
COVID-19 Lockdowns Promoted Recovery of the Yangtze River’s Aquatic Ecosystem

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection
2.3. Data Analysis
2.3.1. Statistical Analyses
2.3.2. Ecological Types
2.3.3. Community Diversity
2.3.4. Community Stability
3. Results
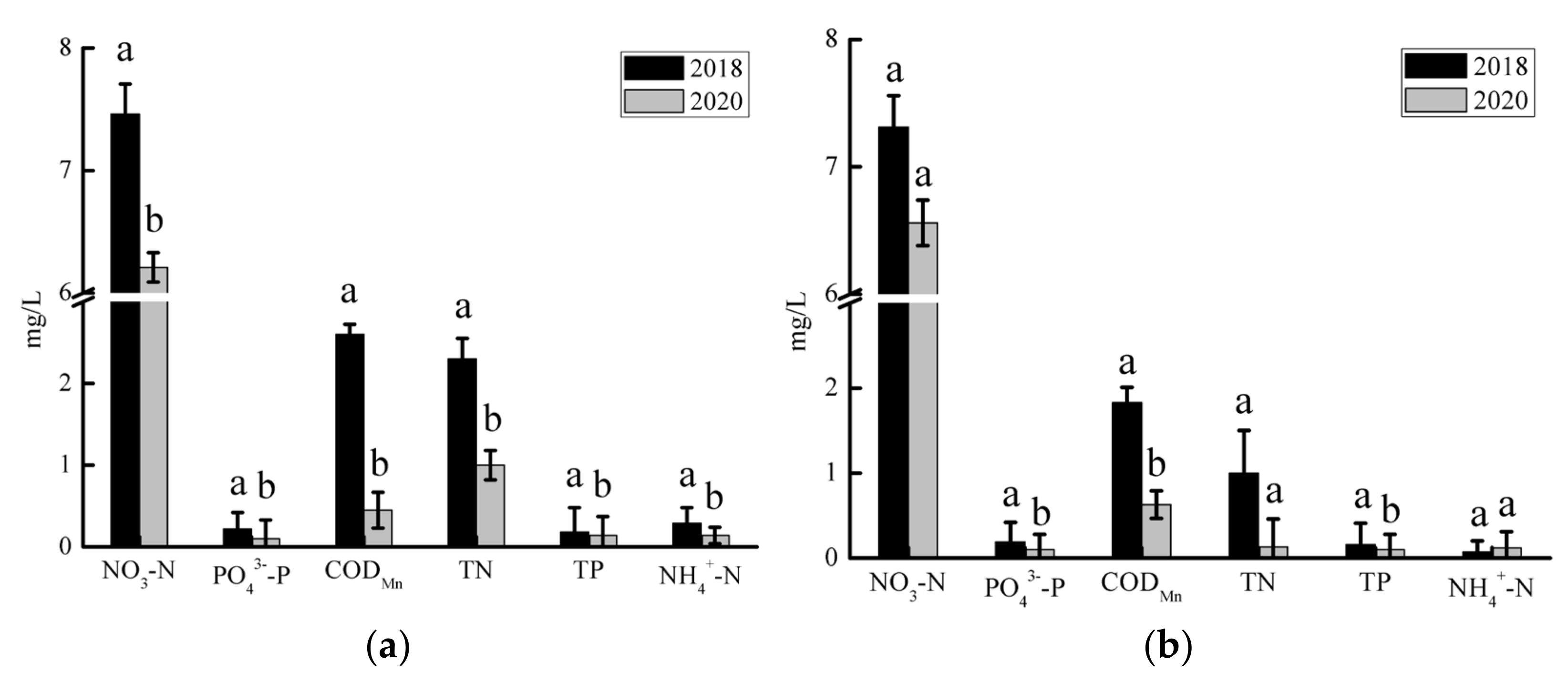
3.1. Spatiotemporal Variations of Water Quality
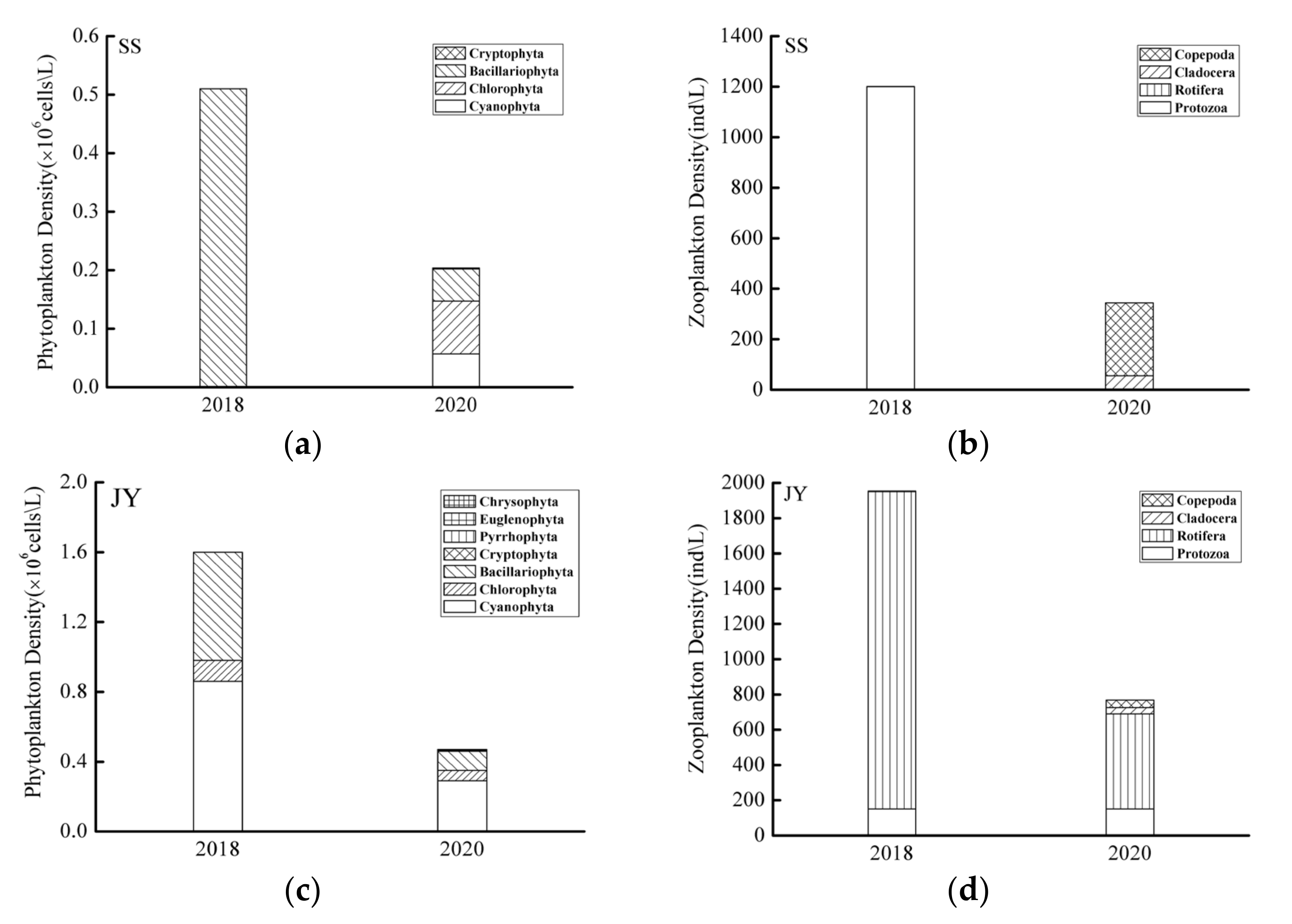
3.2. Spatiotemporal Variations of Phytoplankton and Zooplankton Density
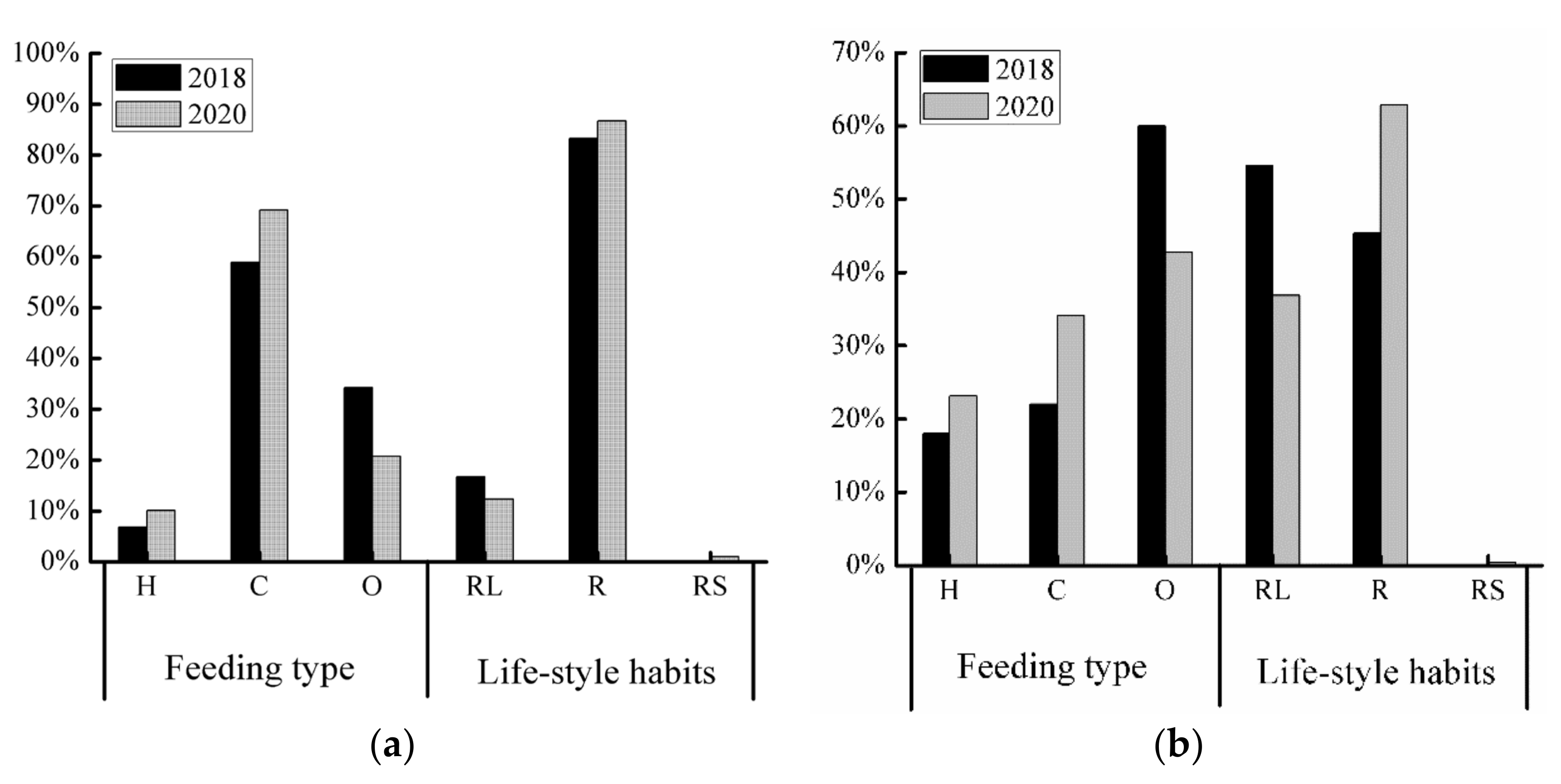
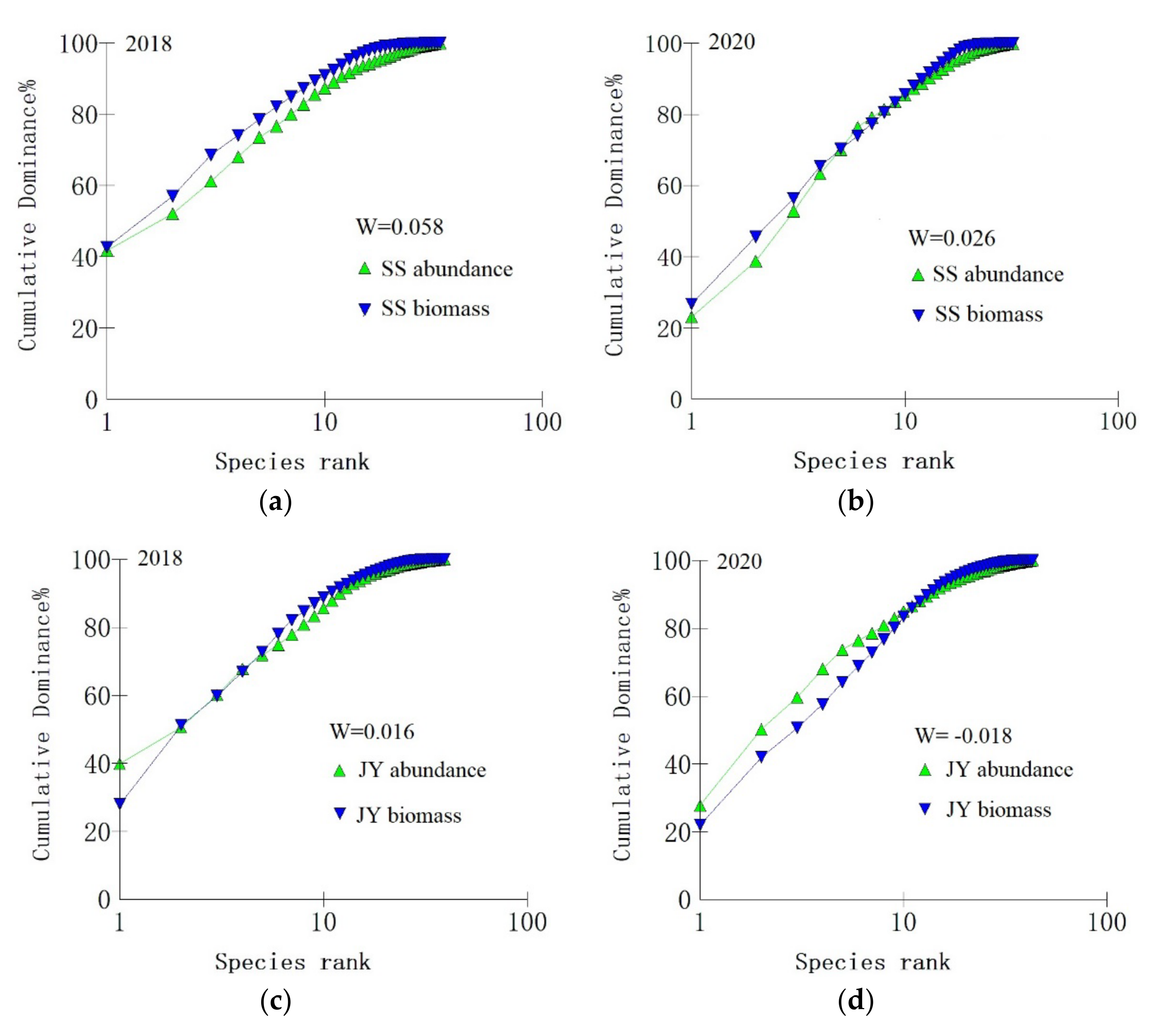
3.3. Spatiotemporal Variations of the Fish Community Structure and Diversity
3.3.1. Ecological Type
3.3.2. Community Stability
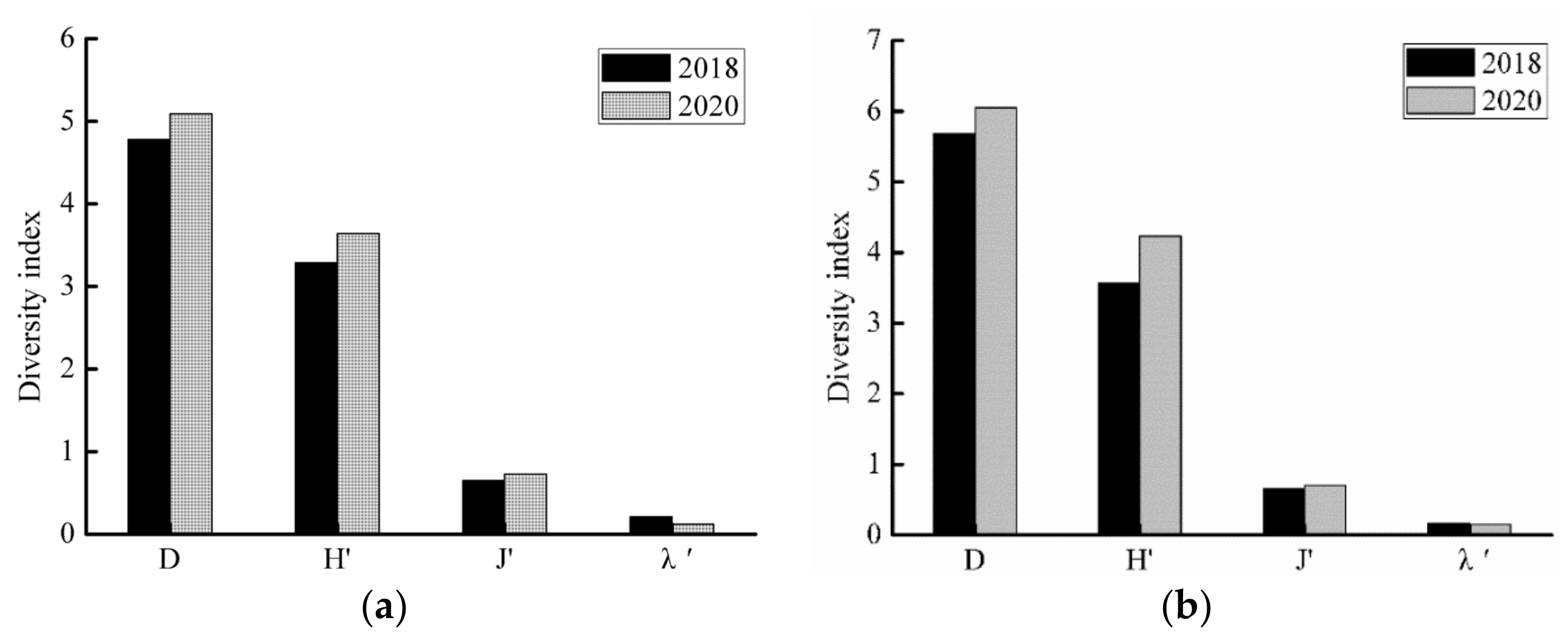
3.3.3. Diversity Characteristics
4. Discussion
4.1. Possible Reasons for Water Quality Improvements
4.2. Plankton Response to Water Quality Changes
4.3. Possible Reasons for Changes in Fish Diversity and Community Structure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Costanza, R.; De Groot, R.; Sutton, P.; Van der Ploeg, S.; Anderson, S.J.; Kubiszewski, I.; Turner, R.K. Changes in the global value of ecosystem services. Glob. Environ. Chang. 2014, 26, 152–158. [Google Scholar] [CrossRef]
- Tickner, D.; Opperman, J.J.; Abell, R.; Acreman, M.; Arthington, A.H.; Bunn, S.E.; Cooke, S.J.; Dalton, J.; Darwall, W.; Edwards, G.; et al. Bending the curve of global freshwater biodiversity loss: An emergency recovery plan. Bioscience 2020, 70, 330–342. [Google Scholar] [CrossRef] [Green Version]
- Collen, B.; Whitton, F.; Dyer, E.E.; Baillie, J.E.; Cumberlidge, N.; Darwall, W.R.; Böhm, M. Global patterns of freshwater species diversity, threat and endemism. Glob. Ecol. Biogeogr. 2014, 23, 40–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, F.; Bremerich, V.; Zarfl, C.; Geldmann, J.; Langhans, S.D.; David, J.N.; Jähnig, S.C. Freshwater megafauna diversity: Patterns, status and threats. Divers. Distrib. 2018, 24, 1395–1404. [Google Scholar] [CrossRef] [Green Version]
- Báldi, A.; Vári, Á. Freshwater ecosystems: Research, policy and applications. Biol. Futura 2020, 71, 333–336. [Google Scholar] [CrossRef]
- Vörösmarty, C.J.; McIntyre, P.B.; Gessner, M.O.; Dudgeon, D.; Prusevich, A.; Green, P.; Davies, P.M. Global threats to human water security and river biodiversity. Nature 2010, 467, 555–561. [Google Scholar] [CrossRef] [Green Version]
- Grill, G.; Lehner, B.; Thieme, M.; Geenen, B.; Tickner, D.; Antonelli, F.; Babu, S.; Borrelli, P.; Cheng, L.; Crochetiere, H.; et al. Mapping the world’s free-flowing rivers. Nature 2019, 569, 215–221. [Google Scholar] [CrossRef]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Lévêque, C. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 8, 163–182. [Google Scholar] [CrossRef]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.J.; Co-Oke, S.J. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef] [Green Version]
- Lai, C.C.; Shih, T.P.; Ko, W.C.; Tang, H.J.; Hsueh, P.R. Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and coronavirus disease-2019 (COVID-19): The epidemic and the challenges. Int. J. Antimicrob. Agents 2020, 55, 105924. [Google Scholar] [CrossRef]
- Acter, T.; Uddin, N.; Das, J.; Akhter, A.; Choudhury, T.R.; Kim, S. Evolution of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) as coronavirus disease 2019 (COVID-19) pandemic: A global health emergency. Sci. Total Environ. 2020, 730, 138996. [Google Scholar] [CrossRef]
- Tang, A.N.; Tong, Z.D.; Wang, H.L.; Dai, Y.X.; Li, K.F.; Liu, J.N.; Wu, W.J.; Yuan, C.; Yu, M.L.; Li, P.; et al. Detection of Novel Coronavirus by RT-PCR in Stool Specimen from Asymptomatic Child, China. Emerg. Infect. Dis. 2020, 26, 1337. [Google Scholar] [CrossRef]
- Gundy, P.M.; Gerba, C.P.; Pepper, I.L. Survival of Coronaviruses in Water and Wastewater. Food Environ. Virol. 2009, 1, 10–14. [Google Scholar] [CrossRef] [Green Version]
- Mancuso, G.; Perulli, G.D.; Lavrnić, S.; Morandi, B.; Toscano, A. Sars-cov-2 from urban to rural water environment: Occurrence, persistence, fate, and influence on agriculture irrigation. A review. Water 2021, 13, 764. [Google Scholar] [CrossRef]
- Pant, R.R.; Bishwakarma, K.; Ur, F.; Qaiser, F.U.R.; Pathak, L.; Jayaswal, G.; Sapkota, B.; Maskey, R. Imprints of COVID-19 lockdown on the surface water quality of Bagmati river basin, Nepal. J. Environ. Manag. 2021, 289, 112522. [Google Scholar] [CrossRef]
- Askitas, N.; Tatsiramos, K.; Verheyden, B. Lockdown strategies, mobility patterns, and COVID-19. J. Sustain. Tour. 2020, 2006, 00531. [Google Scholar] [CrossRef]
- Bates, A.E.; Primack, R.B.; Moraga, P.; Duarte, C.M. COVID-19 pandemic and associated lockdown as a “Global Human Confinement Experiment” to investigate biodiversity conservation. Biol. Conserv. 2020, 248, 108665. [Google Scholar] [CrossRef]
- Liu, D.; Yang, H.; Thompson, J.R.; Li, J.; Loiselle, S.; Duan, H. COVID-19 lockdown improved river water quality in China. Sci. Total. Environ. 2022, 802, 149585. [Google Scholar] [CrossRef]
- Cooke, S.J.; Twardek, W.M.; Lynch, A.J.; Cowx, I.G.; Olden, J.D.; Funge-Smith, S. A global perspective on the influence of the COVID-19 pandemic on freshwater fish biodiversity. Biol. Conserv. 2021, 253, 108932. [Google Scholar] [CrossRef]
- Pinder, A.C.; Raghavan, R.; Britton, J.R.; Cooke, S. COVID-19 and biodiversity: The paradox of cleaner rivers and elevated extinction risk to iconic fish species. Aquat. Conserv. 2020, 30, 1061–1062. [Google Scholar] [CrossRef]
- Bar, H. COVID-19 lockdown: Animal life, ecosystem, and atmospheric environment. Environ. Dev. Sustain. 2021, 23, 8161–8178. [Google Scholar] [CrossRef]
- Saadat, S.; Rawtani, D.; Hussain, C.M. Environmental perspective of COVID-19. Sci. Total. Environ. 2020, 728, 138870. [Google Scholar] [CrossRef] [PubMed]
- Tadano, Y.S.; Potgieter-Vermaak, S.; Kachba, Y.R.; Chiroli, D.M.; Casacio, L.; Santos Silva, J.C.; Godoi, R.H. Dynamic model to predict the association between air quality, COVID-19 cases, and level of lockdown. Environ. Pollut. 2021, 268, 115920. [Google Scholar] [CrossRef] [PubMed]
- Polazzo, F.; Dos Anjos, T.B.O.; Arenas-Sánchez, A.; Romo, S.; Vighi, M.; Rico, A. Effect of multiple agricultural stressors on freshwater ecosystems: The role of community structure, trophic status, and biodiversity-functioning relationships on ecosystem responses. Sci. Total. Environ. 2022, 807, 151052. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Diamond, J. China’s environment in a globalizing world. Nature 2005, 435, 1179–1186. [Google Scholar] [CrossRef]
- Zhang, H.; Jarić, I.; Roberts, D.L.; He, Y.; Du, H.; Wu, J.; Wei, Q. Extinction of one of the world’s largest freshwater fishes: Lessons for conserving the endangered Yangtze fauna. Sci. Total Environ. 2020, 710, 136242. [Google Scholar] [CrossRef]
- Xiong, F.; Olden, J.D.; Lu, Y.; Liu, H.; Qu, X.; Xia, W.; Chen, Y. Riparian land use and in-channel stressors drive fish community structure in the Yangtze River. Landsc. Ecol. 2021, 36, 3079–3095. [Google Scholar] [CrossRef]
- Zhang, M.; Tan, S.; Zhang, Y.; He, J.; Ni, Q. Does land transfer promote the development of new-type urbanization? New evidence from urban agglomerations in the middle reaches of the Yangtze River. Ecol. Indic. 2022, 36, 108705. [Google Scholar] [CrossRef]
- Yang, H.L.; Wu, J.M.; Du, H.; Zhang, H.; Li, J.Y.; Wei, Q.W. Quantifying the Colonization of Environmental Microbes in the Fish Gut: A Case Study of Wild Fish Populations in the Yangtze River. Front. Microbiol. 2021, 12, 828409. [Google Scholar] [CrossRef]
- Dong, C.Y.; Li, J.Y.; Zhang, H.; Yang, H.L.; Xiang, H.; Wu, J.M.; Jiang, H.B. Estimation of Fish Resources in Middle Reaches of Yangtze River. Chin. J. Zool. 2021, 56, 73–79. [Google Scholar] [CrossRef]
- Yang, G.S.; Zhu, C.Q.; Jiang, Z.G. Yangtze Conservation and Development Report 2011; Changjiang Press: Wuhan, China, 2011. [Google Scholar]
- Yang, Y.T.; Yu, D.; Gao, X.; Liu, H.Z. Mechanism of fish community assembly in middle reaches of the Yangtze River. Acta Hydrobiol. Sin. 2020, 44, 1–10. [Google Scholar]
- Wu, T.H.; Liu, J.S.; Deng, J.M. Community structure of phytoplankton and bioassessment of water quality in a large water-carrying lake, Lake Hongze. J. Lake Sci. 2019, 31, 440–448. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.R. Determination of organic pollutants in water and wastewater monitoring. J. Environ. Res. Dev. 2020, 32, 191. [Google Scholar]
- Yang, X.; Ma, J.S.; Zhang, H.; Zhou, Q. Community structure and the water quality during different hydrological periods in Poyang Lake. Acta. Hydrobiol. Sin. 2021, 45, 1093–1103. [Google Scholar] [CrossRef]
- Ding, R.H. The Fishes of Sichuan; Sichuan Publishing House of Science and Technology: Chengdu, China, 1994. [Google Scholar]
- Tong, Y.; Zhang, W.; Wang, X.; Couture, R.M.; Larssen, T.; Zhao, Y.; Lin, Y. Decline in Chinese lake phosphorus concentration accompanied by shift in sources since 2006. Nat. Geosci. 2017, 10, 507–511. [Google Scholar] [CrossRef]
- Yang, J.; Yan, D.; Yang, Q.; Gong, S.; Shi, Z.; Qiu, Q.; Hu, M. Fish species composition, distribution and community structure in the Fuhe River Basin, Jiangxi Province, China. Glob. Ecol. Conserv. 2021, 27, e01559. [Google Scholar] [CrossRef]
- Gao, S.B.; Chi, S.Y.; Li, S.X.; Zheng, J.X.; Dong, F.Y. Fish resource investigation and species diversity analysis of Nanxi River in Zhejiang Province. J. Hydroecol. 2017, 38, 72–81. [Google Scholar]
- Warwick, R.M. A new method for detecting pollution effects on marine macrobenthic communities. Mar. Biol. 1986, 92, 557–562. [Google Scholar] [CrossRef]
- Piperac, M.S.; Miloevi, D.; Simi, V. The application of the abundance/biomass comparison method on riverine fish assemblages: Limits of use in lotic systems. Biol. Nyssana 2015, 6, 25–32. [Google Scholar]
- Bu, H.; Liu, W.; Song, X.; Zhang, Q. Quantitative impacts of population on river water quality in the Jinshui River basin of the South Qinling Mts. China. Environ. Earth Sci. 2016, 75, 292. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, Y.; Arhonditsis, G.B.; Gao, J.; Chen, Q.; Wu, N.; Shi, W. How successful are the restoration efforts of China’s lakes and reservoirs? Environ. Int. 2019, 123, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Sada, R. Hanumante River: Emerging uses, competition and implications. Arab. J. Sci. Eng. 2012, 1, 17–24. [Google Scholar] [CrossRef]
- Gomes-Silva, G.; Cyubahiro, E.; Wronski, T.; Riesch, R.; Apio, A.; Plath, M. Water pollution affects fish community structure and alters evolutionary trajectories of invasive guppies (Poecilia reticulata). Sci. Total Environ. 2020, 730, 138912. [Google Scholar] [CrossRef] [PubMed]
- Kahlon, S.K.; Sharma, G.; Julka, J.M.; Kumar, A.; Sharma, S.; Stadler, F.J. Impact of heavy metals and nanoparticles on aquatic biota. Environ. Chem. Lett. 2018, 16, 919–946. [Google Scholar] [CrossRef]
- Braga, F.; Scarpa, G.M.; Brando, V.E.; Manfe, G.; Zaggia, L. COVID-19 lockdown measures reveal human impact on water transparency in the Venice Lagoon. Sci. Total Environ. 2020, 736, 139612. [Google Scholar] [CrossRef]
- Muduli, P.R.; Kumar, A.; Kanuri, V.V.; Mishra, D.R.; Mishra, D.R.; Sudhakar, A. Water quality assessment of the Ganges River during COVID-19 lockdown. Int. J. Environ. Sci. Technol. 2021, 18, 1645–1652. [Google Scholar] [CrossRef]
- Cooper, M.J.; Martin, R.V.; Hammer, M.S.; Levelt, P.F.; Veefkind, P.; Lamsal, L.N. Global fine-scale changes in ambient NO2 during COVID-19 lockdowns. Nature 2022, 601, 380–387. [Google Scholar] [CrossRef]
- Xiong, W.; Huang, X.; Chen, Y.; Fu, R.; Du, X.; Chen, X.; Zhan, A. Zooplankton biodiversity monitoring in polluted freshwater ecosystems: A technical review. Environ. Sci. Ecotechnol. 2020, 1, 100008. [Google Scholar] [CrossRef]
- Sommer, U.; Adrian, R.; De, L.; Domis, S.; Elser, J.J.; Gaedke, U.; Winder, M. Beyond the Plankton Ecology Group (PEG) Model: Mechanisms Driving Plankton Succession. Annu. Rev. Ecol. Evol. Sci. 2012, 43, 429–448. [Google Scholar] [CrossRef]
- Pan, J.Z.; Xiong, F.; Li, W.C.; Li, Y.X. Spatial-temporal dynamic changes of the water transparency and their influencing factors in Lake Fuxian, Yunnan Province. J. Lake Sci. 2008, 20, 681–686. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Peng, S.T.; Qin, X.B.; Shi, H.H.; Ding, D.W. Phytoplankton assemblages and their relation to environmental factors by Multivariate Statistic Analysis in Bohai Bay. Chin. J. Environ. Sci. 2013, 34, 864–873. [Google Scholar] [CrossRef]
- Liu, Y.; Lv, J.P.; Liu, Q.; Xie, S.L.; Feng, J. Relationship between the growth of phytoplankton and the distribution of nutrients in the fenhe water storage area of taiyuan. Acta Ecol. Sin. 2018, 38, 991–1002. [Google Scholar] [CrossRef]
- Xu, H.; Chen, J.; Zhu, G.W.; Qin, B.Q.; Zhang, Y.L. Effect of concentrations of phosphorus and nitrogen on the dominance of cyanobacteria. J. Lake Sci. 2019, 31, 1239–1247. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, G.; Viehweg, M. Recent developments of the phytoplankton and its activity in the Lower Rhine. SIL Proc. 2017, 22, 1992–2010. [Google Scholar] [CrossRef]
- Hu, J.; Chi, S.Y.; Hu, J.X. Partial Canonical Correspondence Analysis of Phytoplankton Community and Environmental Factors in Nansihu Lake. Environ. Sci. Technol. 2020, 43, 33–39. [Google Scholar]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.L.; Wu, Z.Q.; Li, J.H. Fish fauna, biogeography and conservation of freshwater fish in Poyang Lake Basin, China. Environ. Biol. Fish. 2013, 96, 1229–1243. [Google Scholar] [CrossRef]
- Fulton, E.A.; Smith, A.D.; Smith, D.C.; van Putten, I.E. Human behaviour: The key source of uncertainty in fisheries management. Fish. Fish. 2011, 12, 2–17. [Google Scholar] [CrossRef]
- Willby, N.J.; Eaton, J.W. Backwater habitats and their role in nature conservation on navigable waterways. Hydrobiologia 1996, 340, 333–338. [Google Scholar] [CrossRef]
- Chen, K.; Olden, J.D. Threshold responses of riverine fish communities to land use conversion across regions of the world. Glob. Chang. Biol. 2020, 26, 4952–4965. [Google Scholar] [CrossRef]
- Connell, J.H. Diversity in coral reefs and tropical rainforests. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.F.; Li, X.H.; Yang, J.P.; Lek, S.; Shuai, F.M.; Li, J. Effect of Pearl River closed fishing on Megalobrama hoffmanni recruitment stock. J. Fish. 2014, 38, 503–509. [Google Scholar]
- Heino, J.; Soininen, J.; Alahuhta, J.; Lappalainen, J.; Virtanen, R. A comparative analysis of metacommunity types in the freshwater realm. Ecol. Evol. 2015, 5, 1525–1537. [Google Scholar] [CrossRef] [PubMed]
- Karr, J.R. Seasonality, Resource Availability, and Community Diversity in Tropical Bird Communities. Am. Nat. 1976, 110, 973–994. [Google Scholar] [CrossRef]
- Cheng, J.H.; Yan, L.P.; Lin, L.S.; Yu, L.F.; Liu, Z.F.; Zhou, Y.D. Analyses on the fishery ecological effect of summer close season in the East China Sea region. J. Fish. Sci. 1999, 6, 81–85. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, D.; Yang, H.; Zhang, H.; Du, H.; Yang, J.; Wei, Q. COVID-19 Lockdowns Promoted Recovery of the Yangtze River’s Aquatic Ecosystem. Water 2022, 14, 3622. https://doi.org/10.3390/w14223622
Fang D, Yang H, Zhang H, Du H, Yang J, Wei Q. COVID-19 Lockdowns Promoted Recovery of the Yangtze River’s Aquatic Ecosystem. Water. 2022; 14(22):3622. https://doi.org/10.3390/w14223622
Chicago/Turabian StyleFang, Dongdong, Haile Yang, Hui Zhang, Hao Du, Junlin Yang, and Qiwei Wei. 2022. "COVID-19 Lockdowns Promoted Recovery of the Yangtze River’s Aquatic Ecosystem" Water 14, no. 22: 3622. https://doi.org/10.3390/w14223622
APA StyleFang, D., Yang, H., Zhang, H., Du, H., Yang, J., & Wei, Q. (2022). COVID-19 Lockdowns Promoted Recovery of the Yangtze River’s Aquatic Ecosystem. Water, 14(22), 3622. https://doi.org/10.3390/w14223622