

Effects of Hypoxia and Hypomagnetic Field on Morphometric and Life-History Traits in Freshwater Cladoceran Daphnia magna


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Daphnia magna
2.2. Oxygen
2.3. Magnetic Conditions
2.4. Experimental Procedure and Evaluated Parameters
- GMF, dissolved oxygen 8 mg/L;
- GMF, dissolved oxygen 5 mg/L;
- GMF, dissolved oxygen 2 mg/L;
- HMF, dissolved oxygen 8 mg/L;
- HMF, dissolved oxygen 5 mg/L;
- HMF, dissolved oxygen 2 mg/L.
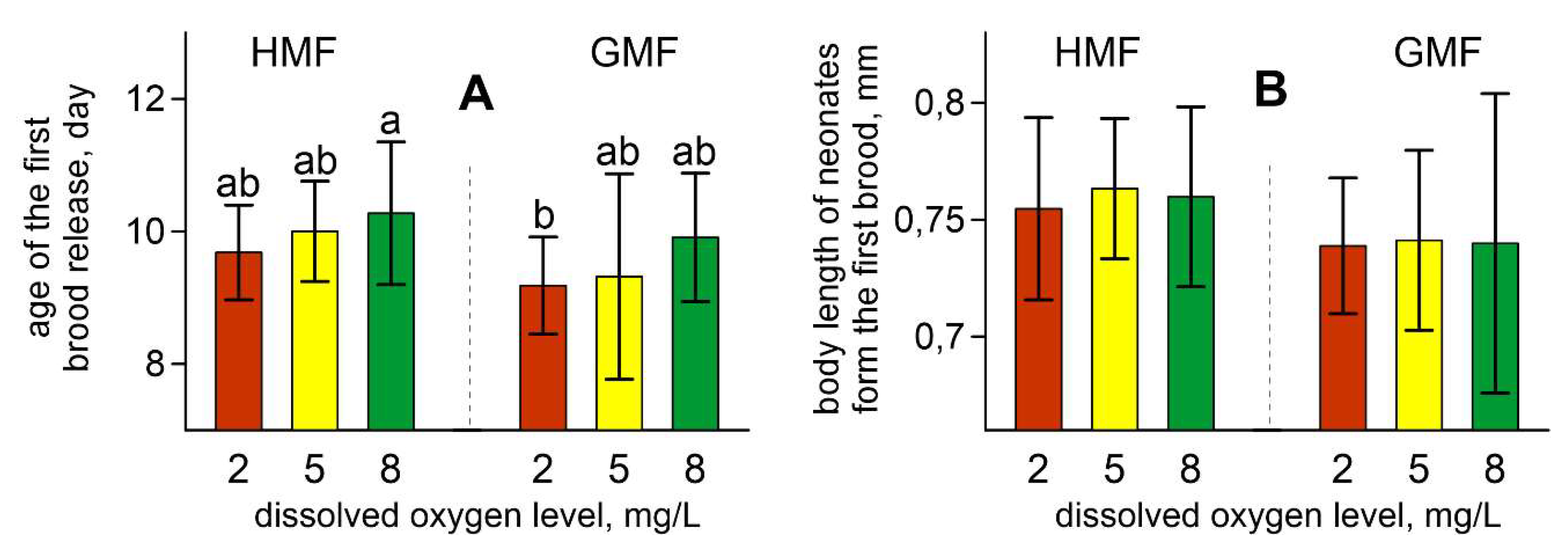
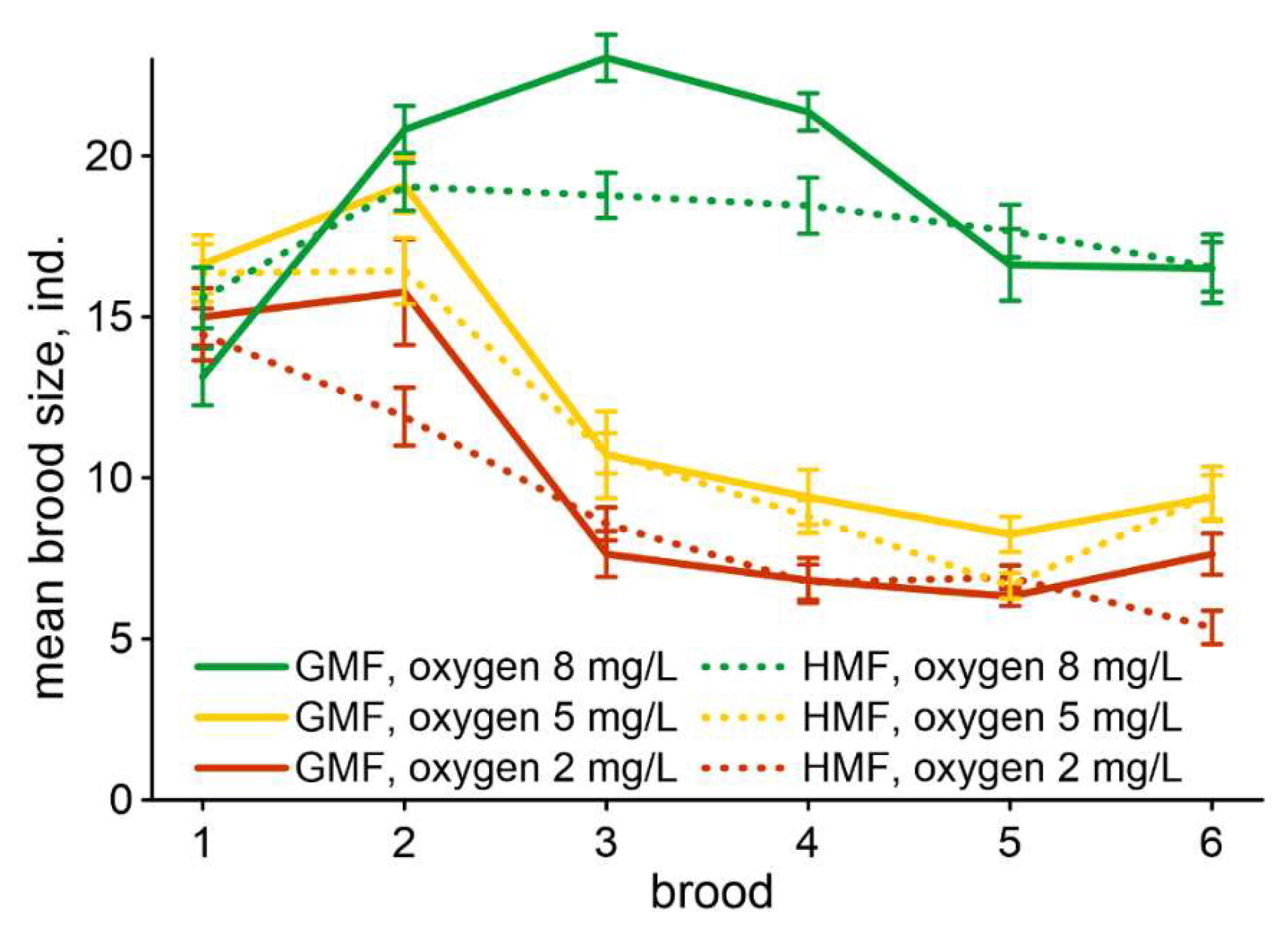
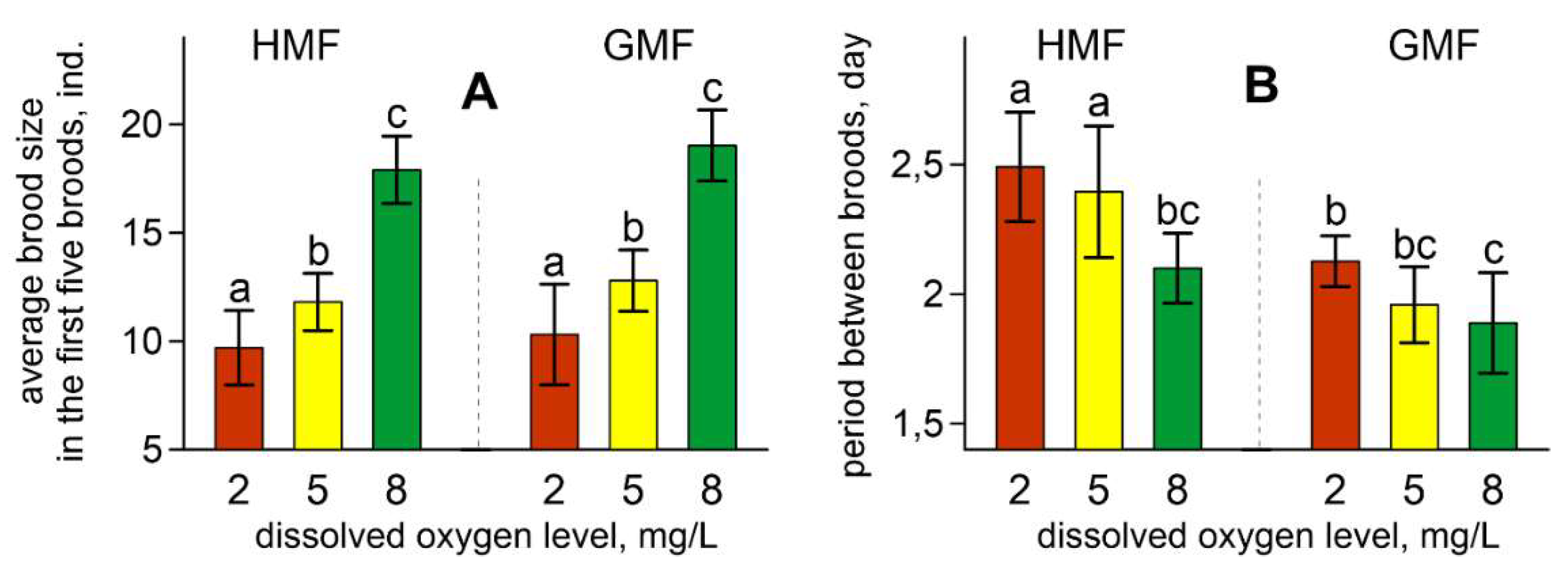
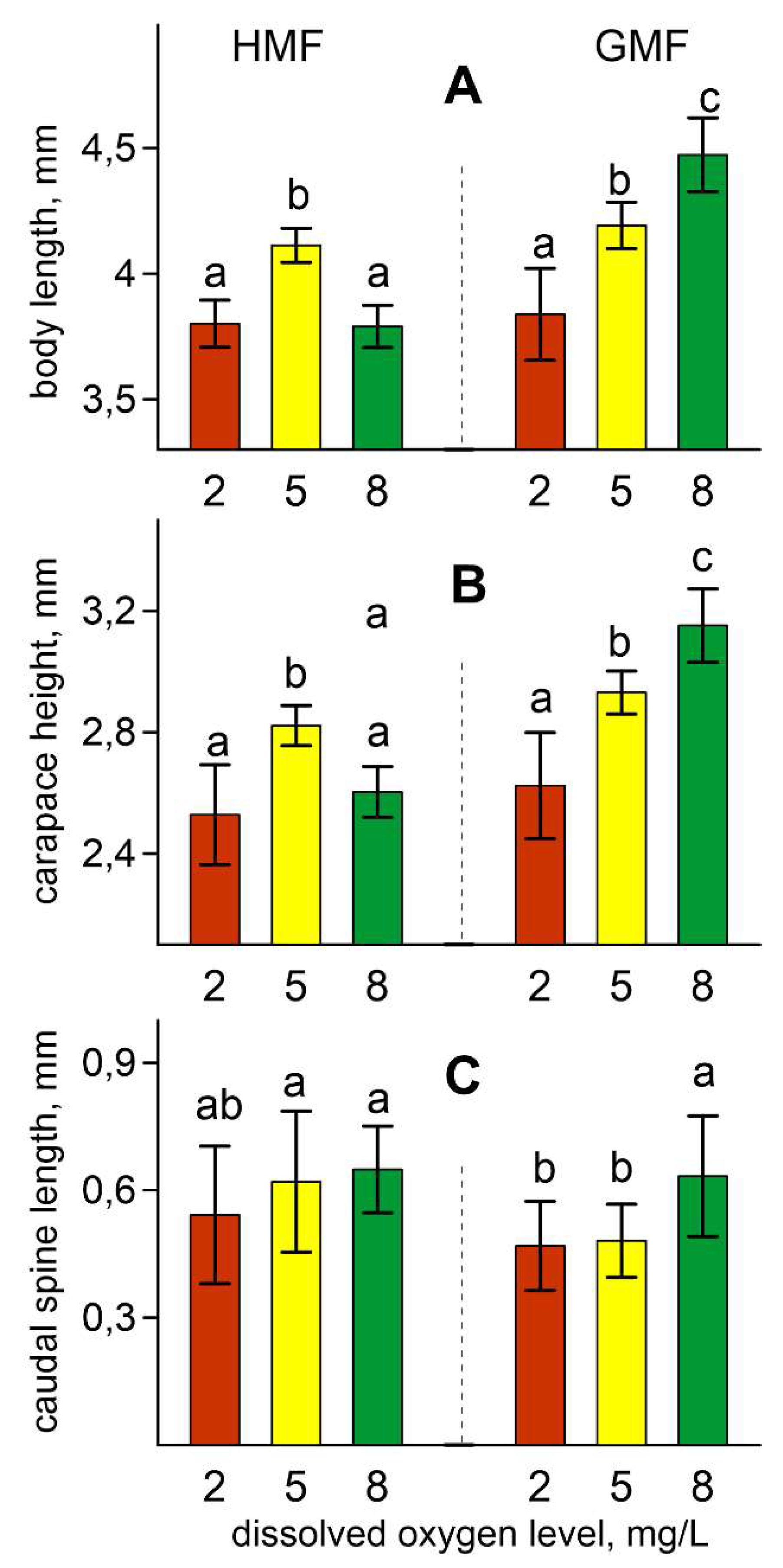
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Breitburg, D.; Levin, L.A.; Oschlies, A.; Grégoire, M.; Chavez, F.P.; Conley, D.J.; Garçon, V.; Gilbert, D.; Gutiérrez, D.; Isensee, K.; et al. Declining oxygen in the global ocean and coastal waters. Science 2018, 359, eaam7240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, R.J.; Rosenberg, R. Introduction to environmental and economic consequences of hypoxia. Int. J. Water Resour. Dev. 2011, 27, 71–82. [Google Scholar] [CrossRef]
- Nixon, S.W. Coastal marine eutrophication: A definition, social causes, and future concerns. Ophelia 1995, 41, 199–219. [Google Scholar] [CrossRef]
- Zhou, F.; Chai, F.; Huang, D.; Wells, M.; Ma, X.; Meng, Q.; Xue, H.; Xuan, J.; Wang, P.; Ni, X.; et al. Coupling and decoupling of high biomass phytoplankton production and hypoxia in a highly dynamic coastal system: The Changjiang (Yangtze River) estuary. Front. Mar. Sci. 2020, 7, 259. [Google Scholar] [CrossRef]
- Whitney, M.M. Observed and projected global warming pressure on coastal hypoxia. Biogeosciences 2022, 19, 4479–4497. [Google Scholar] [CrossRef]
- Roman, M.R.; Brandt, S.B.; Houde, E.D.; Pierson, J.J. Interactive effects of hypoxia and temperature on coastal pelagic zooplankton and fish. Front. Mar. Sci. 2019, 6, 139. [Google Scholar] [CrossRef] [Green Version]
- Vaquer-Sunyer, R.; Duarte, C.M. Temperature effects on oxygen thresholds for hypoxia in marine benthic organisms. Glob. Chang. Biol. 2011, 17, 1788–1797. [Google Scholar] [CrossRef]
- Altieri, A.H.; Gedan, K.B. Climate change and dead zones. Glob. Chang. Biol. 2015, 21, 1395–1406. [Google Scholar] [CrossRef]
- Lutz, T. The magnetic reversal record is not periodic. Nature 1985, 317, 404–407. [Google Scholar] [CrossRef]
- Cande, S.C.; Kent, D.V. Revised calibration of the geomagnetic polarity timescale for the Late Cretaceous and Cenozoic. J. Geophys. Res. 1995, 100, 6093–6095. [Google Scholar] [CrossRef]
- Valet, J.-P.; Fournier, A. Deciphering records of geomagnetic reversals. Rev. Geophys. 2016, 54, 410–446. [Google Scholar] [CrossRef] [Green Version]
- Bogue, S.W.; Glen, J.M.G. Very rapid geomagnetic field change recorded by the partial remagnetization of a lava flow. Geophys. Res. Lett. 2010, 37, L21308. [Google Scholar] [CrossRef] [Green Version]
- Coe, R.S.; Prevot, M.; Camps, P. New evidence for extraordinarily rapid change of the geomagnetic field during a reversal. Nature 1995, 374, 687–692. [Google Scholar] [CrossRef]
- Hulot, G.; Lhuillier, F.; Aubert, J. Earth’s dynamo limit of predictability. Geophys. Res. Lett. 2010, 37, L06305. [Google Scholar] [CrossRef]
- Liu, Q. Will the Earth’s magnetic reverse its polarity in the near future? Solid Earth Sci. 2020, 5, 247–248. [Google Scholar] [CrossRef]
- Cooper, A.; Turney, C.S.M.; Palmer, J.; Hogg, A.; McGlone, M.; Wilmshurst, J.; Lorrey, A.M.; Heaton, T.J.; Russell, J.M.; McCracken, K.; et al. A global environmental crisis 42000 years ago. Science 2021, 371, 811–818. [Google Scholar] [CrossRef]
- Peters, R.H. Daphnia culture. Mem. Ist. Ital. Idrobiol. 1987, 45, 483–495. [Google Scholar]
- ASTM. Standard Practice for Conducting Acute Toxicity Tests with Fishes, Macroinvertebrates and Amphibians; American Standards for Testing and Materials: Philadelphia, PA, USA, 1980. [Google Scholar]
- Bikbulatov, E.S.; Bikbulatova, E.M.; Litvinov, A.S.; Poddubnyi, S.A. Gidrologiya i Gidrokhimiya Ozera Nero (Hydrology and Hydrochemistry of Lake Nero); Rybinskii Dom Pechati: Rybinsk, Russia, 2003; pp. 20–153. [Google Scholar]
- Kay, J.E.; Deser, C.; Phillips, A.; Mai, A.; Hannay, C.; Strand, G.; Arblaster, J.; Bates, S.; Danabasoglu, G.; Edwards, J.; et al. The community Earth system model (CESM) large ensemble project: A community resource for studying climate change in the presence of internal climate variability. Bull. Am. Meteorol. Soc. 2015, 96, 1333–1349. [Google Scholar] [CrossRef] [Green Version]
- OECD. OECD Guideline for Testing of Chemicals, 211: Daphnia magna Reproduction Test; Organization for Economic Cooperation and Development: Paris, France, 2012. [Google Scholar]
- Wilczynski, W.; Babkiewicz, E.; Pukos, S.; Wawrzeńczak, J.; Zebrowski, M.L.; Banasiak, L.; Kudriashov, M.; Maszczyk, P. The effects of hypoxia on threshold food concentrations in different Daphnia Species. Water 2022, 14, 3213. [Google Scholar] [CrossRef]
- Heisey, D.; Porter, K.G. The effect of ambient oxygen concentration on filtering and respiration rates of Daphnia galeata mendotae and Daphnia magna. Limnol. Oceanogr. 1977, 22, 839–845. [Google Scholar] [CrossRef] [Green Version]
- Paul, R.; Colmorgen, M.; Hüller, S.; Tyroller, F.; Zinkler, D. Circulation and respiratory control in millimetre-sized animals (Daphnia magna, Folsomia candida) studied by optical methods. J. Comp. Physiol. B 1997, 167, 399–408. [Google Scholar] [CrossRef]
- Paul, R.J.; Zeis, B.; Lamkemeyer, T.; Seidl, M.; Pirow, R. Control of oxygen transport in the microcrustacean Daphnia: Regulation of haemoglobin expression as central mechanism of adaptation to different oxygen and temperature conditions. Acta Physiol. Scand. 2004, 182, 259–275. [Google Scholar] [CrossRef] [PubMed]
- Zeis, B.; Becher, B.; Goldmann, T.; Clark, R.; Vollmer, E.; Bölke, B.; Bredebusch, I.; Lamkemeyer, T.; Pinkhaus, O.; Pirow, R.; et al. Differential haemoglobin gene expression in the crustacean Daphnia magna exposed to different oxygen partial pressures. Biol. Chem. 2003, 384, 1133–1145. [Google Scholar] [CrossRef]
- Hanazato, T.; Yasuno, M.; Hosomi, M. Significance of a low oxygen layer for a Daphnia population in lake Yunoko, Japan. Hydrobiologia 1989, 185, 19–27. [Google Scholar] [CrossRef]
- Nebeker, A.V.; Dominguez, S.E.; Chapman, G.A.; Onjukka, S.; Stevens, D.G. Effects of low dissolved oxygen on survival, growth and reproduction of Daphnia, Hyalella and Gammarus. Environ. Toxicol. Chem. 1992, 11, 373–379. [Google Scholar] [CrossRef]
- Lyu, K.; Wang, Q.; Yang, Z.; Montagnes, D.J.S. Cumulative effects matter: Multi-brood responses of Daphnia to hypoxia. Freshw. Biol. 2016, 61, 1685–1693. [Google Scholar] [CrossRef] [Green Version]
- Seidl, M.D.; Paul, R.J.; Pirow, R. Effects of hypoxia acclimation on morpho-physiological traits over three generations of Daphnia magna. J. Exp. Biol. 2005, 208, 2165–2175. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, D.; Morley, S.A.; Hughes, R.N. From cells to colonies: At what levels of body organization does the ‘temperature-size rule’ apply? Evol. Dev. 2006, 8, 202–214. [Google Scholar] [CrossRef]
- Czarnoleski, M.; Ejsmont-Karabin, J.; Angilletta, M.J.; Kozlowski, J. Colder rotifers grow larger but only in oxygenated waters. Ecosphere 2015, 6, 164. [Google Scholar] [CrossRef]
- Krylov, V.V.; Bolotovskaya, I.V.; Osipova, E.A. The response of European Daphnia magna Straus and Australian Daphnia carinata King to changes in geomagnetic field. Electromagn. Biol. Med. 2013, 32, 30–39. [Google Scholar] [CrossRef]
- Wan, G.J.; Jiang, S.L.; Zhao, Z.C.; Xu, J.J.; Tao, X.R.; Sword, G.A.; Gao, Y.B.; Pan, W.D.; Chen, F.J. Bio-effects of near-zero magnetic fields on the growth, development and reproduction of small brown planthopper, Laodelphax striatellus and brown planthopper, Nilaparvata lugens. J. Insect Physiol. 2014, 68, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Binhi, V.N.; Prato, F.S. Biological effects of the hypomagnetic field: An analytical review of experiments and theories. PLoS ONE 2017, 12, e0179340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enserink, L.; de la Haye, M.; Maas, H. Reproductive strategy of Daphnia magna: Implications for chronic toxicity tests. Aquat. Toxicol. 1993, 25, 111–123. [Google Scholar] [CrossRef]
- Burns, C.W. Effects of crowding and different food levels on growth and reproductive investment of Daphnia. Oecologia 1995, 101, 234–244. [Google Scholar] [CrossRef]
- Krylov, V.V.; Osipova, E.A. The response of Daphnia magna Straus to the long-term action of low-frequency magnetic fields. Ecotoxicol. Environ. Saf. 2013, 96, 213–219. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krylov, V.V.; Sizova, A.A.; Sizov, D.A. Effects of Hypoxia and Hypomagnetic Field on Morphometric and Life-History Traits in Freshwater Cladoceran Daphnia magna. Water 2022, 14, 3955. https://doi.org/10.3390/w14233955
Krylov VV, Sizova AA, Sizov DA. Effects of Hypoxia and Hypomagnetic Field on Morphometric and Life-History Traits in Freshwater Cladoceran Daphnia magna. Water. 2022; 14(23):3955. https://doi.org/10.3390/w14233955
Chicago/Turabian StyleKrylov, Viacheslav V., Anastasia A. Sizova, and Daniil A. Sizov. 2022. "Effects of Hypoxia and Hypomagnetic Field on Morphometric and Life-History Traits in Freshwater Cladoceran Daphnia magna" Water 14, no. 23: 3955. https://doi.org/10.3390/w14233955
APA StyleKrylov, V. V., Sizova, A. A., & Sizov, D. A. (2022). Effects of Hypoxia and Hypomagnetic Field on Morphometric and Life-History Traits in Freshwater Cladoceran Daphnia magna. Water, 14(23), 3955. https://doi.org/10.3390/w14233955