; Infante, D.M. Predicting habitat suitability for eleven imperiled fluvial freshwater mussels. Hydrobiologia 2018, 809, 265–283. [Google Scholar] [CrossRef]
- Seelbach, P.W.; Hinz, L.C.; Wiley, M.J.; Cooper, A.R. Use of Multiple Linear Regression to Estimate Flow Regimes for All Rivers across Illinois, Michigan, and Wisconsin; Fisheries Research Report 2095; Michigan Department of Natural Resources: Lansing, MI, USA, 2011. [Google Scholar]
- Wehrly, K.E.; Brenden, T.O.; Wang, L. A comparison of statistical approaches for predicting stream temperatures across heterogeneous landscapes. J. Am. Water Resour. Assoc. 2009, 45, 986–997. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Eagle, A.C.; Hay-Chmielewski, E.M.; Cleveland, K.T.; Derosier, A.L.; Herbert, M.E.; Rustem, R.A. Michigan’s Wildlife Action Plan; Michigan Department of Natural Resources: Lansing, MI, USA, 2005. [Google Scholar]
- Lyons, J.; Zorn, T.; Stewart, J.; Seelbach, P.; Wehrly, K.; Wang, L. Defining and characterizing coolwater streams and their fish assemblages in Michigan and Wisconsin, USA. N. Am. J. Fish. Manag. 2009, 29, 1130–1151. [Google Scholar] [CrossRef]
- Wang, L.; Brenden, T.; Seelbach, P.; Cooper, A.; Allan, D.; Clark, R.; Wiley, M. Landscape based identification of human disturbance gradients and reference conditions for Michigan streams. Environ. Monit. Assess. 2008, 141, 1–17. [Google Scholar] [CrossRef]
- Muggeo, V.M.R. Segmented: An R package to fit regression models with broken-line relationships. R news. 2008, 8, 20–25. [Google Scholar]
- Brenden, T.O.; Wang, L.; Su, Z. Quantitative identification of disturbance thresholds in support of aquatic resource management. Environ. Manag. 2008, 42, 821–832. [Google Scholar] [CrossRef]
- Ostroff, A.; Wieferich, D.; Cooper, A.; Infante, D. National Anthropogenic Barrier Dataset (NABD); U.S. Geological Survey, Aquatic GAP Program: Denver, CO, USA, 2013. [Google Scholar] [CrossRef]
- Wieferich, D.J. Waterfalls Linked to the National Hydrography Datasets; U.S. Geological Survey, Aquatic GAP Program: Denver, CO, USA, 2016. [Google Scholar] [CrossRef]
- Cooper, A.R.; Infante, D.M.; Daniel, W.M.; Wehrly, K.E.; Wang, L.; Brenden, T.O. Assessment of dam effects on streams and fish assemblages of the conterminous USA. Sci. Total Environ. 2017, 586, 879–889. [Google Scholar] [CrossRef]
- Cote, D.; Kehler, D.G.; Bourne, C.; Wiersma, Y.F. A new measure of longitudinal connectivity for stream networks. Landsc. Ecol. 2009, 24, 101–113. [Google Scholar] [CrossRef] [Green Version]
- Mahlum, S.; Kehler, D.; Cote, D.; Wiersma, Y.F.; Stanfield, L. Assessing the biological relevance of aquatic connectivity to stream fish communities. Can. J. Fish. Aquat. Sci. 2014, 71, 1852–1863. [Google Scholar] [CrossRef]
- Dean, E.M.; Cooper, A.R.; Wang, L.; Daniel, W.; David, S.; Ernzen, C.; Gido, K.B.; Hale, E.; Haxton, T.J.; Kelso, W.; et al. The North American Freshwater Migratory Fish Database (NAFMFD): Characterizing the migratory life histories of freshwater fishes of Canada, the United States and Mexico. J. Biogeogr. 2022, 49, 1193–1203. [Google Scholar] [CrossRef]
- Sætersdal, M.; Gjerde, I. Prioritising conservation areas using species surrogate measures: Consistent with ecological theory? J. Appl. Ecol. 2011, 48, 1236–1240. [Google Scholar] [CrossRef]
- Larsen, S.; Mancini, L.; Pace, G.; Scalici, M.; Tancioni, L. Weak concordance between fish and macroinvertebrates in Mediterranean streams. PLoS ONE 2012, 7, e51115. [Google Scholar] [CrossRef] [Green Version]
- Westgate, M.J.; Barton, P.S.; Lane, P.W.; Lindenmayer, D.B. Global meta-analysis reveals low consistency of biodiversity congruence relationships. Nat. Commun. 2014, 5, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Backus-Freer, J.; Pyron, M. Concordance among fish and macroinvertebrate assemblages in streams of Indiana, USA. Hydrobiologia 2015, 758, 141–150. [Google Scholar] [CrossRef]
- Schinegger, R.; Palt, M.; Segurado, P.; Schmutz, S. Untangling the effects of multiple human stressors and their impacts on fish assemblages in European running waters. Sci. Total Environ. 2016, 573, 1079–1088. [Google Scholar] [CrossRef] [Green Version]
- Birk, S.; Chapman, D.; Carvalho, L.; Spears, B.M.; Andersen, H.E.; Argillier, C.; Auer, S.; Baattrup-Pedersen, A.; Banin, L.; Beklioğlu, M.; et al. Impacts of multiple stressors on freshwater biota across spatial scales and ecosystems. Nat. Ecol. Evol. 2020, 4, 1060–1068. [Google Scholar] [CrossRef]
- Feio, M.J.; Hughes, M.; Callisto, M.; Nichols, S.; Odume, O.; Quintella, B.; Kuemmerlen, M.; Aguiar, F.; Almeida, S.; Alonso-Eguíalis, P.; et al. The Biological Assessment and Rehabilitation of the World’s Rivers: An Overview. Water 2021, 13, 371. [Google Scholar] [CrossRef]
- Townsend, C.R.; Uhlmann, S.S.; Matthaei, C.D. Individual and combined responses of stream ecosystems to multiple stressors. J. Appl. Ecol. 2008, 45, 1810–1819. [Google Scholar] [CrossRef]
- Guisan, A.; Graham, C.H.; Elith, J.; Huettmann, F.; NCEAS Species Distribution Modelling Group. Sensitivity of predictive species distribution models to change in grain size. Divers. Distrib. 2007, 13, 332–340. [Google Scholar] [CrossRef]
- Whittaker, R.J.; Araújo, M.B.; Jepson, P.; Ladle, R.J.; Watson, J.E.; Willis, K.J. Conservation biogeography: Assessment and prospect. Divers. Distrib. 2005, 11, 3–23. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Ann. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- George, A.L.; Kuhajda, B.R.; Williams, J.D.; Cantrell, M.A.; Rakes, P.L.; Shute, J.R. Guidelines for propagation and translocation for freshwater fish conservation. Fisheries 2009, 34, 529–545. [Google Scholar] [CrossRef]
- Thomas, C.D. Translocation of species, climate change, and the end of trying to recreate past ecological communities. Trends Ecol. Evol. 2011, 26, 216–221. [Google Scholar] [CrossRef]
- Buisson, L.; Thuiller, W.; Lek, S.; Lim, P.U.Y.; Grenouillet, G. Climate change hastens the turnover of stream fish assemblages. Glob. Chang. Biol. 2008, 14, 2232–2248. [Google Scholar] [CrossRef]
- Ackerly, D.D.; Loarie, S.R.; Cornwell, W.K.; Weiss, S.B.; Hamilton, H.; Branciforte, R.; Kraft, N.J.B. The geography of climate change: Implications for conservation biogeography. Divers. Distrib. 2010, 16, 476–487. [Google Scholar] [CrossRef]
- Tracy, E.E.; Infante, D.M.; Cooper, A.R.; Taylor, W.W. An ecological resilience index to improve conservation action for stream fish habitat. Aquat. Conserv. 2022, 32, 951–966. [Google Scholar] [CrossRef]
- Brenden, T.O.; Clark, R.D.; Cooper, A.R.; Seelbach, P.W.; Wang, L.; Aichele, S.S.; Bissell, E.G.; Stewart, J.S. A GIS framework for collecting, managing, and analyzing multiscale landscape variables across large regions for river conservation and management. In Landscape influences on stream habitats and biological assemblages; Hughes, R.M., Wang, L., Seelbach, P.W., Eds.; American Fisheries Society: Bethesda, Maryland, 2006; Symposium 48; pp. 49–74. [Google Scholar]
- U.S. Geological Survey National Elevation Dataset. Available online: https://www.sciencebase.gov/catalog/item/4fcf8fd4e4b0c7fe80e81504#:~:text=The%20National%20Elevation%20Dataset%20 (accessed on 26 October 2022).
- U.S. Geological Survey National Hydrography Dataset. Available online: https://www.usgs.gov/national-hydrography/national-hydrography-dataset (accessed on 26 October 2022).
- Farrand, W.R.; Bell, D.L. Quaternary Geology of Michigan, Michigan Department of Natural Resources, Geological Survey Division. 1982. Available online: https://ngmdb.usgs.gov/Prodesc/proddesc_71889.htm (accessed on 26 October 2022).
- U.S. Department of Agriculture, State Soil Geographic Database (STATSGO) v2. Available online: https://catalog.data.gov/dataset/u-s-general-soil-map-statsgo2 (accessed on 26 October 2022).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Period | ||||
|---|---|---|---|---|
| Model (Units) | Statistic | Current | Mid-Century | Late Century |
| July mean stream temperature (Celsius) | Mean (SD) | 17.61 (2.39) | 19.16 (2.38) | 19.70 (2.38) |
| Range | 8.15–26.39 | 9.60–27.94 | 10.17–28.49 | |
| Median annual stream discharge (cms) | Mean (SD) | 1.41 (7.47) | 1.92 (10.26) | 2.20 (11.77) |
| Range | 0–151.62 | 0–196.51 | 0–219.27 | |
| Stream high flow yield (cms/km2) | Mean (SD) | 0.0851 (0.2442) | 0.0920 (0.2636) | 0.095 (0.2735) |
| Range | 0.0005–25.2262 | 0.0005–26.9667 | 0.0005–27.8942 | |
| Stream low flow yield (cms/km2) | Mean (SD) | 0.0015 (0.0021) | 0.0024 (0.0034) | 0.0030 (0.0043) |
| Range | 0–0.0289 | 0–0.0447 | 0–0.0565 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cooper, A.R.; Wehrly, K.E.; Yeh, S.-K.; Infante, D.M. Influence of Spatial Extent on Contemporary and Future Threat Evaluation for Imperiled Fluvial Fishes and Mussels. Water 2022, 14, 3464. https://doi.org/10.3390/w14213464
Cooper AR, Wehrly KE, Yeh S-K, Infante DM. Influence of Spatial Extent on Contemporary and Future Threat Evaluation for Imperiled Fluvial Fishes and Mussels. Water. 2022; 14(21):3464. https://doi.org/10.3390/w14213464
Chicago/Turabian StyleCooper, Arthur R., Kevin E. Wehrly, Sung-Kang Yeh, and Dana M. Infante. 2022. "Influence of Spatial Extent on Contemporary and Future Threat Evaluation for Imperiled Fluvial Fishes and Mussels" Water 14, no. 21: 3464. https://doi.org/10.3390/w14213464
APA StyleCooper, A. R., Wehrly, K. E., Yeh, S. -K., & Infante, D. M. (2022). Influence of Spatial Extent on Contemporary and Future Threat Evaluation for Imperiled Fluvial Fishes and Mussels. Water, 14(21), 3464. https://doi.org/10.3390/w14213464