
Exploring Key Determinants of the Periphytic Diatom Community in a Southern Brazilian Micro-Watershed

 ,
,  , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
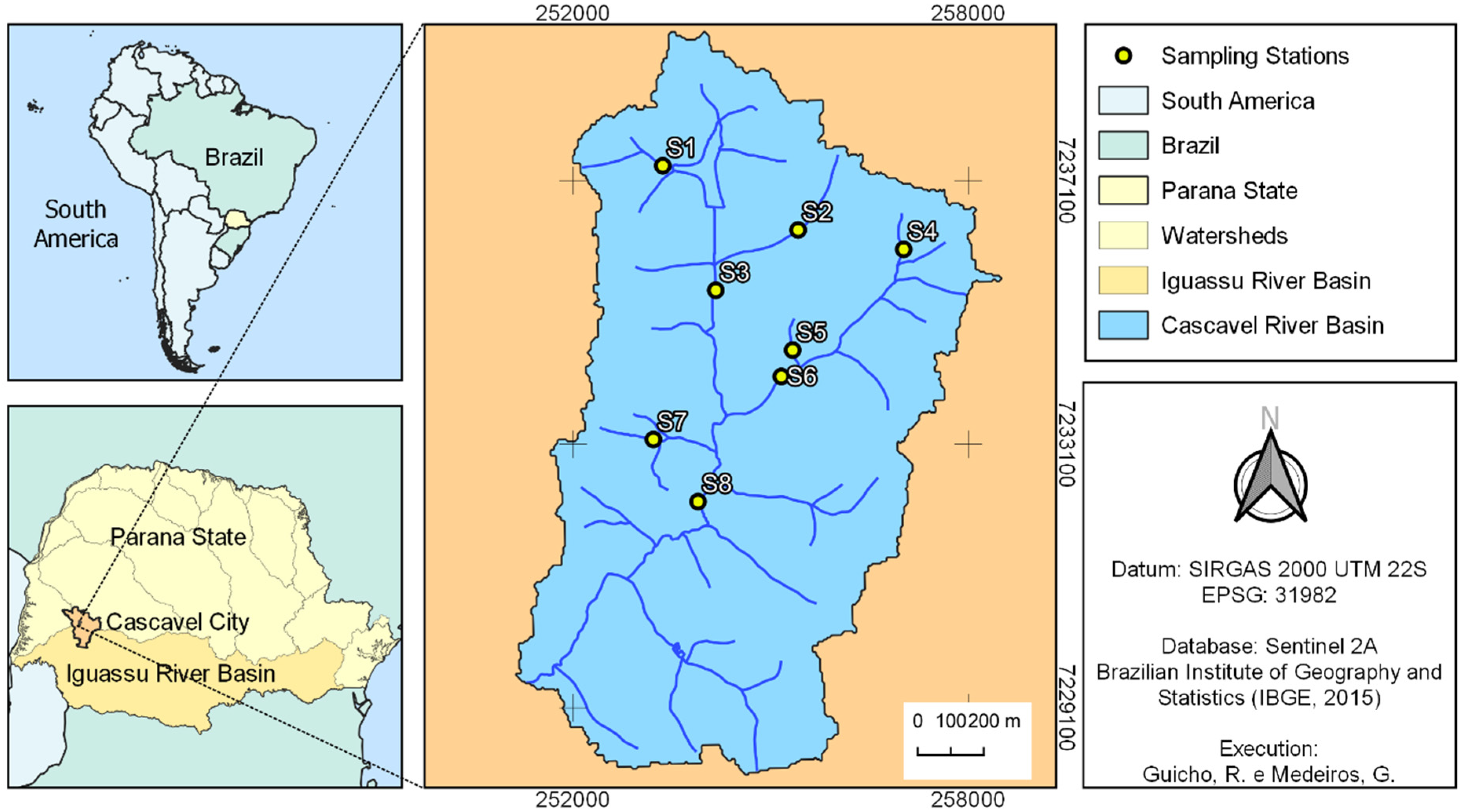
2.1. Study Area
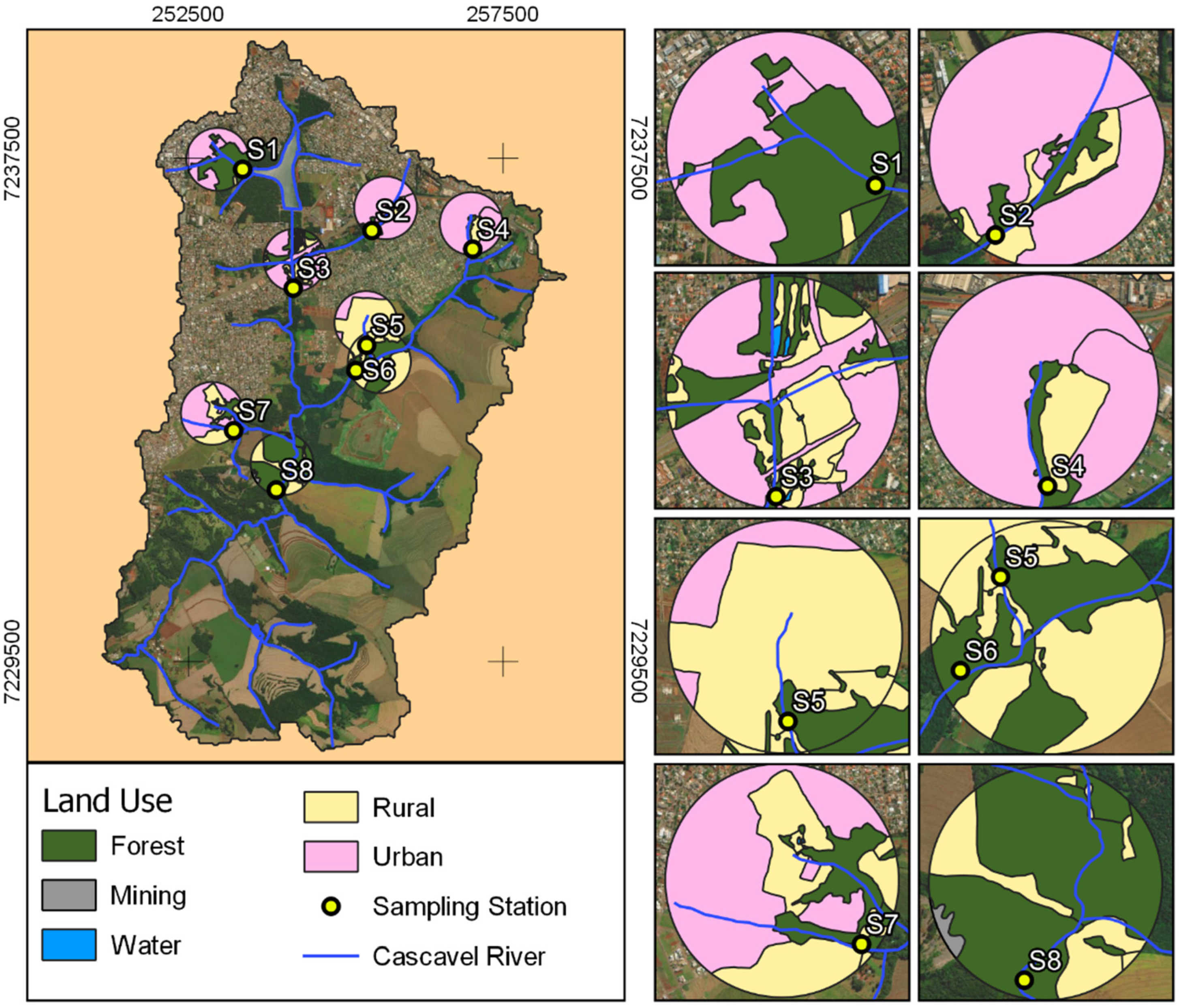
2.2. Sampling Design
2.3. Landscape and Climate Variables
2.4. Local Environmental Variables
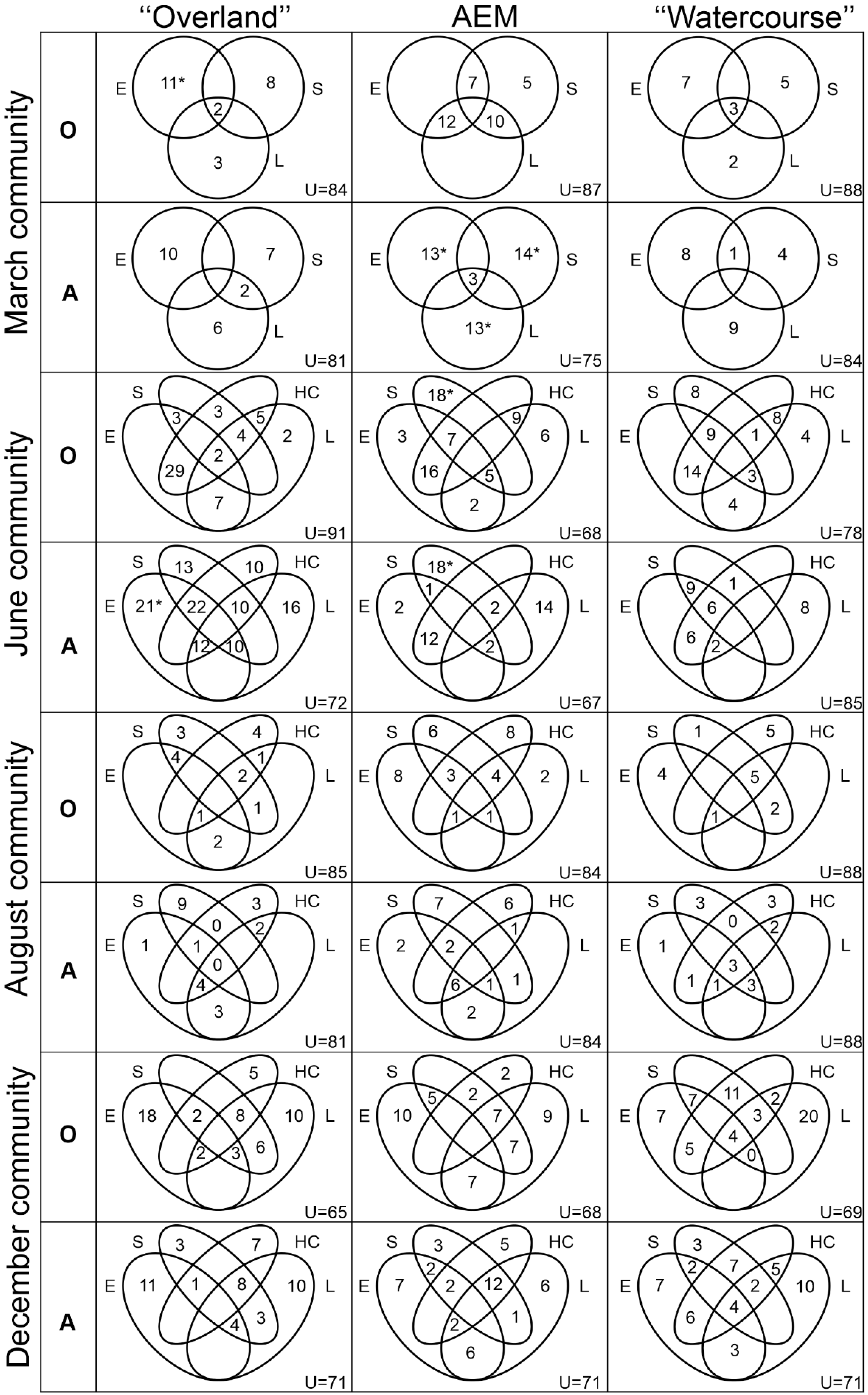
2.5. Dispersal Route Indicators
- (1)
- To test “overland”, we considered the Euclidean distances resulting from the Principal Coordinate of Neighbor Matrices (PCNM, [47]), an ordination matrix generated using the coordinates of the sampling stations, creating a spatial connection network with links at all evaluated sites without directionality (i.e., the sampling stations were connected by two directions, upstream–downstream and downstream–upstream).
- (2)
- To test asymmetric eigenvector mapping “AEM”, the latitude and longitude of the sites were used to relate the points, considering whether the sites had connectivity or not [48].
- (3)
- To test “watercourse”, the distance matrix was generated by drawing a dendritic system over a real river map with Quantum GIS software. Thus, we modeled a spatial network with directional links over the watercourse (considering that the sites were connected only as a function of the river water flow).
2.6. Periphytic Community
2.7. Data Analysis
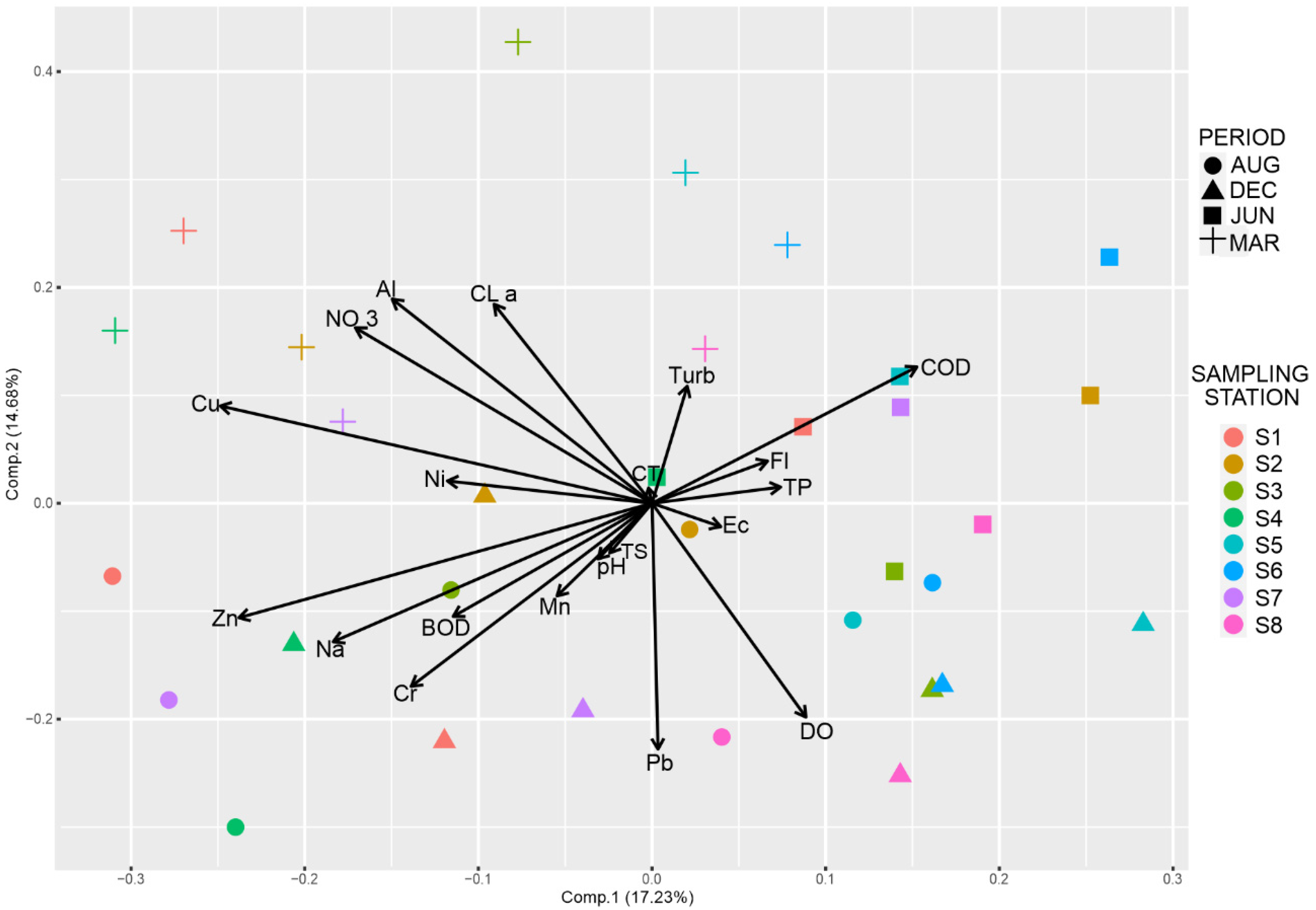
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Station | S1 | S2 | S3 | S4 | S5 | S6 | S7 | S8 | R² | F | p-Value | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Achnanthidium catenatum (J. Bily & Marvan) Lange-Bertalot | mean | - | - | - | 7.5% | 0.9% | - | - | - | 0.220 | 0.966 | 0.478 |
| standard deviation | - | - | - | 14.9% | 1.8% | - | - | - | ||||
| Achnanthidium exiguum (Grunow) D.B.Czarnecki | mean | - | - | - | - | - | - | - | 1.1% | 0,.26 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | - | - | - | 2.2% | ||||
| Achnanthidium macrocephalum (Hustedt) Round & Bukhtiyarova | mean | 1.1% | - | 3.2% | 2.7% | - | - | - | 2.8% | 0.259 | 1.198 | 0.341 |
| standard deviation | 2.2% | - | 3.2% | 5.3% | - | - | - | 3.3% | ||||
| Achnanthidium minutissimum (Kützing) Czarnecki | mean | 5.8% | 3.6% | 3.0% | 5.5% | - | 7.8% | 5.0% | 9.1% | 0.172 | 0.714 | 0.661 |
| standard deviation | 8.7% | 4.4% | 3.7% | 7.1% | - | 9.1% | 6.0% | 9.0% | ||||
| Achnanthidium modestiforme (Lange-Bertalot) Van de Vijver | mean | 1.9% | 1.4% | - | 11.1% | - | 1.4% | - | 2.2% | 0.477 | 3.130 | 0.017 |
| standard deviation | 2.6% | 2.9% | - | 9.9% | - | 2.8% | - | 4.3% | ||||
| Achnanthidium tropicocatenatum G.C.Marquardt, C.E.Wetzel & Ector | mean | 0.4% | 0.7% | 3.6% | - | - | - | - | - | 0.274 | 1.291 | 0.297 |
| standard deviation | 0.7% | 1.5% | 6.0% | - | - | - | - | - | ||||
| Actinella hermes-moreirae Ruwer, Ludwig & Rodrigues | mean | - | 0.2% | - | - | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | 0.5% | - | - | - | - | - | - | ||||
| Aulacoseira ambigua (Grunow) Simonsen | mean | - | 0.5% | 4.4% | - | 0.3% | - | - | 2.8% | 0.379 | 2.089 | 0.085 |
| standard deviation | - | 1.0% | 3.2% | - | 0.6% | - | - | 5.6% | ||||
| Aulacoseira granulata (Ehrenberg) Simonsen | mean | - | 2.0% | - | - | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | 3.9% | - | - | - | - | - | - | ||||
| Aulacoseira pusilla (Meister) Tuji & Houki | mean | 2.1% | - | 1.3% | - | - | - | - | - | 0.230 | 1.026 | 0.439 |
| standard deviation | 4.3% | - | 1.6% | - | - | - | - | - | ||||
| Aulacoseira tenella (Nygaard) Simonsen | mean | - | - | 1.4% | - | 0.8% | - | - | 0.8% | 0.179 | 0.747 | 0.635 |
| standard deviation | - | - | 2.8% | - | 1.6% | - | - | 1.5% | ||||
| Brachysira brebissonii Ross | mean | - | 0.2% | - | - | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | 0.5% | - | - | - | - | - | - | ||||
| Brachysira microcephala (Grunow) Compère | mean | - | - | 1.4% | - | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | 2.9% | - | - | - | - | - | ||||
| Caloneis hyalina Hustedt | mean | - | - | - | 1.0% | 0.9% | - | - | - | 0.200 | 0.858 | 0.552 |
| standard deviation | - | - | - | 2.1% | 1.8% | - | - | - | ||||
| Caloneis westii (Smith) Hendey | mean | - | - | - | - | - | - | 0.7% | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | - | - | 1.4% | - | ||||
| Craticula riparia (Hustedt) Lange-Bertalot | mean | - | 0.2% | - | - | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | 0.5% | - | - | - | - | - | - | ||||
| Craticula submolesta (Hustedt) Lange-Bertalot | mean | - | - | - | 2.6% | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | 5.2% | - | - | - | - | ||||
| Cymbopleura naviculiformis (Auerswald) Krammer | mean | - | - | 0.7% | - | - | - | - | - | 0.465 | 2.979 | 0.021 |
| standard deviation | - | - | 0.9% | - | - | - | - | - | ||||
| Discostella stelligera (Cleve & Grunow) Houk & Klee | mean | 3.5% | 0.2% | 13.7% | 1.9% | - | 1.0% | 1.6% | 7.2% | 0.356 | 1.897 | 0.115 |
| standard deviation | 7.0% | 0.5% | 17.1% | 3.7% | - | 1.9% | 3.1% | 0.8% | ||||
| Encyonema minutiforme Krammer | mean | - | - | - | - | - | 2.1% | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | - | 4.2% | - | - | ||||
| Encyonema neomesianum Krammer | mean | - | - | 0.5% | - | - | 0.7% | - | - | 0.202 | 0.867 | 0.546 |
| standard deviation | - | - | 1.0% | - | - | 1.4% | - | - | ||||
| Encyonema perpusillum (Cleve-Euler) Mann | mean | 0.4% | - | - | - | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | 0.7% | - | - | - | - | - | - | - | ||||
| Encyonema silesiacum (Bleisch) Mann | mean | - | 0.2% | - | 0.5% | - | 0.6% | - | 2.0% | 0.259 | 1.199 | 0.341 |
| standard deviation | - | 0.5% | - | 1.0% | - | 1.1% | - | 3.2% | ||||
| Eunotia bilunaris (Ehrenberg) Schaarschmidt | mean | 3.4% | - | 7.0% | - | 0.3% | 5.8% | 4.2% | 0.3% | 0.330 | 1.686 | 0.160 |
| standard deviation | 6.8% | - | 4.0% | - | 0.6% | 7.2% | 6.4% | 0.5% | ||||
| Eunotia botulitropica Wetzel & Costa | mean | 11.3% | 2.0% | 4.9% | 0.5% | 11.9% | 6.3% | 16.8% | 0.5% | 0.231 | 1.029 | 0.437 |
| standard deviation | 16.6% | 2.9% | 7.3% | 1.1% | 9.6% | 11.4% | 23.6% | 1,1% | ||||
| Eunotia georgii Metzeltin & Lange-Bertalot | mean | - | - | - | - | 1.4% | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | 2.7% | - | - | - | ||||
| Eunotia intricans Lange-Bertalot & Metzeltin | mean | - | - | - | - | - | 0.4% | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | - | 0.9% | - | - | ||||
| Eunotia kruegeri Lange-Bertalot | mean | 0.7% | 0.8% | - | 1.6% | 2,4% | - | - | - | 0.168 | 0.693 | 0.677 |
| standard deviation | 1.4% | 1.5% | - | 3.2% | 4.8% | - | - | - | ||||
| Eunotia meridiana Metzeltin & Lange-Bertalot | mean | 3.5% | 5.6% | 6.8% | 2.0% | 6.1% | 3.5% | 0.8% | 2.2% | 0.135 | 0.535 | 0.799 |
| standard deviation | 4.2% | 4.9% | 9.2% | 2.3% | 10.4% | 4.2% | 1.6% | 4.3% | ||||
| Eunotia rhomboidea Hustedt | mean | 7.0% | 5.6% | 2.8% | 1.9% | 15.8% | 3.1% | 5.8% | 4.8% | 0.333 | 1.716 | 0.153 |
| standard deviation | 5.2% | 4.4% | 4.1% | 3.7% | 14.7% | 3.6% | 3.9% | 6.0% | ||||
| Eunotia subarcuatoides Alles, Nörpel & Lange-Bertalot | mean | - | - | - | - | - | 0.7% | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | - | 1.4% | - | - | ||||
| Eunotia tropico-arcus Metzeltin & Lange-Bertalot | mean | - | - | - | - | - | 0.3% | 0.2% | - | 0.201 | 0.860 | 0.551 |
| standard deviation | - | - | - | - | - | 0.6% | 0.5% | - | ||||
| Eunotia veneris (Kützing) De Toni | mean | 19.6% | - | 1.6% | - | 1.4% | - | 0.2% | 0.7% | 0.239 | 1.075 | 0.409 |
| standard deviation | 36.7% | - | 3.1% | - | 2.7% | - | 0.5% | 1.4% | ||||
| Eunotia yberai Frenguelli | mean | - | - | - | 2.0% | - | - | - | - | 0.465 | 2.983 | 0.021 |
| standard deviation | - | - | - | 2.3% | - | - | - | - | ||||
| Fragilaria gracilis Østrup | mean | - | - | 2.4% | - | 0.3% | 0.9% | - | - | 0.362 | 1.943 | 0.107 |
| standard deviation | - | - | 2.9% | - | 0.6% | 1.8% | - | - | ||||
| Fragilaria pectinalis (Müller) Lyngbye | mean | - | 1.2% | 1.3% | - | - | 2.7% | 0.8% | 1.0% | 0.152 | 0.614 | 0.739 |
| standard deviation | - | 2.4% | 1.5% | - | - | 5.4% | 1.6% | 2.0% | ||||
| Fragilaria tenera var. tenera (Smith) Lange-Bertalot | mean | - | - | - | - | - | 2.0% | - | 1.4% | 0.202 | 0.866 | 0.547 |
| standard deviation | - | - | - | - | - | 3.9% | - | 2.8% | ||||
| Fragilaria sp. | mean | - | - | 0.5% | - | - | 3.1% | - | - | 0.218 | 0.957 | 0.483 |
| standard deviation | - | - | 1.0% | - | - | 6.3% | - | - | ||||
| Fragilariforma javanica (Hustedt) Wetzel, Morales & Ector | mean | - | - | - | - | 1.0% | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | 2.0% | - | - | - | ||||
| Frustulia acidophilissima Wydrzycka & Lange-Bertalot | mean | 1.4% | - | - | 0.5% | 1.0% | 1.5% | 1.7% | 0.5% | 0.107 | 0.410 | 0.887 |
| standard deviation | 2.7% | - | - | 1.1% | 2.0% | 2.9% | 3.4% | 1.1% | ||||
| Frustulia crassinervia (Brébisson ex W.Smith) Lange-Bertalot & Krammer | mean | 1.2% | - | - | 1.5% | 5.4% | - | - | 0.7% | 0.331 | 1.698 | 0.157 |
| standard deviation | 0.9% | - | - | 3.1% | 7.2% | - | - | 1.4% | ||||
| Frustulia guayanensis Metzeltin & Lange Bertalot | mean | - | 0.8% | 0.4% | - | 0.7% | 1.7% | - | - | 0.171 | 0.706 | 0.667 |
| standard deviation | - | 1.5% | 0.8% | - | 1.4% | 3.4% | - | - | ||||
| Frustulia neomundana Lange-Bertalot & Rumrich | mean | - | - | - | - | - | - | 0.8% | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | - | - | 1.6% | - | ||||
| Frustulia saxonica Rabenhorst | mean | - | - | - | - | - | 0.5% | - | 0.5% | 0.200 | 0.858 | 0.552 |
| standard deviation | - | - | - | - | - | 1.0% | - | 1.1% | ||||
| Frustulia undosa Metzeltin & Lange-Bertalot | mean | 0.2% | - | - | - | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | 0.4% | - | - | - | - | - | - | - | ||||
| Frustulia vulgaris (Thwaites) De Toni | mean | - | - | - | - | 0.5% | 1.0% | - | - | 0.206 | 0.890 | 0.530 |
| standard deviation | - | - | - | - | 1.0% | 2.1% | - | - | ||||
| Frustulia weinholdii Hustedt | mean | 0.4% | - | - | - | - | - | - | 0.9% | 0.368 | 1.993 | 0.098 |
| standard deviation | 0.7% | - | - | - | - | - | - | 1.1% | ||||
| Geissleria punctifera (Hustedt) Metzeltin, Lange-Bertalot e García-Rodríguez | mean | - | 0.2% | - | - | 0.5% | - | - | 1.1% | 0.195 | 0.830 | 0.573 |
| standard deviation | - | 0.5% | - | - | 1.0% | - | - | 2.3% | ||||
| Gogorevia exilis(Kütz.) Kulikovskiy and Kociolek | mean | - | - | - | - | - | - | - | 1.4% | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | - | - | - | 2.8% | ||||
| Gomphonema angustatum (Kützing) Rabenhorst | mean | - | - | - | - | - | - | - | 1.6% | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | - | - | - | 3.3% | ||||
| Gomphonema exilissimum (Grunow) Lange-Bertalot & Reichardt | mean | - | - | 0.6% | - | - | 6.3% | - | 1.5% | 0.209 | 0.904 | 0.520 |
| standard deviation | - | - | 1.1% | - | - | 12.5% | - | 3.1% | ||||
| Gomphonema guaraniarum Metzeltin & Lange-Bertalot | mean | - | - | - | - | - | 1.1% | - | 0.8% | 0.201 | 0.864 | 0.548 |
| standard deviation | - | - | - | - | - | 2.1% | - | 1.5% | ||||
| Gomphonema graciledictum Reichardt | mean | - | - | 0.8% | - | - | - | - | 0.3% | 0.207 | 0.894 | 0.527 |
| standard deviation | - | - | 1.6% | - | - | - | - | 0.7% | ||||
| Gomphonema lagenula Kützing | mean | 3.3% | 7.0% | 3.8% | 1.0% | - | 2.7% | 0.8% | 4.5% | 0.202 | 0.869 | 0.544 |
| standard deviation | 4.1% | 7.4% | 6.7% | 2.1% | - | 4.2% | 1.6% | 7.3% | ||||
| Gomphonema naviculoides Smith | mean | 0.4% | - | 0.5% | - | - | 1.3% | - | - | 0.195 | 0.829 | 0.574 |
| standard deviation | 0.7% | - | 1.0% | - | - | 2.7% | - | - | ||||
| Gomphonema obtusatum (Kützing) Grunow | mean | - | - | 0.4% | 1.1% | - | - | - | - | 0.209 | 0.908 | 0.517 |
| standard deviation | - | - | 0.8% | 2.1% | - | - | - | - | ||||
| Gomphonema parvulum (Kützing) Kützing | mean | 5.9% | 13.0% | 19.7% | 2.8% | 9.6% | 22.7% | 3.1% | 10.3% | 0.236 | 1.060 | 0.418 |
| standard deviation | 5.8% | 9.4% | 33.2% | 3.2% | 8.0% | 11.7% | 6.3% | 11.4% | ||||
| Gomphonema pumilum (Grunow) Reichardt & Lange-Bertalot | mean | - | 2.2% | 0.7% | - | - | 0.9% | 2.7% | - | 0.208 | 0.900 | 0.522 |
| standard deviation | - | 2.9% | 0.8% | - | - | 1.8% | 5.4% | - | ||||
| Halamphora montana (Krasske) Levkov | mean | - | 0.7% | - | 1.0% | - | 0.3% | 3.9% | 0.7% | 0.185 | 0.781 | 0.610 |
| standard deviation | - | 1.5% | - | 2.1% | - | 0.6% | 7.8% | 1.4% | ||||
| Hantzschia amphioxys (Ehrenberg) Grunow | mean | - | 0.2% | - | - | - | 0.3% | - | - | 0.200 | 0.859 | 0.552 |
| standard deviation | - | 0.5% | - | - | - | 0.6% | - | - | ||||
| Humidophila contenta (Grunow) Lowe et al. | mean | 0.4% | 4.8% | - | 1.5% | - | - | 7.4% | - | 0.309 | 1.533 | 0.204 |
| standard deviation | 0.9% | 8.9% | - | 3.1% | - | - | 8.5% | - | ||||
| Humidophila subtropica (Metzeltin, Lange-Bertalot & García-Rodríguez) Lowe et al. | mean | - | 1.5% | - | - | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | 2.9% | - | - | - | - | - | - | ||||
| Iconella tenuissima (Hustedt) D.Kapustin & Kulikovskiy | mean | - | - | - | 1.6% | - | 0.5% | - | 0.3% | 0.342 | 1.779 | 0.138 |
| standard deviation | - | - | - | 2.0% | - | 1.0% | - | 0.7% | ||||
| Luticola acidoclinata Lange-Bertalot | mean | - | 0.2% | - | - | 2.8% | 1.9% | - | 0.9% | 0.262 | 1.214 | 0.333 |
| standard deviation | - | 0.5% | - | - | 3.8% | 3.9% | - | 1.1% | ||||
| Luticola goeppertiana (Bleisch) Mann | mean | - | - | 0.4% | - | 1.5% | - | - | 2.5% | 0.194 | 0.826 | 0.576 |
| standard deviation | - | - | 0.8% | - | 2.9% | - | - | 5.0% | ||||
| Luticola hustedtii Levkov, Metzeltin & Pavlov | mean | 0.2% | - | - | - | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | 0.4% | - | - | - | - | - | - | - | ||||
| Luticola permuticoides Metzeltin e Lange-Bertalot | mean | - | 0.7% | - | - | - | 1.0% | - | - | 0.202 | 0.866 | 0.546 |
| standard deviation | - | 1.4% | - | - | - | 1.9% | - | - | ||||
| Navicula angusta Grunow | mean | - | 0.2% | - | - | 1.1% | - | - | - | 0.215 | 0.938 | 0.496 |
| standard deviation | - | 0.5% | - | - | 2.1% | - | - | - | ||||
| Navicula cryptocephala Kützing | mean | 1.5% | 7.5% | 2.3% | - | - | 0.7% | - | 4.3% | 0.475 | 3.102 | 0.018 |
| standard deviation | 1.5% | 6.8% | 2.8% | - | - | 1.4% | - | 3.7% | ||||
| Navicula cryptotenella Lange-Bertalot | mean | 1.3% | 0.2% | 1.5% | - | 10.0% | - | 1.2% | - | 0.472 | 3.063 | 0.019 |
| standard deviation | 2.0% | 0.5% | 2.2% | - | 10.4% | - | 1.5% | - | ||||
| Navicula eichhorniaephila Manguin ex Kociolek & Reviers | mean | - | - | - | 0.5% | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | 1.1% | - | - | - | - | ||||
| Navicula leptostriata E.G.Jørgensen | mean | - | - | 0.4% | - | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | 0.8% | - | - | - | - | - | ||||
| Navicula radiosa Kützing | mean | - | 2.0% | 0.3% | - | 0.5% | 0.6% | - | - | 0.186 | 0.783 | 0.608 |
| standard deviation | - | 4.1% | 0.7% | - | 1.0% | 1.3% | - | - | ||||
| Navicula rostellata Kützing | mean | - | - | - | - | - | 1.1% | - | 0.3% | 0.215 | 0.939 | 0.496 |
| standard deviation | - | - | - | - | - | 2.3% | - | 0.5% | ||||
| Navicula salinicola Hustedt | mean | 0.2% | 3.6% | - | - | - | - | - | - | 0.431 | 2.596 | 0.038 |
| standard deviation | 0.4% | 4.4% | - | - | - | - | - | - | ||||
| Navicula symmetrica Patrick | mean | - | 1.7% | - | - | - | - | - | 1.4% | 0.201 | 0.860 | 0.551 |
| standard deviation | - | 3.4% | - | - | - | - | - | 2.8% | ||||
| Neidium essequiboanum Metzeltin & Krammer | mean | - | - | - | - | - | 0.5% | - | 0.8% | 0.203 | 0.876 | 0.540 |
| standard deviation | - | - | - | - | - | 1.0% | - | 1.7% | ||||
| Neidium gracile f. aequale Hustedt | mean | 0.4% | - | - | - | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | 0.7% | - | - | - | - | - | - | - | ||||
| Neidium iridis (Ehrenberg) Cleve | mean | - | - | - | - | 0.5% | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | 1.0% | - | - | - | ||||
| Neidium tenuissimum Hustedt | mean | 0.7% | - | - | - | - | - | 1.4% | - | 0.205 | 0.886 | 0.533 |
| standard deviation | 1.4% | - | - | - | - | - | 2.7% | - | ||||
| Nitzschia amphibia Grunow | mean | 0.7% | - | - | - | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | 1.4% | - | - | - | - | - | - | - | ||||
| Nitzschia brevissima Grunow | mean | - | - | - | 0.5% | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | 1.0% | - | - | - | - | ||||
| Nitzschia palea (Kützing) Smith | mean | 1.6% | 4.5% | 0.9% | 2.1% | 1.0% | 0.6% | - | 1.3% | 0.268 | 1.255 | 0.313 |
| standard deviation | 2.6% | 3.7% | 1.0% | 4.1% | 2.0% | 1.1% | - | 2.1% | ||||
| Nitzschia perminuta Grunow in Van Heurck | mean | 0.4% | - | - | - | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | 0.7% | - | - | - | - | - | - | - | ||||
| Nupela pardinhoensis Bes, Torgan & Ector in Bes et al. | mean | 2.4% | 2.6% | - | 3.9% | 2.4% | 2.1% | 8.3% | 2.1% | 0.223 | 0.982 | 0.467 |
| standard deviation | 2.8% | 3.6% | - | 3.5% | 4.8% | 4.2% | 10.1% | 3.1% | ||||
| Nupela praecipuoides Tremarin & Ludwig | mean | 1.1% | - | - | 2.0% | 4.3% | 1.9% | 5.4% | 7.3% | 0.284 | 1.363 | 0.266 |
| standard deviation | 2.1% | - | - | 2.9% | 6.5% | 3.9% | 4.5% | 8.7% | ||||
| Nupela semifasciata Amaral, T.Ludwig et Bueno | mean | 2.7% | 2.7% | - | 2.4% | 1.5% | 2.6% | 8.3% | 2.4% | 0.141 | 0.564 | 0.777 |
| standard deviation | 5.4% | 5.4% | - | 3.5% | 2.9% | 3.9% | 14.4% | 4.9% | ||||
| Nupela wellneri (Lange-Bertalot) Lange-Bertalot | mean | - | - | - | - | - | - | 1.1% | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | - | - | 2.3% | - | ||||
| Orthoseira roeseana (Rabenhorst) Pfitzer | mean | - | - | - | - | 2.0% | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | 4.1% | - | - | - | ||||
| Pinnularia brauniana (Grunow) Studnicka | mean | - | - | 0.4% | - | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | 0.8% | - | - | - | - | - | ||||
| Pinnularia divergens W.Smith | mean | 0.4% | - | 1.4% | 0.5% | - | - | - | - | 0.195 | 0.832 | 0.572 |
| standard deviation | 0.7% | - | 2.9% | 1.1% | - | - | - | - | ||||
| Pinnularia graciloides var. latecapitata Metzeltin & Krammer | mean | - | - | - | - | - | 0.6% | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | - | 1.1% | - | - | ||||
| Pinnularia laucensis Lange-Bertalot | mean | 0.4% | - | - | - | 0.7% | 1.5% | - | 0.5% | 0.172 | 0.712 | 0.662 |
| standard deviation | 0.7% | - | - | - | 1.4% | 3.0% | - | 1.1% | ||||
| Pinnularia microstauron var. rostrata Krammer | mean | 0.4% | - | - | - | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | 0.7% | - | - | - | - | - | - | - | ||||
| Pinnularia obscura Krasske | mean | - | - | - | - | 2.0% | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | 4.1% | - | - | - | ||||
| Pinnularia subanglica Krammer | mean | - | 0.8% | - | - | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | 1.5% | - | - | - | - | - | - | ||||
| Pinnularia subgibba Krammer | mean | - | - | - | 0.5% | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | 1.0% | - | - | - | - | ||||
| Pinnularia subinterrupta Krammer & Schroeter | mean | - | - | - | - | 0.3% | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | 0.6% | - | - | - | ||||
| Pinnularia sp. | mean | 1.4% | 0.2% | - | - | - | - | - | - | 0.217 | 0.951 | 0.488 |
| standard deviation | 2.7% | 0.5% | - | - | - | - | - | - | ||||
| Pinnularia sp.2 | mean | 0.2% | - | - | - | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | 0.4% | - | - | - | - | - | - | - | ||||
| Pinnularia tabellaria Ehrenberg | mean | 1.1% | - | - | - | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | 2.1% | - | - | - | - | - | - | - | ||||
| Placogeia kriegeri (K.Krasske) Bukhtiyarova | mean | - | - | - | 0.5% | - | 0.3% | - | - | 0.204 | 0.880 | 0.537 |
| standard deviation | - | - | - | 1.0% | - | 0.6% | - | - | ||||
| Placoneis elginensis (W.Gregory) E.J.Cox | mean | - | - | - | - | 0.7% | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | 1.4% | - | - | - | ||||
| Placoneis hambergii (Hustedt) K.Bruder | mean | - | - | - | - | - | - | - | 1.1% | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | - | - | - | 2.2% | ||||
| Planothidium frequentissimum (Lange-Bertalot) Lange-Bertalot | mean | - | - | - | - | 0.7% | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | 1.4% | - | - | - | ||||
| Psammothidium hustedtii (Krasske) Mayama | mean | - | - | - | - | - | - | 0.2% | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | - | - | 0.5% | - | ||||
| Rhopalodia gibberula var. vanheurckii O. Müller | mean | - | - | 0.4% | - | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | 0.8% | - | - | - | - | - | ||||
| Sellaphora laevissima (Kützing) D.G.Mann | mean | - | - | - | - | 0.5% | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | 1.0% | - | - | - | ||||
| Sellaphora nigri (De Notaris) Wetzel & Ector | mean | 9.1% | 9.3% | - | 18.8% | 6.0% | 1.0% | 14.5% | 4.4% | 0.324 | 1.641 | 0.172 |
| standard deviation | 8.0% | 10.7% | - | 20.9% | 5.6% | 1.9% | 11.5% | 5.4% | ||||
| Sellaphora sassiana (Metzeltin & Lange-Bertalot) | mean | - | - | 2.4% | 1.5% | - | - | - | - | 0.202 | 0.870 | 0.544 |
| standard deviation | - | - | 4.8% | 3.1% | - | - | - | - | ||||
| Sellaphora saugerresii (Desmazières) Wetzel & Mann | mean | - | 7.9% | 1.2% | 10.1% | 0.3% | - | 3.0% | 3.7% | 0.159 | 0.649 | 0.712 |
| standard deviation | - | 15.9% | 1.8% | 20.2% | 0.6% | - | 6.0% | 5.1% | ||||
| Sellaphora sp. | mean | - | - | - | 1.8% | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | 3.6% | - | - | - | - | ||||
| Sellaphora sp.2 | mean | - | 0.1% | - | - | - | - | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | 0.2% | - | - | - | - | - | - | ||||
| Spicaticribra kingstonii Johansen, Kociolek & Lowe | mean | - | - | 1.0% | - | - | - | - | 0.3% | 0.616 | 5.509 | 0.001 |
| standard deviation | - | - | 0.7% | - | - | - | - | 0.5% | ||||
| Stauroneis gracilis Ehrenberg | mean | - | - | - | - | - | 0.6% | - | - | 0.226 | 1.000 | 0.455 |
| standard deviation | - | - | - | - | - | 1.1% | - | - | ||||
| Surirella angusta Kützing | mean | 0.2% | - | - | 1.5% | - | - | - | - | 0.219 | 0.961 | 0.481 |
| standard deviation | 0.4% | - | - | 3.1% | - | - | - | - | ||||
| Surirella roba Leclercq | mean | - | 0.4% | - | - | 0.3% | - | - | - | 0.200 | 0.859 | 0.551 |
| standard deviation | - | 0.7% | - | - | 0.6% | - | - | - | ||||
| Ulnaria ulna (Nitzsch) Compère | mean | 0.2% | 0.2% | 0.1% | 1.5% | - | - | - | 2.1% | 0.247 | 1.126 | 0.380 |
| standard deviation | 0.4% | 0.5% | 0.3% | 3.1% | - | - | - | 3.1% |
References
- Tang, W.; Pei, Y.; Zheng, H.; Zhao, Y.; Shu, L.; Zhang, H. Twenty years of China’s water pollution control: Experiences and challenges. Chemosphere 2022, 295, 133875. [Google Scholar] [CrossRef]
- Elsheikh, A.H.; Panchal, H.N.; Sengottain, S.; Alsaleh, N.A.; Ahmadein, M. Applications of Heat Exchanger in Solar Desalination: Current Issues and Future Challenges. Water 2022, 14, 852. [Google Scholar] [CrossRef]
- Elsheikh, A.H.; Sharshir, S.W.; Mostafa, M.E.; Essa, F.A.; Ahmed Ali, M.K. Applications of nanofluids in solar energy: A review of recent advances. Renew. Sustain. Energy Rev. 2018, 82, 3483–3502. [Google Scholar] [CrossRef]
- Chiquito Gesualdo, G.; Sone, J.S.; de Galvão, C.O.; Martins, E.S.; Montenegro, S.M.G.L.; Tomasella, J.; Mendiondo, E.M. Unveiling water security in Brazil: Current challenges and future perspectives. Hydrol. Sci. J. 2021, 66, 759–768. [Google Scholar] [CrossRef]
- Getirana, A.; Libonati, R.; Cataldi, M. Brazil is in water crisis—It needs a drought plan. Nature 2021, 600, 218–220. [Google Scholar] [CrossRef]
- Kakore, B.G.; Mamun, M.; Lee, S.-J.; An, K.-G. Land-Use Pattern as a Key Factor Determining the Water Quality, Fish Guilds, and Ecological Health in Lotic Ecosystems of the Asian Monsoon Region. Water 2022, 14, 2765. [Google Scholar] [CrossRef]
- Albert, J.S.; Destouni, G.; Duke-Sylvester, S.M.; Magurran, A.E.; Oberdorff, T.; Reis, R.E.; Winemiller, K.O.; Ripple, W.J. Scientists’ warning to humanity on the freshwater biodiversity crisis. Ambio 2021, 50, 85–94. [Google Scholar] [CrossRef]
- Zhang, Y.; Peng, C.; Huang, S.; Wang, J.; Xiong, X.; Li, D. The relative role of spatial and environmental processes on seasonal variations of phytoplankton beta diversity along different anthropogenic disturbances of subtropical rivers in China. Environ. Sci. Pollut. Res. 2019, 26, 1422–1434. [Google Scholar] [CrossRef]
- Pineda, A.; Bortolini, J.C.; Rodrigues, L.C. Effects of space and environment on phytoplankton distribution in subtropical reservoirs depend on functional features of the species. Aquat. Sci. 2022, 84, 5. [Google Scholar] [CrossRef]
- Zhou, S.; Wu, N.; Zhang, M.; Peng, W.; He, F.; Guo, K.; Yan, S.; Zou, Y.; Qu, X. Local environmental, geo-climatic and spatial factors interact to drive community distributions and diversity patterns of stream benthic algae, macroinvertebrates and fishes in a large basin, Northeast China. Ecol. Indic. 2020, 117, 106673. [Google Scholar] [CrossRef]
- Huszar, V.L.M.; Nabout, J.C.; Appel, M.O.; Santos, J.B.O.; Abe, D.S.; Silva, L.H.S. Environmental and not spatial processes (directional and non-directional) shape the phytoplankton composition and functional groups in a large subtropical river basin. J. Plankton Res. 2015, 37, 1190–1200. [Google Scholar] [CrossRef] [Green Version]
- Leibold, M.A.; Holyoak, M.; Mouquet, N.; Amarasekare, P.; Chase, J.M.; Hoopes, M.F.; Holt, R.D.; Shurin, J.B.; Law, R.; Tilman, D.; et al. The metacommunity concept: A framework for multi-scale community ecology. Ecol. Lett. 2004, 7, 601–613. [Google Scholar] [CrossRef]
- Lansac-Tôha, F.M.; Bini, L.M.; Heino, J.; Meira, B.R.; Segovia, B.T.; Pavanelli, C.S.; Bonecker, C.C.; Deus, C.P.; Benedito, E.; Alves, G.M.; et al. Scale-dependent patterns of metacommunity structuring in aquatic organisms across floodplain systems. J. Biogeogr. 2021, 48, 872–885. [Google Scholar] [CrossRef]
- Rodriguez, J.M.M. Geoecologia das Paisagens: Uma visão Geossistêmica da Análise Ambiental, 6th ed.; Rodriguez, J.M.M., da Silva, E.V., de Paula Brito Cavalcanti, A., Eds.; Imprensa Universitária: Fortaleza, Brazil, 2021; Volume 41, ISBN 8572821481. [Google Scholar]
- Stenger-Kovács, C.; Lengyel, E.; Sebestyén, V.; Szabó, B. Effects of land use on streams: Traditional and functional analyses of benthic diatoms. Hydrobiologia 2020, 847, 2933–2946. [Google Scholar] [CrossRef]
- Li, C.; Wang, J.; Jiang, Y.; Bai, Y.; Cheng, L. The evolution of different dissolved organic matter components and release characteristics of heavy metals in leaching process from sewage sludge under simulated rain. Environ. Sci. Pollut. Res. 2022, 29, 86651–86664. [Google Scholar] [CrossRef]
- Docile, T.; Rosa, D.C.O.; Figueiró, R.; Nessimian, J. Urbanisation alters the flow of energy through stream food webs. Insect Conserv. Divers. 2016, 9, 416–426. [Google Scholar] [CrossRef]
- Barnum, T.R.; Weller, D.E.; Williams, M. Urbanization reduces and homogenizes trait diversity in stream macroinvertebrate communities. Ecol. Appl. 2017, 27, 2428–2442. [Google Scholar] [CrossRef]
- Heino, J.; Melo, A.S.; Siqueira, T.; Soininen, J.; Valanko, S.; Bini, L.M. Metacommunity organisation, spatial extent and dispersal in aquatic systems: Patterns, processes and prospects. Freshw. Biol. 2015, 60, 845–869. [Google Scholar] [CrossRef]
- Soininen, J.; Teittinen, A. Fifteen important questions in the spatial ecology of diatoms. Freshw. Biol. 2019, 64, 2071–2083. [Google Scholar] [CrossRef] [Green Version]
- Soininen, J. Environmental and spatial control of freshwater diatoms—A review. Diatom Res. 2007, 22, 473–490. [Google Scholar] [CrossRef]
- Costa, A.P.T.; Schneck, F. Diatoms as indicators in running waters: Trends of studies on biological assessment and monitoring. Environ. Monit. Assess. 2022, 194, 695. [Google Scholar] [CrossRef] [PubMed]
- Zorzal-Almeida, S.; Lehmkuhl, E.A.; Marquardt, G.C.; Morais, K.S.; Trancoso, M.S.; Silva-Lehmkuhl, A.M. 177 years of diatom studies in Brazil: Knowledge, gaps, and perspectives. An. Acad. Bras. Cienc. 2022, 94, e20210959. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Song, Z.; Zhou, J.; Soininen, J.; Tan, L.; Cai, Q.; Tang, T. Differences in diversity and community assembly processes between planktonic and benthic diatoms in the upper reach of the Jinsha River, China. Hydrobiologia 2022, 849, 1577–1591. [Google Scholar] [CrossRef]
- Maurya, P.K.; Ali, S.A.; Alharbi, R.S.; Yadav, K.K.; Alfaisal, F.M.; Ahmad, A.; Ditthakit, P.; Prasad, S.; Jung, Y.-K.; Jeon, B.-H. Impacts of Land Use Change on Water Quality Index in the Upper Ganges River near Haridwar, Uttarakhand: A GIS-Based Analysis. Water 2021, 13, 3572. [Google Scholar] [CrossRef]
- Khouni, I.; Louhichi, G.; Ghrabi, A. Use of GIS based Inverse Distance Weighted interpolation to assess surface water quality: Case of Wadi El Bey, Tunisia. Environ. Technol. Innov. 2021, 24, 101892. [Google Scholar] [CrossRef]
- Setia, R.; Lamba, S.; Chander, S.; Kumar, V.; Singh, R.; Litoria, P.K.; Singh, R.P.; Pateriya, B. Spatio-temporal variations in water quality, hydrochemistry and its controlling factors in a perennial river in India. Appl. Water Sci. 2021, 11, 169. [Google Scholar] [CrossRef]
- Shurin, J.B.; Cottenie, K.; Hillebrand, H. Spatial autocorrelation and dispersal limitation in freshwater organisms. Oecologia 2009, 159, 151–159. [Google Scholar] [CrossRef]
- Manning, F.S.; Curtis, P.J.; Walker, I.R.; Pither, J. Potential long-distance dispersal of freshwater diatoms adhering to waterfowl plumage. Freshw. Biol. 2021, 66, 1136–1148. [Google Scholar] [CrossRef]
- Jamoneau, A.; Passy, S.I.; Soininen, J.; Leboucher, T.; Tison-Rosebery, J. Beta diversity of diatom species and ecological guilds: Response to environmental and spatial mechanisms along the stream watercourse. Freshw. Biol. 2018, 63, 62–73. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Soininen, J.; Han, B.-P.; Declerck, S.A.J. Effects of connectivity, dispersal directionality and functional traits on the metacommunity structure of river benthic diatoms. J. Biogeogr. 2013, 40, 2238–2248. [Google Scholar] [CrossRef]
- Padial, A.A.; Ceschin, F.; Declerck, S.A.J.; De Meester, L.; Bonecker, C.C.; Lansac-Tôha, F.A.; Rodrigues, L.; Rodrigues, L.C.; Train, S.; Velho, L.F.M.; et al. Dispersal Ability Determines the Role of Environmental, Spatial and Temporal Drivers of Metacommunity Structure. PLoS ONE 2014, 9, e111227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortolini, J.C.; Pineda, A.; Rodrigues, L.C.; Jati, S.; Velho, L.F.M. Environmental and spatial processes influencing phytoplankton biomass along a reservoirs-river-floodplain lakes gradient: A metacommunity approach. Freshw. Biol. 2017, 62, 1756–1767. [Google Scholar] [CrossRef]
- Zorzal-Almeida, S.; Soininen, J.; Bini, L.M.; Bicudo, D.C. Local environment and connectivity are the main drivers of diatom species composition and trait variation in a set of tropical reservoirs. Freshw. Biol. 2017, 62, 1551–1563. [Google Scholar] [CrossRef]
- De Oliveira, P.H.F.; Machado, K.B.; Teresa, F.B.; Heino, J.; Nabout, J.C. Spatial processes determine planktonic diatom metacommunity structure of headwater streams. Limnologica 2020, 84, 125813. [Google Scholar] [CrossRef]
- Medeiros, G.; Padial, A.A.; Wedig Amaral, M.W.; Ludwig, T.A.V.; Bueno, N.C. Environmental variables likely influence the periphytic diatom community in a subtropical lotic environment. Limnologica 2020, 80, 125718. [Google Scholar] [CrossRef]
- Tonkin, J.D.; Heino, J.; Altermatt, F. Metacommunities in river networks: The importance of network structure and connectivity on patterns and processes. Freshw. Biol. 2018, 63, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Tonkin, J.D.; Altermatt, F.; Finn, D.S.; Heino, J.; Olden, J.D.; Pauls, S.U.; Lytle, D.A. The role of dispersal in river network metacommunities: Patterns, processes, and pathways. Freshw. Biol. 2018, 63, 141–163. [Google Scholar] [CrossRef] [Green Version]
- Wojciechowski, J.; Heino, J.; Bini, L.M.; Padial, A.A. The strength of species sorting of phytoplankton communities is temporally variable in subtropical reservoirs. Hydrobiologia 2017, 800, 31–43. [Google Scholar] [CrossRef]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Gonc, L.D.M.; Sparovek, G. Köppen’s climate classification map for Brazil Clayton. Meteorol. Z. 2013, 22, 711–728. [Google Scholar] [CrossRef]
- Cembranel, A.S.; Sampaio, S.C.; Remor, M.B.; Gotardo, J.T.; Rosa, P.M.D. Geochemical background in an oxisol. Eng. Agrícola 2017, 37, 565–573. [Google Scholar] [CrossRef]
- de Cascavel, P.M. Recuperação Ambiental da Bacia Hidrográfica do Rio Cascavel; FUNDETEC: Sincelejo, Colombia, 1995. [Google Scholar]
- Instituto Brasileiro de Geografia e Estatística. Manual Técnico de Uso da Terra; Instituto Brasileiro de Geografia e Estatística: Rio de Janeiro, Brazil, 2013; ISBN 9788524043079.
- APHA. Standard Methods for the Examination of Water and Wastewater, 22nd ed.; Franson, M.A.H., Ed.; Copyright: Washington, DC, USA, 2012; ISBN 978-0875530130. [Google Scholar]
- CETESB. ANA Guia Nacional de Coleta e Preservação de Amostras, 2nd ed.; Brandão, C.J., Botelho, M.J.C., Sato, M.I.Z., Lamparelli, M.C., Eds.; CETESB: São Paulo, Brazil, 2011.
- USEPA 3050B. Acid Digestion of Sediments, Sludges, and Soils; USEPA: Washington, DC, USA, 1996; pp. 1–12.
- Borcard, D.; Legendre, P. All-scale spatial analysis of ecological data by means of principal coordinates of neighbour matrices. Ecol. Modell. 2002, 153, 51–68. [Google Scholar] [CrossRef]
- Blanchet, F.G.; Legendre, P.; Borcard, D. Modelling directional spatial processes in ecological data. Ecol. Modell. 2008, 215, 325–336. [Google Scholar] [CrossRef]
- Simonsen, R. The Diatom Plankton of the Indian Ocean Expedition of RV “Meteor”, 1964–1965; Borntraeger: Berlin, Germany, 1974; ISBN 3443200192 9783443200190. [Google Scholar]
- Moreira-Filho, H.; Valente Mreira, I. Avaliacao taxonomica e ecologica das Diatomaceas (Bacillariophyceae) epifitas em algas pluricelulares obtidas nos litorais dos Estados do Parana, Santa Catarina e Sao Paulo. Bol. Mus. Bot. Munic. 1981, 1, 17. [Google Scholar]
- Kobayasi, H.; Mayama, S. Most pollution-tolerant diatoms of severely polluted rivers in the vicinity of Tokyo. Jpn. J. Phycol. 1982, 30, 188–196. [Google Scholar]
- Pappas, J.L.; Stoermer, E.F. Quantitative method for determining a representative algal sample count. J. Phycol. 1996, 32, 693–696. [Google Scholar] [CrossRef] [Green Version]
- Battarbee, R.W. Diatom Analysis. In Handbook of Holocene Palaeoecology and Palaeohydrology; Berglund, B.E., Ralska-Jasiewiczowa, M., Eds.; International Geological Correlation Programme; Wiley: New York, NY, USA, 1986; p. 869. ISBN 9780471906919. [Google Scholar]
- Lobo, E.A.; Katoh, K.; Aruga, Y. Response of epilithic diatom assemblages to water pollution in rivers located in the Tokyo Metropolitan area, Japan. Freshw. Biol. 1995, 34, 191–204. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology, Volume 24. Dev. Environ. Model. 1988, 24, 870. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Gower, J.C. Some distance properties of latent root and vector methods used in multivariate analysis. Biometrika 1966, 53, 325–338. [Google Scholar] [CrossRef]
- Tuomisto, H.; Ruokolainen, K. Analyzing or explaining beta diversity? Understanding the targets of different methods of analysis. Ecology 2006, 87, 2697–2708. [Google Scholar] [CrossRef]
- Radbruch, L.; De Lima, L.; Knaul, F.; Wenk, R.; Ali, Z.; Bhatnaghar, S.; Blanchard, C.; Bruera, E.; Buitrago, R.; Burla, C.; et al. Redefining Palliative Care—A New Consensus-Based Definition. J. Pain Symptom Manag. 2020, 60, 754–764. [Google Scholar] [CrossRef]
- Ntongani, W.A.; Andrew, S.M.; R Development Core Team. R: A Language and Environment for Statistical Computing; R Development Core Team: Vienna, Austria, 2011; Volume 1, ISBN 3900051070. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package “Vegan” Title Community Ecology Package Version 2.5-7. 2020. [Google Scholar]
- Dray, S.; Pélissier, R.; Couteron, P.; Fortin, M.-J.; Legendre, P.; Peres-Neto, P.R.; Bellier, E.; Bivand, R.; Blanchet, F.G.; De Cáceres, M.; et al. Community ecology in the age of multivariate multiscale spatial analysis. Ecol. Monogr. 2012, 82, 257–275. [Google Scholar] [CrossRef]
- Bivand, R. R Packages for Analyzing Spatial Data: A Comparative Case Study with Areal Data. Geogr. Anal. 2022, 54, 488–518. [Google Scholar] [CrossRef]
- Cima, E.G.; Uribe-Opazo, M.A.; Da Rocha Junior, W.F.; De Souza Fragoso, R.M. A spatial analysis of western paranÁ: Scenarios for regional development. Rev. Bras. Gest. E Desenvolv. Reg. 2021, 17, 151–164. [Google Scholar]
- Guicho, R.; Ferreira, J.H.D.; Medeiros, G.; Siqueira, J.A.C.; de Souza, S.N.M.; Prior, M. Method for estimating the wind power micro and minigeneration applied to a city with a subtropical climate in South America. Res. Soc. Dev. 2021, 10, e105101220009. [Google Scholar] [CrossRef]
- da Silva, J.N.; da Cunha, L.S.; de Oliveira, N.E.C. O Uso De Parâmetros Físico-Químicos Na Delimitação De Contaminação Por Lixiviado Em Áreas Degradadas Por Resíduos Sólidos Urbanos. Uma Revisão Sistemática De Literatura Com Ênfase Em Metanálise. Rev. Bras. Geogr. Física 2022, 15, 1587. [Google Scholar] [CrossRef]
- De Almeida, T.P.; Macena, D.Â.; Simões, J.S.T.; Mareco, E.A.; Calciolari Rossi, R.; Favareto, A.P.A. Análise de parâmetros de qualidade da água e teste de genotoxicidade em peixes da bacia hidrográfica do rio Pirapozinho—SP, Brasil. Res. Soc. Dev. 2022, 11, e46711319309. [Google Scholar] [CrossRef]
- Peres, K.K.; Guicho, R.; Medeiros, G.; Amaral, M.W.W.; da Silva, T.T.; Pilatti, M.C.; Prior, M.; Bueno, N.C. Environmental fragility as an indicator of the risk of contamination by human action in watersheds used for public supply in western Paraná, Brazil. Environ. Earth Sci. 2022, 81, 486. [Google Scholar] [CrossRef]
- De Carvalho, L.H.W.; Laudares, S.; Libório, M.P.; Ekel, M.P.; de Oliveira Marques, R. Urban Settlement Spatial Analysis in Permanent Preservation Area of M’boicy Watershed River, Foz do Iguassu City in Brazil. Int. J. Geosci. 2016, 7, 1222–1231. [Google Scholar] [CrossRef] [Green Version]
- Odeh, T.; Mohammad, A.H.; Pradhanang, S.M.; Ismail, M.; Rödiger, T. GIS-based analytical modeling on evaluating impacts of urbanization in Amman water resources, Jordan. Environ. Earth Sci. 2022, 81, 160. [Google Scholar] [CrossRef]
- Passy, S.I.; Larson, C.A.; Jamoneau, A.; Budnick, W.; Heino, J.; Leboucher, T.; Tison-Rosebery, J.; Soininen, J. Biogeographical patterns of species richness and abundance distribution in stream diatoms are driven by climate and water chemistry. Am. Nat. 2018, 192, 605–617. [Google Scholar] [CrossRef] [Green Version]
- Erős, T.; Lowe, W.H. The Landscape Ecology of Rivers: From Patch-Based to Spatial Network Analyses. Curr. Landsc. Ecol. Rep. 2019, 4, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Vilmi, A.; Karjalainen, S.M.; Hellsten, S.; Heino, J. Bioassessment in a metacommunity context: Are diatom communities structured solely by species sorting? Ecol. Indic. 2016, 62, 86–94. [Google Scholar] [CrossRef]
- Viana, D.S.; Chase, J.M. Spatial scale modulates the inference of metacommunity assembly processes. Ecology 2019, 100, e02576. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Soininen, J.; Chen, K.; Wang, B. Environmental Factors Override Dispersal-Related Factors in Shaping Diatom and Macroinvertebrate Communities Within Stream Networks in China. Front. Ecol. Evol. 2020, 8, 141. [Google Scholar] [CrossRef]
- Ferraz Sampaio, M.; Ferreira, P.C.; Veiga Ludwig, T.A.; Padial, A.A. Temporal stability in beta diversity does not guarantee surrogacy or compositional stability in a micro-phytoplankton metacommunity. Limnetica 2023, 42, 1. [Google Scholar] [CrossRef]
- Marquardt, G.C.; Padial, A.A.; Bicudo, C.E.M. Variance partitioning of deconstructed tropical diatom communities in reservoirs cascade. Aquat. Sci. 2018, 80, 17. [Google Scholar] [CrossRef]
- Vilmi, A.; Tolonen, K.T.; Karjalainen, S.M.; Heino, J. Metacommunity structuring in a highly-connected aquatic system: Effects of dispersal, abiotic environment and grazing pressure on microalgal guilds. Hydrobiologia 2017, 790, 125–140. [Google Scholar] [CrossRef]
- Gilbert, B.; Lechowicz, M.J. Neutrality, niches, and dispersal in a temperate forest understory. Proc. Natl. Acad. Sci. USA 2004, 101, 7651–7656. [Google Scholar] [CrossRef] [Green Version]
- Kaushal, S.S.; Belt, K.T. The urban watershed continuum: Evolving spatial and temporal dimensions. Urban Ecosyst. 2012, 15, 409–435. [Google Scholar] [CrossRef]
- Mutinova, P.T.; Kahlert, M.; Kupilas, B.; McKie, B.G.; Friberg, N.; Burdon, F.J. Benthic Diatom Communities in Urban Streams and the Role of Riparian Buffers. Water 2020, 12, 2799. [Google Scholar] [CrossRef]
- Foets, J.; Wetzel, C.E.; Teuling, A.J.; Pfister, L. Temporal and spatial variability of terrestrial diatoms at the catchment scale: Controls on communities. PeerJ 2020, 8, e8296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Şahin, B.; Barınova, S. Role of Altitude in Formation of Diatom Diversity of High Mountain Protected Glacier Lakes in the Kaçkar Mountains National Park, Rize, Turkey. Environments 2022, 9, 127. [Google Scholar] [CrossRef]
- Poradowska, A. Diatoms (Bacillariophyta) from the Genus Eunotia and Pinnularia developing on Soils in the Open Landscape of the Low Beskids. J. Ecol. Eng. 2020, 21, 257–270. [Google Scholar] [CrossRef]
- Soininen, J.; Paavola, R.; Muotka, T. Benthic diatom communities in boreal streams: Community structure in relation to environmental and spatial gradients. Ecography 2004, 27, 330–342. [Google Scholar] [CrossRef]
- Tuji, A.; Williams, D.M. Type Examination of Japanese Diatoms Described by Friedrich Meister (1913) from Lake Suwa. Natl. Sci. Mus. 2007, 33, 69–79. [Google Scholar]
- Lobo, E.A.; Schuch, M.; Heinrich, C.G.; Ben, A.; Düpont, A.; Wetzel, C.E.; Ector, L. Development of the Trophic Water Quality Index (TWQI) for subtropical temperate Brazilian lotic systems. Environ. Monit Assess 2015, 187, 354. [Google Scholar] [CrossRef]
- Levkov, Z.; Mitic-Kopanja, D.; Reichardt, E. The Diatom Genus Gomphonema from the Republic og Macedonia; Lange-Bertalot, H., Ed.; Diatoms of Europe; Koeltz Botanical Books: Oberreifenberg, Germany, 2016; ISBN 9783946583004/3946583008. [Google Scholar]
- Domingues, R.B.; Guerra, C.C.; Barbosa, A.B.; Galvão, H.M. Are nutrients and light limiting summer phytoplankton in a temperate coastal lagoon? Aquat. Ecol. 2015, 49, 127–146. [Google Scholar] [CrossRef]
- Soininen, J.; Jamoneau, A.; Rosebery, J.; Passy, S.I. Global patterns and drivers of species and trait composition in diatoms. Glob. Ecol. Biogeogr. 2016, 25, 940–950. [Google Scholar] [CrossRef]
- Lange, K.; Liess, A.; Piggott, J.J.; Townsend, C.R.; Matthaei, C.D. Light, nutrients and grazing interact to determine stream diatom community composition and functional group structure. Freshw. Biol. 2011, 56, 264–278. [Google Scholar] [CrossRef]
- Tapolczai, K.; Bouchez, A.; Stenger-Kovács, C.; Padisák, J.; Rimet, F. Taxonomy- or trait-based ecological assessment for tropical rivers? Case study on benthic diatoms in Mayotte island (France, Indian Ocean). Sci. Total Environ. 2017, 607–608, 1293–1303. [Google Scholar] [CrossRef]
- Stenger-Kovács, C.; Lengyel, E.; Buczkó, K.; Padisák, J.; Korponai, J. Trait-based diatom functional diversity as an appropriate tool for understanding the effects of environmental changes in soda pans. Ecol. Evol. 2020, 10, 320–335. [Google Scholar] [CrossRef] [Green Version]
- Jüttner, I.; Ector, L.; Reichardt, E.; Van De Vijver, B.; Jarlman, A.; Krokowski, J.; Cox, E.J. Gomphonema varioreduncum sp. Nov., a new species from northern and western Europe and a re-examination of Gomphonema exilissimum. Diatom Res. 2013, 28, 303–316. [Google Scholar] [CrossRef]
- Faustino, S.B.; Fontana, L.; Cristina, E.; Bartozek, R.; Eduardo, C.; Bicudo, D.M.; Bicudo, D.D.C. Composition and distribution of diatom assemblages from core and surface sediments of a water supply reservoir in Southeastern Brazil. Biota Neotrop. 2016, 16, 1–23. [Google Scholar] [CrossRef]
- Marra, R.C.; Tremarin, P.I.; Algarte, V.M.; Ludwig, T.A.V. Epiphytic diatoms (Diatomeae) from Piraquara II urban reservoir, Paraná state. Biota Neotrop. 2016, 16, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Pillsbury, R.; Stevenson, R.J.; Munn, M.D.; Waite, I. Relationships between diatom metrics based on species nutrient traits and agricultural land use. Environ. Monit. Assess. 2019, 191, 228. [Google Scholar] [CrossRef]
- Nystrand, M.I.; Österholm, P.; Nyberg, M.E.; Gustafsson, J.P. Metal speciation in rivers affected by enhanced soil erosion and acidity. Appl. Geochem. 2012, 27, 906–916. [Google Scholar] [CrossRef]
- Martins, W.A.; Martins, L.L.; de Maria, I.C.; de Moraes, J.F.L.; Pedro Júnior, M.J. Reduction of sediment yield by riparian vegetation recovery at distinct levels of soil erosion in a tropical watershed. Ciência Agrotecnologia 2021, 45, e028220. [Google Scholar] [CrossRef]
- Falasco, E.; Ector, L.; Wetzel, C.E.; Badino, G.; Bona, F. Looking back, looking forward: A review of the new literature on diatom teratological forms (2010–2020). Hydrobiologia 2021, 848, 1675–1753. [Google Scholar] [CrossRef]
- Morin, S.; Cordonier, A.; Lavoie, I.; Arini, A.; Blanco, S.; Duong, T.T.; Torne’s, E.; Bonet, B.; Corcoll, N.; Faggiano, L.; et al. Consistency in Diatom Response. In Emerging and Priority Pollutants in Rivers; Lavoie, I., Fortin, C., Arini, A., Blanco, S., Becares, E., Duong, T.T., Tornés, E., Torne’s, E., Sabater, S., Bonet, B., et al., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 118–146. ISBN 9783642257223. [Google Scholar]
- Pandey, L.K.; Lavoie, I.; Morin, S.; Park, J.; Lyu, J.; Choi, S.; Lee, H.; Han, T. River water quality assessment based on a multi-descriptor approach including chemistry, diatom assemblage structure, and non-taxonomical diatom metrics. Ecol. Indic. 2018, 84, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, H.G.; Bhering, S.B.; Bognola, I.A.; Curcio, G.R.; de Carvalho Junior, W.; Chagas, C.D.S.; Manzatto, C.V.; Aglio, M.L.D.; Silva, J.D.S.; Chaffin, C.E. Mapa de Solos do Estado do Paraná. Embrapa Solos 2007, 96, 73. [Google Scholar]
- Costa, L.F.; Wetzel, C.E.; Lange-Bertalot, H.; Ector, L.; Bicudo, D.C. Taxonomy and Ecology of Eunotia Species (Bacillariophyta) in Southeastern Brazilian Reservoirs; Cramer: Stuttgart, Germany, 2017; ISBN 9783443570552. [Google Scholar]
- Amaral, M.W.W.; Medeiros, G.; Daufenbach, V.; Suszek Gonçalves, M.; Alvim Veiga Ludwig, T.; Catarina Bueno, N. Nupela semifasciata (Bacillariophyceae), a new species from subtropical lotic environments in Western Paraná State, Brazil. Fottea 2021, 21, 152–163. [Google Scholar] [CrossRef]
- Ortiz-Colón, A.I.; Piñero-Santiago, L.E.; Rivera, N.M.; Sosa, M.A. HHS Public Access. J. Env. Anal Toxicol 2016, 6, 1000353. [Google Scholar] [CrossRef]
- Tao, Y.; Yuan, Z.; Xiaona, H.; Wei, M. Ecotoxicology and Environmental Safety Distribution and bioaccumulation of heavy metals in aquatic organisms of different trophic levels and potential health risk assessment from. Ecotoxicol. Environ. Saf. 2012, 81, 55–64. [Google Scholar] [CrossRef]
- Virtanen, L.; Soininen, J. The roles of environment and space in shaping stream diatom communities. Eur. J. Phycol. 2012, 47, 160–168. [Google Scholar] [CrossRef]
- Pajunen, V.; Luoto, M.; Soininen, J. Climate is an important driver for stream diatom distributions. Glob. Ecol. Biogeogr. 2016, 25, 198–206. [Google Scholar] [CrossRef]
- Zorzal-Almeida, S.; Salim, A.; Andrade, M.R.M.; de Nascimento, M.N.; Bini, L.M.; Bicudo, D.C. Effects of land use and spatial processes in water and surface sediment of tropical reservoirs at local and regional scales. Sci. Total Environ. 2018, 644, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, M.; Launay, B.; Le Goff, G.; Pella, H.; Douady, C.J.; Datry, T. Fragmentation promotes the role of dispersal in determining 10 intermittent headwater stream metacommunities. Freshw. Biol. 2020, 65, 2169–2185. [Google Scholar] [CrossRef]
- Cook, J.R. Decompressive Procedures. Vet. Clin. N. Am. Small Anim. Pract. 1992, 22, 917–921. [Google Scholar] [CrossRef] [PubMed]
- Vilmi, A.; Karjalainen, S.M.; Heino, J. Ecological uniqueness of stream and lake diatom communities shows different macroecological patterns. Divers. Distrib. 2017, 23, 1042–1053. [Google Scholar] [CrossRef] [Green Version]
- Wetzel, C.E.; de Bicudo, D.C.; Ector, L.; Lobo, E.A.; Soininen, J.; Landeiro, V.L.; Bini, L.M. Distance Decay of Similarity in Neotropical Diatom Communities. PLoS ONE 2012, 7, e45071. [Google Scholar] [CrossRef] [PubMed]
- Bried, J.T.; Vilmi, A. Improved detection of mass effect species assembly for applied metacommunity thinking. J. Appl. Ecol. 2022, 59, 921–926. [Google Scholar] [CrossRef]
- Record, S.; Voelker, N.M.; Zarnetske, P.L.; Wisnoski, N.I.; Tonkin, J.D.; Swan, C.; Marazzi, L.; Lany, N.; Lamy, T.; Compagnoni, A.; et al. Novel Insights to Be Gained From Applying Metacommunity Theory to Long-Term, Spatially Replicated Biodiversity Data. Front. Ecol. Evol. 2021, 8, 612794. [Google Scholar] [CrossRef]





| Sampling Sites | Geographic Coordinates | |
|---|---|---|
| Latitude | Longitude | |
| S1 | 24°57′32.93″ S | 53°26′33.32″ W |
| S2 | 24°58′35.81″ S | 53°26′6.85″ W |
| S3 | 25°0′13.36″ S | 53°26′19.12″ W |
| S4 | 24°58′17.29″ S | 53°24′24.75″ W |
| S5 | 24°59′5.99″ S | 53°25′23.83″ W |
| S6 | 24°59′18.90″ S | 53°25′32.20″ W |
| S7 | 24°59′48.82″ S | 53°26′42.08″ W |
| S8 | 25°0′13.36″ S | 53°26′19.12″ W |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medeiros, G.; Padial, A.A.; Amaral, M.W.W.; Guicho, R.; Pilatti, M.C.; Sampaio, S.C.; Ludwig, T.A.V.; Bueno, N.C.; dos Reis, R.R. Exploring Key Determinants of the Periphytic Diatom Community in a Southern Brazilian Micro-Watershed. Water 2022, 14, 3913. https://doi.org/10.3390/w14233913
Medeiros G, Padial AA, Amaral MWW, Guicho R, Pilatti MC, Sampaio SC, Ludwig TAV, Bueno NC, dos Reis RR. Exploring Key Determinants of the Periphytic Diatom Community in a Southern Brazilian Micro-Watershed. Water. 2022; 14(23):3913. https://doi.org/10.3390/w14233913
Chicago/Turabian StyleMedeiros, Gabriela, André Andrian Padial, Mailor Wellinton Wedig Amaral, Ricardo Guicho, Maria Clara Pilatti, Silvio Cesar Sampaio, Thelma Alvim Veiga Ludwig, Norma Catarina Bueno, and Ralpho Rinaldo dos Reis. 2022. "Exploring Key Determinants of the Periphytic Diatom Community in a Southern Brazilian Micro-Watershed" Water 14, no. 23: 3913. https://doi.org/10.3390/w14233913
APA StyleMedeiros, G., Padial, A. A., Amaral, M. W. W., Guicho, R., Pilatti, M. C., Sampaio, S. C., Ludwig, T. A. V., Bueno, N. C., & dos Reis, R. R. (2022). Exploring Key Determinants of the Periphytic Diatom Community in a Southern Brazilian Micro-Watershed. Water, 14(23), 3913. https://doi.org/10.3390/w14233913