
Nitrate Water Contamination from Industrial Activities and Complete Denitrification as a Remediation Option


Abstract
:1. Introduction
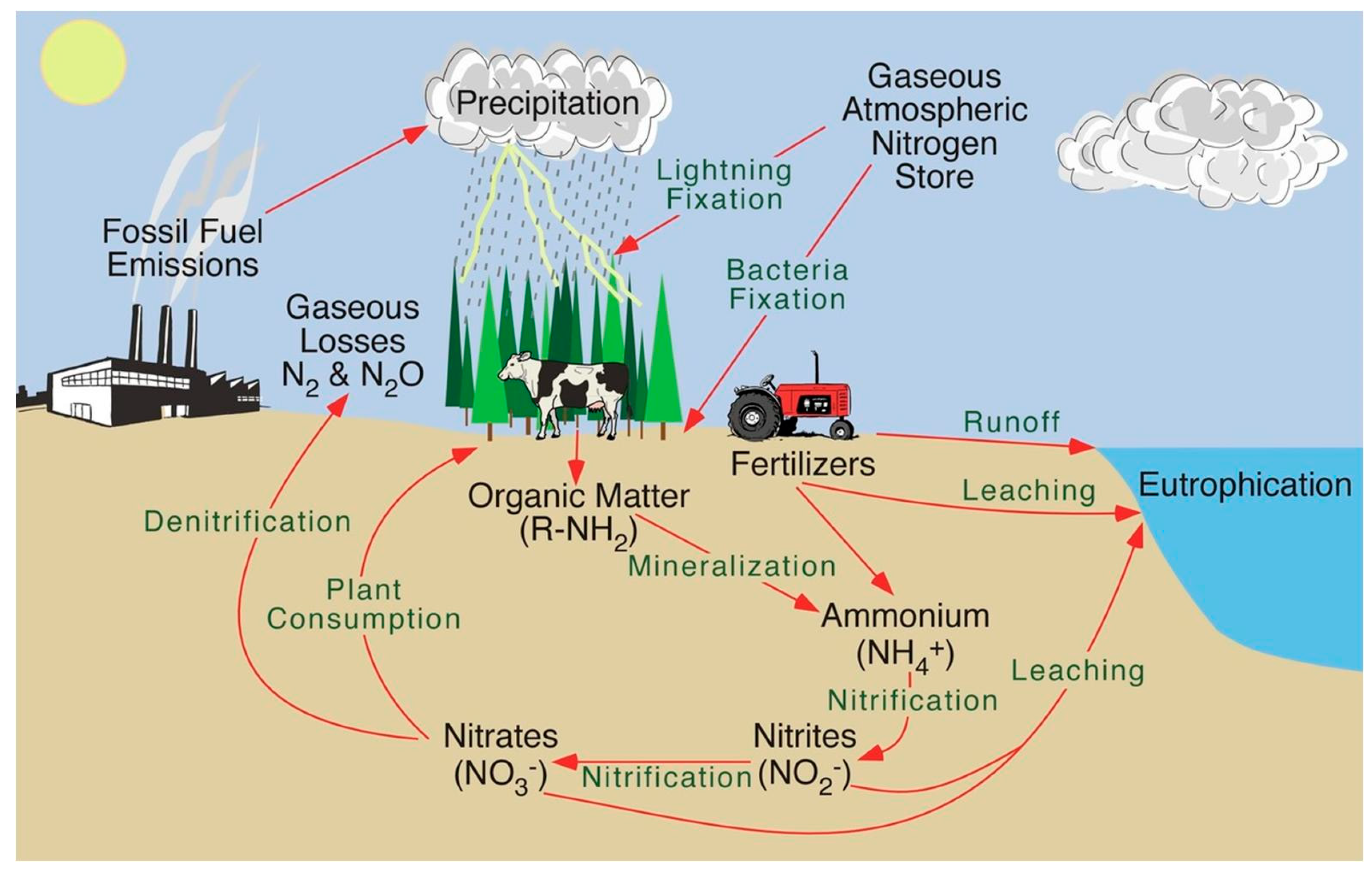
2. Biogeochemical Nitrogen Cycle
3. Sources and Effects of Nitrate Contamination in Water
3.1. Nitrate Contamination from Inorganic Fertilizers
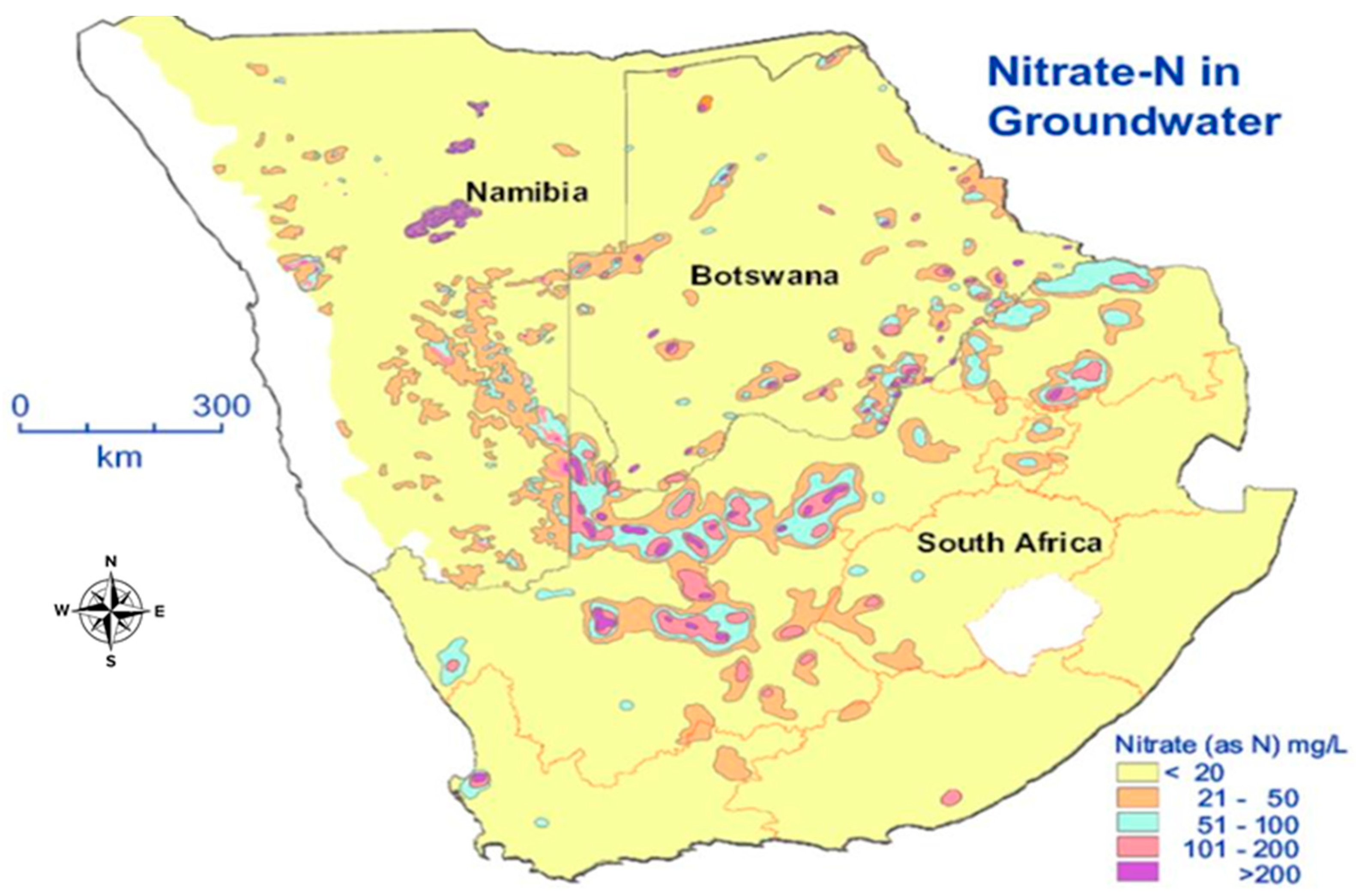
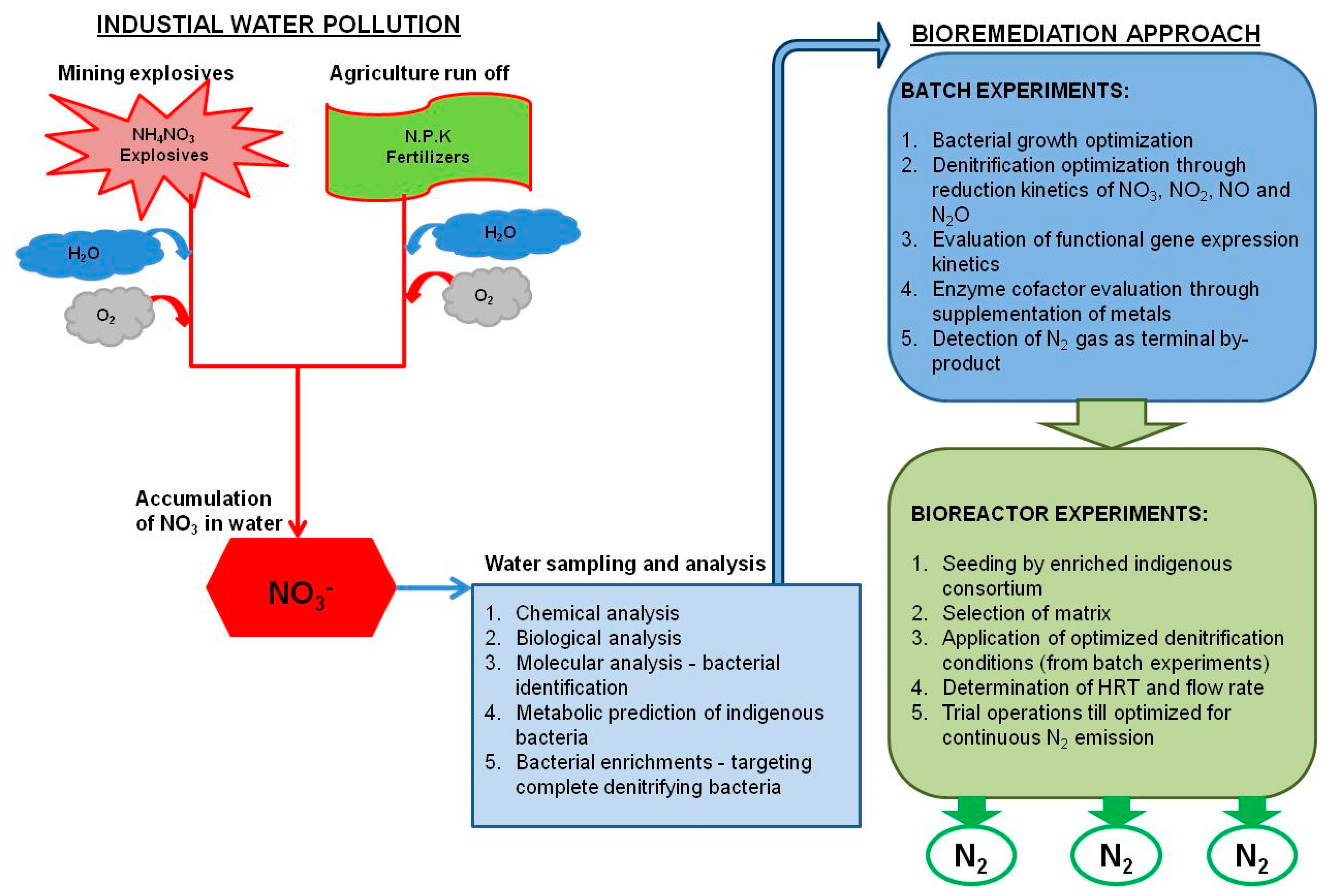
3.2. Nitrate Contamination from Mining Activities
4. Methods Applied for Mitigating Nitrate Contamination in Wastewater
4.1. Chemical Nitrate Remediation
4.2. Physical Nitrate Remediation
4.3. Bioremediation as an Option for Mitigating Nitrate Contamination
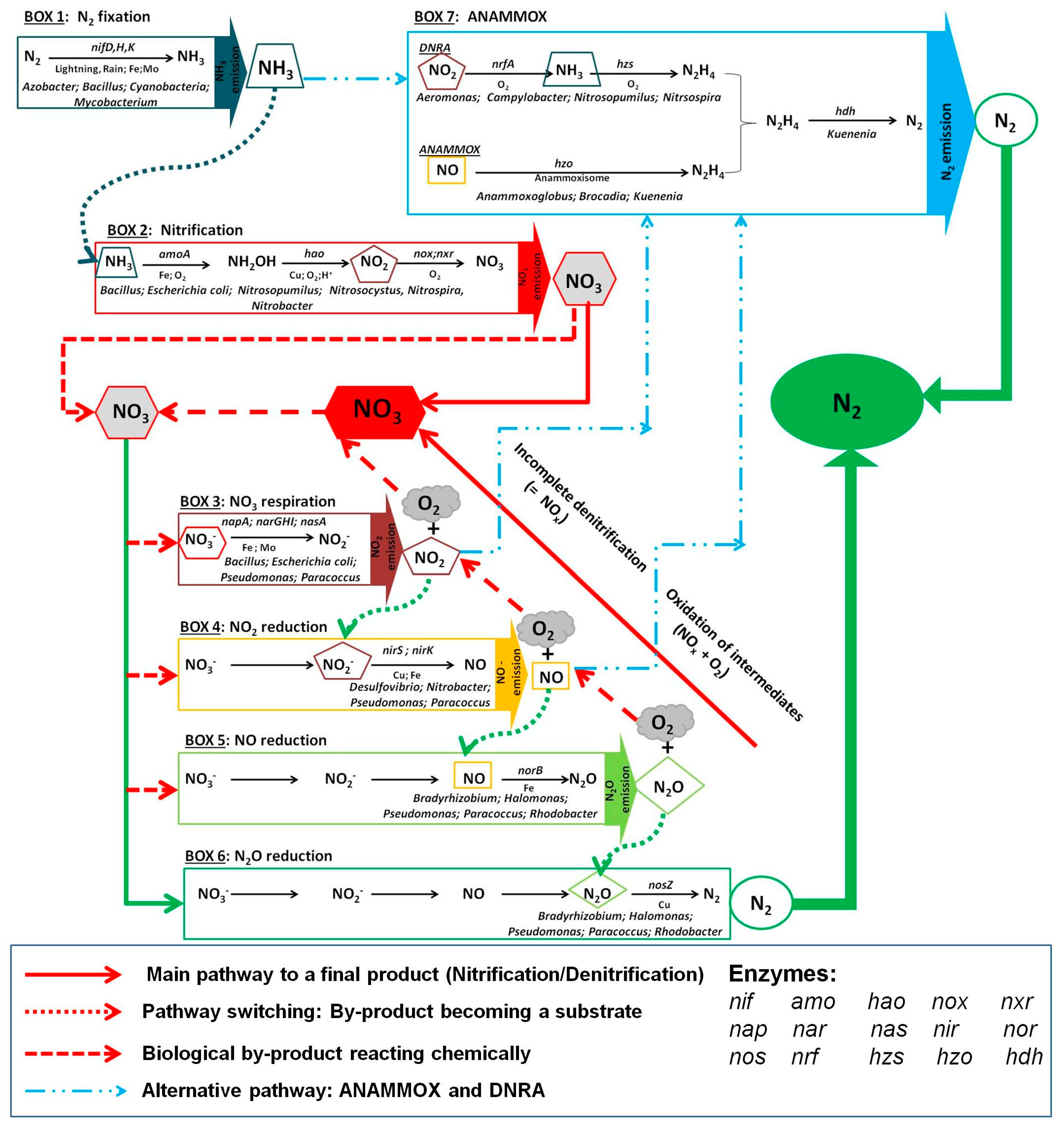
5. Microbial Nitrogen Metabolism and Biochemical Pathways
6. Application of Denitrification in Bioreactors
6.1. Use of aMatrix as a Bacterial Support in Bioreactors
6.2. Importance of SeedingBioreactor Culture
6.3. Choice of Carbon Sources in Denitrification
7. Factors That Inhibit and Promote Bacterial Denitrification
7.1. Effects of Temperature on Denitrification
7.2. Effects of pH Changes in a Denitrification System
7.3. Oxidation Reduction Potential (ORP) as a Redox Indicator in Denitrifcation
7.4. Influence of Metals on Denitrification
8. Recommendations for Optimizing Denitrification for Wastewater Remediation
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Howard, G.; Calow, R.; Macdonald, A.; Bartram, J. Climate Change and Water and Sanitation: Likely Impacts and Emerging Trends for Action. Annu. Rev. Environ. Resour. 2016, 41, 253–276. [Google Scholar] [CrossRef] [Green Version]
- Khanichaidecha, W.; Nakaruk, A.; Ratananikom, K.; Eamrat, R.; Kazama, F. Heterotrophic nitrification and aerobic denitrification using pure-culture bacteria for wastewater treatment. J. Water Reuse Desalination 2019, 9, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Santucci, L.; Carol, E.; Tanjal, C. Industrial waste as a source of surface and groundwater pollution for more than half a century in a sector of the Río de la Plata coastal plain (Argentina). Chemosphere 2018, 206, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Seckler, D.; Barker, R.; Amarasinghe, U. Water scarcity in the twenty-first century. Int. J. Water Resour. Dev. 1999, 15, 29–42. [Google Scholar] [CrossRef]
- Payus, C.; Huey, L.A.; Adnan, F.; Rimba, A.B.; Mohan, G.; Chapagain, S.K.; Roder, G.; Gasparatos, A.; Fukushi, K. Impact of extreme drought climate on water security in North Borneo: Case study of Sabah. Water 2020, 12, 1135. [Google Scholar] [CrossRef]
- Kihampa, C.; Wenaty, A. Impact if mining and farming activities on water and sediment quality of the Mara river basin, Tanzania. Res. J. Chem. Sci. 2013, 3, 15–24. [Google Scholar]
- Pimentel, D.; Berger, B.; Filiberto, D.; Newton, M.; Wolfe, B.; Karabinakis, E.; Clark, S.; Poon, E.; Abbett, E.; Nandagopal, S. Water Resources: Agricultural and Environmental Issues. BioScience 2004, 54, 909–918. [Google Scholar] [CrossRef] [Green Version]
- Fagiya, A.O.; Ipinmoroti, M.O.; Chirenje, T. Environmental pollution in Africa. Environ. Dev. Sustain. 2018, 20, 41–73. [Google Scholar] [CrossRef]
- Sparacino-Watkins, C.; Stolz, J.F.; Basu, P. Nitrate and periplasmic nitrate reductases. Chem. Soc. Rev. 2013, 43, 676–706. [Google Scholar] [CrossRef]
- Meng, Q.; Feng, Q.; Wu, Q.; Meng, L.; Cao, Z. Distribution characteristics of nitrogen and phosphorus in mining induces subsidence wetland in Panbei coal mine, China. Procedia Earth Planet. 2009, 1, 1237–1241. [Google Scholar]
- Maherry, A.; Tredoux, G.; Clarke, S.; Pengelbrecht, P. State of Nitrate Pollution in Groundwater in South Africa. Available online: http://researchspace.csir.co.za/dspace/bitstream/handle/10204/4288/Maherry_2010_P.pdf;jsessionid=C3B27245662E9D7865F7FDB24E1F19FF?sequence=1 (accessed on 17 January 2020).
- Mahvi, A.H.; Nouri, J.; Babaei, A.A.; Nabizadeh, R. Agricultural impact of groundwater nitrate pollution. Int. J. Sci. Technol. 2005, 2, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Stadler, S.; Talma, A.; Tredoux, G.; Wrabel, J. Identification of sources and infiltration regimes of nitrate in the semi-arid Kalahari: Regional differences and implications for groundwater management. Water SA 2012, 38, 213–224. [Google Scholar] [CrossRef] [Green Version]
- Negovanović, M.; Kričak, L.; Milanović, S.; Dokić, N.; Simić, N. Ammonium nitrate explosion hazards opasnosti od eksplozijekojepoticu od amonijumnitrata. Undergr. Min. Eng. 2015, 27, 49–63. [Google Scholar]
- Dwivedi, U.N.; Misha, S.; Singh, P.; Tripathi, R.D. Nitrate pollution and its remediation. In Environmental Bioremediation Technologies; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Liu, L.; Zheng, X.; Wei, X.; Kai, Z.; Xu, Y. Excessive application of chemical fertilizer and organophosphorus pesticides induced total phosphorus loss from planting causing surface water eutrophication. Sci. Rep. 2021, 11, 23015. [Google Scholar] [CrossRef]
- Agrawal, A.; Pandey, R.S.; Sharma, B. Water Pollution with Special Reference to Pesticide Contamination in India. J. Water Resour. Prot. 2010, 2, 432–448. [Google Scholar] [CrossRef] [Green Version]
- Ward, M.H.; Jones, R.R.; Brender, J.D.; de Kok, T.M.; Weyer, P.J.; Nolan, B.T.; Villanueva, C.M.; van Breda, S.G. Drinking water nitrate and human health: An updated review. Int. J. Environ. Res. Public Health 2018, 15, 1557. [Google Scholar] [CrossRef] [Green Version]
- Glass, C.; Silverstein, J. Denitrification kinetics of high nitrate concentration water: pH effect on inhibition and nitrite accumulation. Water Res. 1998, 32, 831–839. [Google Scholar] [CrossRef]
- Huno, S.K.M.; Rene, E.R.; van Hullebusch, E.D.; Annachhatre, A.P. Nitrate removal from groundwater: A review of natural and engineered processes. J. Water Supply: Res. Technol.-Aqua 2018, 67, 885–902. [Google Scholar] [CrossRef] [Green Version]
- Tagma, T.; Hsissou, Y.; Bouchaou, L.; Bouragba, L.; Boutaleb, S. Groundwater nitrate pollution in Souss-Massa basin (South-west Morocco). Afr. J. Environ. Sci. Technol. 2009, 3, 301–309. [Google Scholar]
- Feng, W.; Wang, C.; Lei, X.; Wang, H.; Zhang, X. Distribution of Nitrate Content in Groundwater and Evaluation of Potential Health Risks: A Case Study of Rural Areas in Northern China. Int. J. Environ. Res. Public Health 2020, 17, 9390. [Google Scholar] [CrossRef]
- Beutel, M.W.; Horne, A.J.; Taylor, W.D.; Losee, R.F.; Whitney, R.D. Effects of oxygen and nitrate on nutrient release from profundal sediments of a large, oligo-mesotrophic reservoir, Lake Mathews, California. Lake Reserv. Manag. 2008, 24, 18–29. [Google Scholar] [CrossRef]
- Schaider, L.A.; Swetschinki, L.; Campbell, C.; Rudel, R.A. Environmental justice and drinking water quality: Are there socioeconomic disparities in nitrate levels in U.S. drinking water? Environ. Health 2019, 18, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coronel, G.; Chang, M.; Rodríguez-Delfín, A. Nitrate Reductase Activity and Chlorophyll Content in Lettuce Plants Grown Hydroponically and Organically. In Proceedings of the International Symposium on Soilless Culture and Hydroponics 843, Lima, Peru, 25–28 August 2008; pp. 137–144. [Google Scholar]
- Coffey, M.; Pal, M.; Stamp, J.; Hamilton, A.; Johnson, T. A review of water quality responses to air temperature and precipitation changes 2: Nutrients, algal blooms, sediment, pathogens. J. Am. Water Resour. Assoc. 2018, 55, 844–868. [Google Scholar] [CrossRef] [PubMed]
- Cammack, R.; Joannou, C.; Cui, X.-Y.; Martinez, C.T.; Maraj, S.R.; Hughes, M.N. Nitrite and nitrosyl compounds in food preservation. Biochim. et Biophys. Acta 1999, 1411, 475–488. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.; Song, J.; Kong, F.; Wen, M.; Zhang, G. Nitrate pollution and preliminary source identification of surface water in a semi-arid river basin, using isotopic and hydrochemical approaches. Water 2016, 8, 328. [Google Scholar] [CrossRef]
- Panagiotaras, D.; Koulougliotis, D.; Nikolopoulos, D.; Kalarakis, A.N.; Yiannopoulos, A.C.; Pikios, K. Biogeochemical cycling of nutrients and thermodynamic aspects. J. Thermodyn. Catalysis. 2015, 6, 1–9. [Google Scholar]
- Stein, L.Y.; Klotz, M.G. The nitrogen cycle. Curr. Biol. 2016, 26, 94–98. [Google Scholar] [CrossRef] [Green Version]
- Stüeken, E.E.; Kipp, M.; Koehler, M.C.; Buick, R. The evolution of Earth’s biogeochemical nitrogen cycle. Earth-Sci. Rev. 2016, 160, 220–239. [Google Scholar] [CrossRef] [Green Version]
- Park, Z.S.; Kim, J.; Woo, Y.Y.; Lee, H.; Kim, J.H.; Moon, Y.H. Forced circulation of nitrogen gas for accelerated and eco-friendly cooling of metallic parts. Appl. Sci. 2019, 9, 3679. [Google Scholar] [CrossRef] [Green Version]
- Kuypers, M.M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef]
- Pajares, S.; Bohannan, B.J.M. Ecology of nitrogen fixing, nitrifying and denitrifying microorganisms in tropical forest soils. Front. Microbiol. 2016, 7, 1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scolow, R.H. Nitrogen management and the future of food: Lessons from the management of energy ad carbon. Proc. Natl. Acad. Sci. USA 1999, 96, 6001–6008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, W.; Liu, X.-Y.; Hu, C.-C.; Chen, G.-Y.; Walters, W.W.; Michalski, G.; Liu, C.-Q. Important contributions of non-fossil fuel nitrogen oxides emissions. Nat. Commun. 2021, 12, 243. [Google Scholar] [CrossRef] [PubMed]
- Skiba, U.M.; Rees, R.M. Nitrous Oxide, Climate Change and Agriculture; CABI: Wallingford, UK, 2014. [Google Scholar]
- Pidwirny, M. “The Nitrogen Cycle”. In Fundamentals of Physical Geography, 2nd ed.; Available online: http://www.physicalgeography.net/fundamentals/9s.html (accessed on 17 January 2022).
- Stefanakis, A.I.; Zouzias, D.; Marsellos, A. Groundwater pollution: Human and natural sources and risks. Environ. Sci. Eng. 2017, 4, 82–102. [Google Scholar]
- Withers, P.J.A.; Neal, C.; Jarvie, H.P.; Doody, D.G. Agriculture and Eutrophication: Where Do We Go from Here? Sustainability 2014, 6, 5853–5875. [Google Scholar] [CrossRef] [Green Version]
- Bosman, C. The hidden dragon: Nitrate pollution from open-pit mines—A case study from the Limpopo province, South Africa. In Proceedings of the International Mine Water Conference, Pretoria, South Africa, 19–23 October 2009. [Google Scholar]
- Cyplik, P.; Marecik, R.; Piotrowska-Cyplik, A.; Olejnik, A.; Drozdzynska, A.; Chrzanowski, L. Biological denitrification of high nitrate processing wastewaters from explosives production plant. Water Air Soil Pollut. 2012, 223, 1791–1800. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; He, R.; Han, W.; Sun, X.; Li, J.; Wang, L. Biological denitrification of high-nitrate wastewater in a modified anoxic/oxic-membrane bioreactor (A/O-MBR). J. Hazard. Mater. 2009, 172, 595–600. [Google Scholar] [CrossRef]
- Soomro, F.; Rafique, T.; Michalski, G.; Ali, S.A.; Naseem, S.; Khan, M.U. Occurrence and delineation of high nitrate contamination in the groundwater of Mithi sub-district, Thar Desert, Pakistan. Environ. Earth Sci. 2017, 76, 355. [Google Scholar] [CrossRef]
- Alex, R.; Kitalika, A.; Mogusu, E.; Njau, K. Sources of Nitrate in Ground Water Aquifers of the Semiarid Region of Tanzania. Geofluids 2021, 2021, 6673013. [Google Scholar] [CrossRef]
- Tredoux, G.; Engelbrecht, P.; Israel, S. Nitrate in Groundwater. In Why Is It a Hazard and How to Control It? Report to Water Research Commission; Water Research Commission: Pretoria, South Africa, 2009; pp. 1–22. ISBN 978-1-77005-873-6. [Google Scholar]
- Zendehbad, M.; Cepuder, P.; Loiskandl, W.; Stumpp, C. Source identification of nitrate contamination in the urban aquifer of Mashhad, Iran. J. Hydrol. Reg. Stud. 2019, 25, 100618. [Google Scholar] [CrossRef]
- Laursen, A.E.; Carlton, R.G. Response to atrazine of respiration, nitrification, and denitrification in stream sediments measured with oxygen and nitrate microelectrodes. FEMS Microbiol. Ecol. 1999, 29, 229–240. [Google Scholar] [CrossRef]
- Lee, C.; Beauchemin, K.A. A review of feeding supplementary nitrate to ruminant animals: Nitrate toxicity, methane emissions, and production performance. Can. J. Anim. Sci. 2014, 94, 557–570. [Google Scholar] [CrossRef]
- Avisar, O.; Kronfeld, J.; Talma, A.S. Amelioration of groundwater nitrate contamination following installation of a central sewage system in two Israeli villages. Environ. Earth Sci. 2008, 58, 515–520. [Google Scholar] [CrossRef]
- Verlicchi, P.; Grillini, V. Surface Water and Groundwater Quality in South Africa and Mozambique—Analysis of the Most Critical Pollutants for Drinking Purposes and Challenges in Water Treatment Selection. Water 2020, 12, 305. [Google Scholar] [CrossRef] [Green Version]
- Schoeman, J.J. Nitrate-nitrogen removal with small-scale reverse osmosis, electrodialysis and ion-exchange units in rural areas. Water SA 2009, 35, 5. [Google Scholar] [CrossRef] [Green Version]
- Amatya, I.M.; Kansakar, E.B.; Tare, E.V.; Fiksdal, L. Impact of temperature on biological denitrification process. J. Inst. Eng. 2015, 7, 121–126. [Google Scholar] [CrossRef] [Green Version]
- Clark, C.M.; Bell, M.D.; Boyd, J.W.; Compton, J.E.; Davidson, E.A.; Davis, C.; Fenn, M.E.; Geiser, L.; Jones, L.; Blett, T.F. Nitrogen-induced terrestrial eutrophication: Cascading effects and impacts on ecosystems services. Ecosphere 2017, 8, e01877. [Google Scholar] [CrossRef] [Green Version]
- Glibert, P.M.; Harrison, J.; Heil, C.; Seitzinger, S. Escalating Worldwide use of Urea—A Global Change Contributing to Coastal Eutrophication. Biogeochemistry 2006, 77, 441–463. [Google Scholar] [CrossRef]
- Basada, H.; Werner, K. An assessment of the effects of Africa’s water crisis on food security and management. Int. J. Water Resour. Dev. 2015, 3, 120–133. [Google Scholar] [CrossRef] [Green Version]
- Rahman, K.M.A.; Zhang, D. Effects of fertilizer broadcasting on the excessive use of inorganic fertilizers and environmental sustainability. Sustainability 2018, 10, 759. [Google Scholar] [CrossRef] [Green Version]
- Yousaf, M.; Li, J.; Lu, J.; Ren, T.; Cong, R.; Fahad, S.; Li, X. Effects of fertilization on crop production and nutrient-supplying capacity under rice-oilseed rape rotation system. Sci. Rep. 2017, 7, 1270. [Google Scholar] [CrossRef] [PubMed]
- Zavale, H.; Matchaya, G.; Vilissa, D.; Nhemachena, C.; Nhlengethwa, S.; Wilson, D. Dynamics of the Fertilizer Value Chain in Mozambique. Sustainability 2020, 12, 4691. [Google Scholar] [CrossRef]
- Heffer, P.; Gruere, A.; Roberts, T. Assessment of Fertilizer Use by Crop at the Global Level. Available online: https://www.fertilizer.org/images/library_downloads/agcom.13.39%20-%20fubc%20assessment%202010.pdf (accessed on 17 January 2022).
- Li, Z.; Zhang, R.; Xia, S.; Wang, L.; Liu, C.; Zhang, R.; Fan, Z.; Chen, F.; Liu, Y. Interactions between N, P and K fertilizers affect the environment and the yield and quality of satsumas. Glob. Ecol. Conserv. 2019, 19, e00663. [Google Scholar] [CrossRef]
- Kochakinezhad, H.; Peyvast, G.; Kashi, A.K.; Olfati, J.A.; Asadii, A. A comparison of organic and chemical fertilizers for tomato production. J. Org. Syst. 2012, 7, 14–25. [Google Scholar]
- Liu, C.-W.; Sung, Y.; Chen, B.-C.; Lai, H.-Y. Effects of Nitrogen Fertilizers on the Growth and Nitrate Content of Lettuce (Lactuca sativa L.). Int. J. Environ. Res. Public Health 2014, 11, 4427–4440. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.N.; Mobin, M.; Abbas, Z.K.; Alamri, S.A. Fertilizers and their contaminants in soils, surface and groundwater. Encycl. Anthr. 2018, 5, 225–240. [Google Scholar]
- Fuadi, M.; Sutiarso, L.; Radi; Virgawati, S.; Nugraheni, P.H.T. Design of liquid fertilizer applicator based on Variable Rate Application (VRA) for Soybean. IOP Conf. Ser. Earth Environ. Sci. 2019, 355, 012009. [Google Scholar] [CrossRef]
- Damashek, J.; Francis, C.A. Microbial nitrogen cycling in estuaries: From genes to ecosystems. Estuaries Coasts 2018, 41, 626–660. [Google Scholar] [CrossRef]
- Hirel, B.; Tetu, T.; Lea, P.J.; Dubois, F. Improving Nitrogen Use Efficiency in Crops for Sustainable Agriculture. Sustainability 2011, 3, 1452–1485. [Google Scholar] [CrossRef]
- Ye, L.; Zhao, X.; Bao, E.; Li, J.; Zou, Z.; Cao, K. Bio-organic fertilizer with reduced rates of chemical fertilization improves soil fertility and enhances tomato yield and quality. Sci. Rep. 2020, 10, 177. [Google Scholar] [CrossRef] [Green Version]
- Hasnain, M.; Chen, J.; Ahmed, N.; Memon, S.; Wang, L.; Wang, Y.; Wang, P. The Effects of Fertilizer Type and Application Time on Soil Properties, Plant Traits, Yield and Quality of Tomato. Sustainability 2020, 12, 9065. [Google Scholar] [CrossRef]
- Kakar, K.; Xuan, T.D.; Noori, Z.; Aryan, S.; Gulab, G. Effects of Organic and Inorganic Fertilizer Application on Growth, Yield, and Grain Quality of Rice. Agriculture 2020, 10, 544. [Google Scholar] [CrossRef]
- Carvalho, F.P. Mining industry and sustainable development: Time for change. Food Energy Secur. 2017, 6, 61–77. [Google Scholar] [CrossRef]
- Ngcofe, L.; Cole, D.I. The distribution of the economic mineral resource potential in the Western Cape Province. S. Afr. J. Sci. 2014, 110, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Africa Mining IQ (2019). Available online: www.projectsiq.co.za (accessed on 17 January 2022).
- Mattila, K.; Zaitsev, G.; Langwaldt, J. Biological removal of nutrients from mine waters. In Biologinen ravinteiden poisto kaivosvedestä; Finnish Forest Research Institute: Rovaniemi, Finland, 2007. [Google Scholar]
- Suceska, M.; Tumara, B.S.; Skrlec, V.; Stankovic, S. Prediction of concentration of toxic gases produced by detonation of commercial explosives by thermochemical equilibrium calculations. Def. Technol. 2021. [Google Scholar] [CrossRef]
- Mikić, M.; Rajković, R.; Jovanović, M.; Maksimović, M. The impact of blasting on the environment in the open pit mining. Min. Metall. Inst. Bor 2017, 3–4, 165–170. [Google Scholar] [CrossRef]
- Crosson, C.C.; Tonking, M.J.H.; Paterson, G. The development of drilling and blasting practice at Palaborwa mining company limited. J. S. Afr. Inst. Min. Metall. 1976, 76, 325–340. [Google Scholar]
- Suppajari, P.; Elie, M.; Baron, M.; Gonzalez-Rodriguez, J. Classification of ANFO samples based on their fuel composition by GC–MS and FTIR combined with chemometrics. Forensic Sci. Int. 2019, 301, 415–425. [Google Scholar] [CrossRef]
- Brochu, S. Assessment of ANFO on the Environment; Technical Investigation; Defence Research and Development Canada Valcartier (QUEBEC): Quebec, QC, Canada, 2010. [Google Scholar]
- Biessikirski, A.; Kuterasinski, L.; Dworzak, M.; Pyra, J.; Twardosz, M. Comparison of structure, morphology, and topography of fertilizer-based explosives applied in the mining industry. Microchem. J. 2019, 144, 39–44. [Google Scholar] [CrossRef]
- Turcotte, R.; Badeen, C.M.; Goldthorp, S. Minimum Burning Pressures of Water-based Emulsion Explosives. J. Vis. Exp. 2017, e56167. [Google Scholar] [CrossRef]
- Ignarro, L.J. After 130 years, the molecular mechanism of action of nitroglycerin is revealed. Proc. Natl. Acad. Sci. USA 2002, 99, 7816–7817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purwantini, E.; Mukhopadhyay, B. Conversion of NO2 to NO by reduced coenzyme F420 protects mycobacteria from nitrosative damage. Proc. Natl. Acad. Sci. USA 2009, 106, 6333–6338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, M.; Khare, P.; Chakravarty, S.; Saikia, D.; Saikia, B.K. Physico-chemical and elemental investigation of aqueous leaching of high sulfur coal and mine overburden from Ledo coalfield of Northeast India. Int. J. Coal Sci. Technol. 2018, 5, 265–281. [Google Scholar] [CrossRef] [Green Version]
- Dold, B. Evolution of Acid Mine Drainage Formation in Sulphidic Mine Tailings. Minerals 2014, 4, 621–641. [Google Scholar] [CrossRef] [Green Version]
- Wei, W.; Ma, R.; Sun, Z.; Zhou, A.; Bu, J.; Long, X.; Liu, Y. Effects of Mining Activities on the Release of Heavy Metals (HMs) in a Typical Mountain Headwater Region, the Qinghai-Tibet Plateau in China. Int. J. Environ. Res. Public Health 2018, 15, 1987. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.; Sinha, A. Review on treatment of acid mine drainage with waste materials: A novel approach. Glob. NEST J. 2018, 20, 512–528. [Google Scholar]
- Song, X.; Yang, X.; Hallerman, E.; Jiang, Y.; Huang, Z. Effects of Hydraulic Retention Time and Influent Nitrate-N Concentration on Nitrogen Removal and the Microbial Community of an Aerobic Denitrification Reactor Treating Recirculating Marine Aquaculture System Effluent. Water 2020, 12, 650. [Google Scholar] [CrossRef] [Green Version]
- Sangwan, S.; Dukare, A. Microbe-mediated bioremediation: An Eco-friendly sustainable approach for environmental clean-up. In Advances in Soil Microbiology: Recent Trends and Future Prospects; Springer: Singapore, 2018. [Google Scholar] [CrossRef]
- Wang, J.; Chu, L. Biological nitrate removal from water and wastewater by solid-phase denitrification process. Biotechnol. Adv. 2016, 34, 1103–1112. [Google Scholar] [CrossRef]
- Murphy, A.P. Chemical removal of nitrate from water. Nature 1991, 350, 223–225. [Google Scholar] [CrossRef]
- Gologan, D.; Popescu, M.E. The optimization of coagulation-flocculation process in a conventional treatment water plant. IORS J. Environ. Sci. Toxicol. Food Technol. 2016, 10, 33–39. [Google Scholar] [CrossRef]
- Archna; Sharma, S.K.; Sobti, R.C. Nitrate removal from ground water: A review. Environ. J. Chem. 2012, 9, 154616. [Google Scholar] [CrossRef]
- Christianson, L.E.; Schipper, L. Moving Denitrifying Bioreactors beyond Proof of Concept: Introduction to the Special Section. J. Environ. Qual. 2016, 45, 757–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, T.; Christianson, L. Performance of Denitrifying Bioreactors at Reducing Agricultural Nitrogen Pollution in a Humid Subtropical Coastal Plain Climate. Water 2017, 9, 112. [Google Scholar] [CrossRef] [Green Version]
- Widory, D.; Petelet-Giraud, E.; Negrel, P.; Ladouche, B. Tracking the sources of nitrate in groundwater using coupled nitrogen and boron isotopes: A synthesis. Environ. Sci. Technol. 2005, 39, 539–548. [Google Scholar] [CrossRef]
- Petrounias, P.; Rogkala, A.; Giannakopoulou, P.P.; Tsikouras, B.; Lampropoulou, P.; Kalaitzidis, S.; Hatzipanagiotou, K.; Lambrakis, N.; Christopoulou, M.A. An Experimental Study for the Remediation of Industrial Waste Water Using a Combination of Low Cost Mineral Raw Materials. Minerals 2019, 9, 207. [Google Scholar] [CrossRef] [Green Version]
- Tugaoen, H.O.; Garcia-Segura, S.; Hristovski, K.; Westerhoff, P. Challenges in photocatalytic reduction of nitrate as a water treatment technology. Sci. Total Environ. 2017, 599–600, 1524–1551. [Google Scholar] [CrossRef]
- Liu, G.; You, S.; Ma, M.; Huang, H.; Ren, N. Removal of nitrate by photocatalytic denitrification using nonlinear optical material. Environ. Sci. Technol. 2016, 50, 11218–11225. [Google Scholar] [CrossRef]
- Shrimali, M.; Singh, K. New methods of nitrate removal from water. Environ. Pollut. 2001, 112, 351–359. [Google Scholar] [CrossRef]
- Santos, A.S.G.G.; Restivo, J.; Orge, C.A.; Pereira, M.F.R.; Soares, O.S.G.P. Nitrate Catalytic Reduction over Bimetallic Catalysts: Catalyst Optimization. C 2020, 6, 78. [Google Scholar] [CrossRef]
- Tokazhanov, G.; Ramazanova, E.; Hamid, S.; Bae, S.; Lee, W. Advances in the catalytic reduction of nitrate by metallic catalysts for high efficiency and N2 selectivity: A review. Chem. Eng. J. 2019, 384, 123252. [Google Scholar] [CrossRef]
- Werth, C.J.; Yan, C.; Troutman, J.P. Factors Impeding Replacement of Ion Exchange with (Electro)Catalytic Treatment for Nitrate Removal from Drinking Water. ACS EST Engg. 2020, 1, 6–20. [Google Scholar] [CrossRef]
- González-Martínez, A.; de Simón-Martín, M.; López, R.; Táboas-Fernández, R.; Bernardo-Sánchez, A. Remediation of Potential Toxic Elements from Wastes and Soils: Analysis and Energy Prospects. Sustainability 2019, 11, 3307. [Google Scholar] [CrossRef] [Green Version]
- McAdam, E.; Judd, S. A review of membrane bioreactor potential for nitrate removal from drinking water. Desalination 2006, 196, 135–148. [Google Scholar] [CrossRef]
- Park, I.-K.; Hou, J.; Yun, J.; Lee, H.-D.; Lee, C.-H. A Study to Enhance the Nitrate-Nitrogen Removal Rate without Dismantling the NF Module by Building a PFSA Ionomer-Coated NF Module. Membranes 2021, 11, 769. [Google Scholar] [CrossRef]
- Mahvi, A.H.; Malakootian, M.; Fataheizadeh, A.; Ehrampoish, M.H. Nitrate removal from aqueous solutions by nanofiltration. Desalination Water Treat. 2017, 29, 326–330. [Google Scholar] [CrossRef]
- Mogheir, Y.K.H.; Albahnasawi, A.M.; Abuhabib, A.A. Optimal use of nanofiltration for nitrate removal from Gaza strip municipal wells. IUG J. Nat. Eng. Stud. 2014, 22, 45–60. [Google Scholar]
- Hoinkis, J.; Valero-Fretag, S.; Caporgno, M.P.; Patzold., C. Removal of nitrate and fluoride by nanofiltration—A comparative study. Desalination Water Treat. 2011, 30, 278–288. [Google Scholar] [CrossRef]
- Zou, L.; Zhang, S.; Liu, J.; Cao, Y.; Qian, G.; Li, Y.-Y.; Xu, Z.P. Nitrate removal from groundwater using negatively charged nanofiltration membrane. Environ. Sci. Pollut. Res. 2018, 26, 34197–34204. [Google Scholar] [CrossRef]
- Bottas, A.; Gaidoumi, A.E.; Ksakas, A.; Kherbeche, A. Adsorption study for the removal of nitrate from water using local clay. Sci. World J. 2019, 2019, 9529618. [Google Scholar] [CrossRef] [Green Version]
- Naidu, L.D.; Saravanan, S.; Chidambaram, M.; Goel, M.; DAS, A.; Babu, J.S.C. Nanofiltration in Transforming Surface Water into Healthy Water: Comparison with Reverse Osmosis. J. Chem. 2015, 2015, 26869. [Google Scholar] [CrossRef]
- Kurama, H.; Poetzschke, J.; Haseneder, R. The application of membrane filtration for the removal of ammonium ions from potable water. Water Res. 2002, 36, 2905–2909. [Google Scholar] [CrossRef]
- Wu, J.; Cao, M.; Tong, D.; Finkelstein, Z.; Hoek, E.M.V. A critical review of point-of-use drinking water treatment in the United States. npj Clean Water 2021, 4, 40. [Google Scholar] [CrossRef]
- Mohseni-Bandpi, A.; Elliott, D.J.; Zazouli, M.A. Biological nitrate removal processes from drinking water supply—A review. J. Environ. Health Sci. Eng. 2013, 11, 35. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Ye, Q.; Sun, Q.; Cai, X.; Ni, J.; Li, Z.; Xie, D. Removal of Nitrate in Simulated Water at Low Temperature by a Novel Psychrotrophic and Aerobic Bacterium, Pseudomonas taiwanensis Strain J. BioMed Res. Int. 2018, 2018, 4984087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarty, P.L. What is the best biological process for nitrogen removal: When and why? Environ. Sci. Technol. 2018, 52, 3835–3841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamp, A.; Hogslund, S.; Risgaard-Petersen, N.; Stief, P. Nitrate storage and dissimilatory nitrate reduction by eukaryotic microbes. Front. Microbiol. 2015, 6, 1492. [Google Scholar] [CrossRef] [Green Version]
- Nogales, B.; Timmis, K.N.; Nedwell, D.B.; Osborn, A.M. Detection and Diversity of Expressed Denitrification Genes in Estuarine Sediments after Reverse Transcription-PCR Amplification from mRNA. Appl. Environ. Microbiol. 2002, 68, 5017–5025. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Weisener, C.G.; Ni, J.; He, B.; Xie, D.; Li, Z. Nitrate assimilation, dissimilatory nitrate reduction to ammonium, and denitrification coexist in Pseudomonas putida Y-9 under aerobic conditions. Bioresour. Technol. 2020, 312, 123597. [Google Scholar] [CrossRef]
- Kinidi, L.; Salleh, S. Phytoremediation of Nitrogen as Green Chemistry for Wastewater Treatment System. Int. J. Chem. Eng. 2017, 2017, 1961205. [Google Scholar] [CrossRef]
- Sundaralingam, T.; Gnanavelrajah, N. Phytoremediation Potential of Selected Plants for Nitrate and Phosphorus from Ground Water. Int. J. Phytoremediation 2013, 16, 275–284. [Google Scholar] [CrossRef]
- Shyamala, S.; Manikandan, N.A.; Pakshirajan, K.; Tang, V.T.; Rene, E.R.; Park, H.S.; Behera, S.K. Phytoremediation of nitrate contaminated water using ornamental plants. J. Water Supply Res. Technol.-Aqua 2019, 68, 731–743. [Google Scholar] [CrossRef]
- Li, Y.; Luo, J.; Yu, J.; Xia, L.; Zhou, C.; Cai, L.; Ma, X. Improvement of the phytoremediation efficiency of Neyraudiareynaudiana for lead-zinc mine-contaminated soil under the interactive effect of earthworms and EDTA. Sci. Rep. 2018, 8, 6417. [Google Scholar] [CrossRef] [PubMed]
- Rezvani, F.; Sarrafzadeh, M.-H.; Ebrahimi, S.; Oh, H.-E. Nitrate removal from drinking water with a focus on biological methods: A review. Environ. Sci. Pollut. Res. 2017, 26, 1124–1141. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Qiu, T.-L.; Han, M.-L.; Li, J.; Wang, X. Heterotrophic Denitrification of Nitrate-Contaminated Water Using Different Solid Carbon Sources. Procedia Environ. Sci. 2011, 10, 72–77. [Google Scholar] [CrossRef] [Green Version]
- Rajta, A.; Bhatia, R.; Setia, H.; Pathania, P. Role of heterotrophic aerobic denitrifying bacteria in nitrate removal from wastewater. J. Appl. Microbiol. 2019, 128, 1261–1278. [Google Scholar] [CrossRef]
- Zhou, W.; Sun, Y.; Wu, B.; Zhang, Y.; Huang, M.; Miyanaga, T.; Zhang, Z. Autotrophic denitrification for nitrate and nitrite removal using sulfur-limestone. J. Environ. Sci. 2021, 23, 1761–1769. [Google Scholar] [CrossRef]
- Kim, S.; Jung, H.; Kim, K.S.; Kim, I.S. Treatment of High Nitrate-Containing Wastewaters by Sequential Heterotrophic and Autotrophic Denitrification. J. Environ. Eng. 2004, 130, 1475–1480. [Google Scholar] [CrossRef]
- Liu, F.; Wang, S.; Zhang, X.; Qian, F.; Wang, Y.; Yin, Y. Nitrate Removal from Actual Wastewater by Coupling Sulfur-Based Autotrophic and Heterotrophic Denitrification under Different Influent Concentrations. Water 2021, 13, 2913. [Google Scholar] [CrossRef]
- Casella, S.; Payne, W.J. Potential of denitrifiers for soil environment protection. FEMS Microbiol. Lett. 1996, 140, 1–8. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Wang, W.; Sun, Z.; Li, J. Investigation of growth kinetics and partial denitrification performance in strain Acinetobacter johnsonii under different environmental conditions. R. Soc. Open Sci. 2019, 6, 191275. [Google Scholar] [CrossRef] [Green Version]
- Zehr, J.P.; Ward, B.B. Nitrogen Cycling in the Ocean: New Perspectives on Processes and Paradigms. Appl. Environ. Microbiol. 2002, 68, 1015–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Rodríguez, A.J.; Rhen, M.; Melican, K.; Richter-Dahlfors, A. Nitrate Metabolism Modulates Biosynthesis of Biofilm Components in Uropathogenic Escherichia coli and Acts as a Fitness Factor during Experimental Urinary Tract Infection. Front. Micrbiol. 2020, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Vivián, C.; Cabello, P.; Martínez-Luque, M.; Blasco, R.; Castillo, F. Prokaryotic nitrate reduction: Molecular properties and functional distinction among bacterial nitrate reductases. J. Bacteriol. 1999, 181, 6573–6584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zumft, W.G. Cell biology and molecular basis of denitrification. Microbiol. Mol. Biol. Rev. 1997, 61, 533–616. [Google Scholar] [PubMed]
- Line, L.; Ealhede, M.; Kolpen, M.; Kühl, M.; Ciofu, O.; Bjarnsholt, T.; Moser, C.; Toyofuku, M.; Nomura, N.; Høiby, N.; et al. Physiological levels of nitrate support anoxic growth by denitrification of Pseudomonas aeruginosa at growth rates reported in cystic fibrosis lungs and sputum. Front. Microbiol. 2014, 5, 554. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Jiang, J.; Lv, Q.; Yang, B.; Zheng, M.; Gao, X.; Han, J.; Zhang, Y.; Yang, Y. Denitrification performance of Pseudomonas fluorescens Z03 immobilized by graphene oxide-modified polyvinyl-alcohol and sodium alginate gel beads at low temperature. R. Soc. Open Sci. 2020, 7, 191542. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Jiang, Y.; Chang, Z.; Wang, J.; Song, X.; Huang, Z.; Chen, S.; Li, J. Denitrification characteristics and pathways of a facultative anaerobic denitrifying strain, Pseudomonas denitrificans G1. J. Biosci. Bioeng. 2020, 129, 715–722. [Google Scholar] [CrossRef]
- Dalsgaard, T.; Stewart, F.J.; Thamdrup, B.; De Brabandere, L.; Revsbech, N.P.; Ulloa, O.; Canfield, D.E.; DeLong, E.F. Oxygen at Nanomolar Levels Reversibly Suppresses Process Rates and Gene Expression in Anammox and Denitrification in the Oxygen Minimum Zone off Northern Chile. mBio 2014, 5, e01966-14. [Google Scholar] [CrossRef] [Green Version]
- Foglar, L.; Briški, F.; Sipos, L.; Vuković, M. High nitrate removal from synthetic wastewater with the mixed bacterial culture. Bioresour. Technol. 2005, 96, 879–888. [Google Scholar] [CrossRef]
- Heylen, K.; Vanparys, B.; Wittebolle, L.; Verstraete, W.; Boon, N.; De Vos, P. Cultivation of Denitrifying Bacteria: Optimization of Isolation Conditions and Diversity Study. Appl. Environ. Microbiol. 2006, 72, 2637–2643. [Google Scholar] [CrossRef] [Green Version]
- Erbanová, E.; Palarčík, J.; Slezák, M.; Mikulášek, P. Removing of nitrates from waste water by using pond culture. Procedia Eng. 2012, 42, 1552–1560. [Google Scholar] [CrossRef] [Green Version]
- Larose, C.; Dommergue, A.; Vogel, T.M. Microbial nitrogen cycling in Arctic snowpacks. Environ. Res. Lett. 2013, 8, 035004. [Google Scholar] [CrossRef]
- Ward, B.B.; Jensen, M.M. The microbial nitrogen cycle. Front. Microbiol. 2014, 5, 553. [Google Scholar] [CrossRef] [Green Version]
- Strohm, T.O.; Griffin, B.; Zumft, W.G.; adSchink, B. Growth yields in bacterial denitrification and nitrate ammonification. Appl. Environ. Microbiol. 2007, 73, 1420–1424. [Google Scholar] [CrossRef] [Green Version]
- Marietou, A. Nitrate reduction in sulfate-reducing bacteria. FEMS Microbiol. Lett. 2016, 363, fnw155. [Google Scholar] [CrossRef] [PubMed]
- Cole, J. Nitrate reduction to ammonia by enteric bacteria: Redundancy, or a strategy for survival during oxygen starvation. FEMS Microbiol. Lett. 1996, 136, 1–11. [Google Scholar] [CrossRef]
- Van der Hoek, J.P.; Duijff, R.; Reinstra, O. Nitrogen Recovery from Wastewater: Possibilities, Competition with Other Resources, and Adaptation Pathways. Sustainability 2018, 10, 4605. [Google Scholar] [CrossRef] [Green Version]
- Palacin-Lizarbe, C.; Camarero, L.; Catalan, J. Denitrification temperature dependence in remote, cold, and N-poor lake sediments. Water Resour. Res. 2018, 54, 1161–1173. [Google Scholar] [CrossRef]
- Rodziewicz, J.; Ostrowska, K.; Janczukowicz, W.; Mielcarek, A. Effectiveness of Nitrification and Denitrification Processes in Biofilters Treating Wastewater from De-Icing Airport Runways. Water 2019, 11, 630. [Google Scholar] [CrossRef] [Green Version]
- Thakur, I.S.; Medhi, K. Nitrification and denitrification processes for mitigation of nitrous oxide from waste water treatment plants for biovalorization: Challenges and opportunities. Bioresour. Technol. 2019, 282, 502–513. [Google Scholar] [CrossRef]
- Yin, Z.; Bi, X.; Xu, C. Ammonia-oxidizing archaea (AOA) play with ammonia-oxidizing bacteria (AOB) in nitrogen removal from wastewater. Archaea 2018, 2018, 8429145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daims, H.; Wagner, M. Nitrospira. Trends Microbiol. 2018, 26, 462–463. [Google Scholar] [CrossRef] [PubMed]
- Subbarao, G.V.; Ito, O.; Sahrawat, K.L.; Berry, W.L.; Nakahara, K.; Ishikawa, T.; Watanabe, T.; Suenaga, K.; Rondon, M.; Rao, I.M. Scope and Strategies for Regulation of Nitrification in Agricultural Systems—Challenges and Opportunities. Crit. Rev. Plant Sci. 2006, 25, 303–335. [Google Scholar] [CrossRef] [Green Version]
- Zhu, T.; Meng, T.; Zhang, J.; Zhong, W.; Muller, C.; Cai, Z. Fungi-dominant heterotrophic nitrification in a subtropical forest soil of China. J. Soil Sediments 2015, 15, 705–709. [Google Scholar] [CrossRef]
- Karl, D.M.; Michaels, A.F. Nitrogen cycle. In Encyclopedia of Ocean Sciences; Elsevier: Amsterdam, The Netherlands, 2001. [Google Scholar] [CrossRef]
- Yoon, S.; Sanford, R.A.; Löffler, F.E. Nitrite Control over Dissimilatory Nitrate/Nitrite Reduction Pathways in Shewanellaloihica Strain PV-4. Appl. Environ. Microbiol. 2015, 81, 3510–3517. [Google Scholar] [CrossRef] [Green Version]
- Braker, G.; Zhou, J.; Wu, L.; Devol, A.H.; Tiedje, J.M. Nitrite Reductase Genes (nirK and nirS) as Functional Markers to Investigate Diversity of Denitrifying Bacteria in Pacific Northwest Marine Sediment Communities. Appl. Environ. Microbiol. 2000, 66, 2096–2104. [Google Scholar] [CrossRef] [Green Version]
- Priemé, A.; Braker, G.; Tiedje, J.M. Diversity of Nitrite Reductase (nirK and nirS) Gene Fragments in Forested Upland and Wetland Soils. Am. Soc. Microbiol. 2002, 68, 1893–1900. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.F.; Sobey, M.N.; Smith, C.J.; Rusmana, I.; Phillips, W.; Stott, A.; Osborn, A.M.; Nedwella, D.B. Dissimilatory reduction of nitrate to ammonium, not denitrification or anammox, dominates benthic nitrate reduction in tropical estuaries. Am. Soc. Limnol. Oceanogr. 2011, 56, 279–291. [Google Scholar] [CrossRef]
- Skalska, K.; Miller, J.S.; Ledakowicz, S. Kinetics of nitric oxide oxidation. Chem. Pap. 2010, 64, 269–272. [Google Scholar] [CrossRef]
- Griendling, K.K.; Touyz, R.M.; Zweier, J.L.; Chilian, W.; Chen, Y.-R.; Harrison, D.G.; Bhatnagar, A. Measurement of Reactive Oxygen Species, Reactive Nitrogen Species, and Redox-Dependent Signaling in the Cardiovascular System. Circ. Res. 2016, 119, e39–e75. [Google Scholar] [CrossRef]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS Sources in Physiological and Pathological Conditions. Oxid. Med. Cell. Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, C.M.; Machado, A.; Matos, P.; Bordalo, A.A. Impact of copper on the diversity, abundance and transcription of nitrite and nitrous oxide reductase genes in an urban European estuary. FEMS Microbiol. Ecol. 2011, 77, 274–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granger, J.; Ward, B.B. Accumulation of nitrogen oxides in copper-limited cultures of denitrifying bacteria. Limnol. Oceanogr. 2003, 48, 313–318. [Google Scholar] [CrossRef]
- Schmidt, I.; van Spanning, R.J.M.; Jetten, M.S.M. Denitrification and ammonia oxidation by Nitrosomonas europaea wild-type, and NirK- and NorB-deficient mutants. Microbiology 2004, 150, 4107–4114. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Burger, M.; Doane, T.A.; Horwath, W.R. Ammonia oxidation pathways and nitrifier denitrification are significant sources of N2O and NO under low oxygen availability. Proc. Natl. Acad. Sci. USA 2013, 110, 6328–6333. [Google Scholar] [CrossRef] [Green Version]
- Nelson, M.B.; Berlemont, R.; Martiny, A.C.; Martiny, J.B.H. Nitrogen Cycling Potential of a Grassland Litter Microbial Community. Appl. Environ. Microbiol. 2015, 81, 7012–7022. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, B.; Le Scornet, A.; Sauviac, L.; Remy, A.; Bruand, C.; Meilhoc, E. The Nitrate Assimilatory Pathway in Sinorhizobiummeliloti: Contribution to NO Production. Front. Microbiol. 2019, 10, 1526. [Google Scholar] [CrossRef]
- Maintinguer, S.I.; Sakamoto, I.K.; Adorno, M.A.T.; Varesche, M.B.A. Evaluation of the microbial diversity of denitrifying bacteria in batch reactor. Braz. J. Chem. Eng. 2013, 30, 457–465. [Google Scholar] [CrossRef]
- Kong, L.; Jing, H.; Kataoka, T.; Buchwald, C.; Liu, H. Diversity and Spatial Distribution of Hydrazine Oxidoreductase (hzo) Gene in the Oxygen Minimum Zone Off Costa Rica. PLoS ONE 2013, 8, e78275. [Google Scholar] [CrossRef]
- Wilson, R.W.; Murphy, K.L.; Sutton, P.M.; Lackey, S.L. Design and cost comparison of biological nitrogen removal processes. J. Water Pollut. Control. Fed. 1981, 53, 1294–1302. [Google Scholar]
- Tekere, M. Microbial Bioremediation and Different Bioreactors Designs Applied. In Biotechnology and Bioengineering; IntechOpen: London, UK, 2019. [Google Scholar]
- Langwaldt, J.H.; Puhakka, J.A. On-site biological remediation of contaminated groundwater: A review. Environ. Pollut. 2000, 107, 187–197. [Google Scholar] [CrossRef]
- Klein, R.; Tischler, J.S.; Műhling, M.; Schlömann, M. Bioremediation of mine water. Adv. Biochem. Eng./Biotechnol. 2013, 141, 109–172. [Google Scholar]
- Williams, P.; Botes, E.; Maleke, M.; Ojo, A.; DeFlaun, M.; Howell, J.; Borch, R.; Jordan, R.; van Heerden, E. Effective bioreduction of hexavalent chromium–contaminated water in fixed-film bioreactors. Water SA 2014, 40, 549. [Google Scholar] [CrossRef] [Green Version]
- Sun, P.; Zhuge, Y.; Zhang, J.; Cai, Z. Soil pH was the main controlling factor of the denitrification rates and N2/N2O emission ratios in forest and grassland soils along the Northeast China Transect (NECT). Soil Sci. Plant Nutr. 2012, 58, 517–525. [Google Scholar] [CrossRef]
- Sharma, S.; Bhattacharya, A. Drinking water contamination and treatment techniques. Appl. Water Sci. 2017, 7, 1043–1067. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.-Y.; Yang, P.; Liu, L.-S.; Liu, Z.-J. Formulation and Characterization of a Heterotrophic Nitrification-Aerobic Denitrification Synthetic Microbial Community and its Application to Livestock Wastewater Treatment. Water 2020, 12, 218. [Google Scholar] [CrossRef] [Green Version]
- Khanitchaidecha, W.; Sumino, T.; Kazama, F. Influence of carbon source on biological nitrogen removal by immobilised bacteria. J. Water Resour. Prot. 2010, 2, 527–531. [Google Scholar] [CrossRef] [Green Version]
- van Niel, E.W.J.; Arts, P.A.M.; Wesselink, B.J.; Robertson, L.A.; Kuenen, J.G. Competition between heterotrophic and autotrophic nitrifiers for ammonia in chemostat cultures. FEMS Microbil. Ecol. 1993, 102, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Robertson, L.A.; Kuenen, J.G. Combined heterotrophic nitrification and aerobic denitrification in Thiosphaera pantotropha and other bacteria. Antonie van Leeuwenhoek 1990, 57, 139–152. [Google Scholar] [CrossRef]
- Lee, N.M.; Welander, T. The effect of different carbon sources on respiratory denitrification in biological wastewater treatment. J. Ferment. Bioeng. 1996, 82, 277–285. [Google Scholar] [CrossRef]
- Xu, D.; Xiao, E.; Xu, P.; Lin, L.; Zhou, Q.; Xu, D.; Wu, Z. Bacterial community and nitrate removal by simultaneous heterotrophic and autotrophic denitrification in a bioelectrochemically-assisted constructed wetland. Bioresour. Technol. 2017, 245, 993–999. [Google Scholar] [CrossRef]
- Gou, J.; Hong, C.U.; Deng, M.; Chen, J.; Hou, J.; Li, D.; He, X. Effect of Carbon to Nitrogen Ratio on Water Quality and Community Structure Evolution in Suspended Growth Bioreactors through Biofloc Technology. Water 2019, 11, 1640. [Google Scholar] [CrossRef] [Green Version]
- Bethke, C.M.; Sanford, R.A.; Kirk, M.; Jin, Q.; Flynn, T. The thermodynamic ladder in geomicrobiology. Am. J. Sci. 2011, 311, 183–210. [Google Scholar] [CrossRef]
- Huang, S.; Zheng, Z.; Wei, Q.; Han, I.; Jaffé, P.R. Performance of sulfur-based autotrophic denitrification and denitrifiers for wastewater treatment under acidic conditions. Bioresour. Technol. 2019, 294, 122176. [Google Scholar] [CrossRef]
- Wlodarczyk, T.; Stepniewaska, Z.; Brzinska, M. Denitrification, organic matter and redox potential transformations in Cambisols. Int. Agrophysics 2003, 17, 219–227. [Google Scholar]
- Zou, G.; Ylinen, A.; Di Capua, F.; Papirio, S.; Lakaniemi, A.M.; Puhakka, J. Impact of heavy metals on denitrification of simulated mining wastewaters. Adv. Mater. Res. 2013, 825, 500–503. [Google Scholar] [CrossRef]
- Hollocher, T.C.; Kristjánsson, J.K. Thermophilic denitrifying bacteria: A survey of hot springs in Southwestern Iceland. FEMS Microbiol. Ecol. 1992, 101, 113–119. [Google Scholar] [CrossRef]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Jan, R.; Kang, S.; Kim, K.; Lee, I. Thermotolerance effect of plant growth-promoting Bacillus cereus SA1 on soybean during heat stress. BMC Microbiol. 2020, 20, 175. [Google Scholar] [CrossRef]
- Canion, A.; Kostka, J.E.; Gihring, T.M.; Huettel, M.; van Beusekom, J.E.E.; Gao, H.; Lavik, G.; Kuypers, M.M.M. Temperature response of denitrification and anammox reveals the adaptation of microbial communities to in situ temperatures in permeable marine sediments that span 50° in latitude. Biogeosciences 2014, 11, 309–320. [Google Scholar] [CrossRef] [Green Version]
- Carrera, J.; Vicent, T.; Lafuente, F.J. Influence of temperature on denitrification of an industrial high-strength nitrogen wastewater in a two-sludge system. Water SA 2003, 29, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Saleh-Lakha, S.; Shannon, K.E.; Henderson, S.L.; Goyer, C.; Trevors, J.T.; Zebarth, B.J.; Burton, D.L. Effect of pH and Temperature on Denitrification Gene Expression and Activity in Pseudomonas mandelii. Appl. Environ. Microbiol. 2009, 75, 3903–3911. [Google Scholar] [CrossRef] [Green Version]
- Prinčič, A.; Mahne, I.; Megušar, F.; Paul, E.A.; Tiedje, J.M. Effects of pH and oxygen and ammonium concentrations on the community structure of nitrifying bacteria from wastewater. Appl. Environ. Microbiol. 1998, 64, 3584–3590. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, H.; da Costa, T.B.; Wolff, D.B.; Platzer, C.; da Costa, R.H.R. The potential of denitrification for the stabilization of activated sludge processes affected by low alkalinity problems. Braz. Arch. Biol. Biotechnol. 2007, 50, 329–337. [Google Scholar] [CrossRef] [Green Version]
- He, T.; Xie, D.; Ni, J.; Li, Z.; Li, Z. Effect of Cobalt, Cadmium and Manganese on Nitrogen Removal Capacity of Arthrobacter arilaitensis Y-10. Water 2020, 12, 1701. [Google Scholar] [CrossRef]
- Magalhães, C.; Costa, J.; Teixeira, C.; Boralo, A. Impact of trace metals on denitrification in estuarine sediments of the Douro River estuary, Portugal. Mar. Chem. 2007, 107, 332–341. [Google Scholar] [CrossRef]
- Zhu-Barker, X.; Steenwerth, K.L. Chapter Six—Nitrous Oxide Production from Soils in the Future: Processes, Controls, and Responses to Climate Change. Dev. Soil Sci. 2018, 35, 131–183. [Google Scholar] [CrossRef]
- Rivett, M.; Buss, S.R.; Morgan, P.; Smith, J.W.N.; Bemment, C.D. Nitrate attenuation in groundwater: A review of biogeochemical controlling processes. Water Res. 2008, 42, 4215–4232. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Location | Contaminated Water Body | NO3−Concentration mg/L | Reference |
|---|---|---|---|---|
| Explosives factory | China | Wastewater catchment | 3600 | [43] |
| Explosives factory | Poland | Wastewater catchment | 3000 | [42] |
| Farm | Pakistan | Groundwater | 1610 | [44] |
| Farm | Namibia | Groundwater | 1000 | [13] |
| Sewage | Tanzania | Groundwater | 929 | [45] |
| Farm | South Africa | Surface water | 193 | [46] |
| Sewage | Iran | Aquifer | 166 | [47] |
| Mine | China | Groundwater | 109 | [22] |
| Mine | South Africa | River | 50 | [41] |
| Metabolism | Electron Donating/Accepting Half-Reaction | e− don/acc | ∆G (Kj mol−1) |
|---|---|---|---|
| Autotrophy | CH4COO− + 4H2O → 2HCO3− + 9H+ + 8e− | e− donating | −216 |
| Hydrogenotrophy | 4H2(aq) → 8H+ + 8e− | e− donating | −185 |
| Denitrification | 8e− + 8/5 NO3− + 48/5H+→4/5N2(aq) + 24/5H2O | e− accepting | −550 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moloantoa, K.M.; Khetsha, Z.P.; van Heerden, E.; Castillo, J.C.; Cason, E.D. Nitrate Water Contamination from Industrial Activities and Complete Denitrification as a Remediation Option. Water 2022, 14, 799. https://doi.org/10.3390/w14050799
Moloantoa KM, Khetsha ZP, van Heerden E, Castillo JC, Cason ED. Nitrate Water Contamination from Industrial Activities and Complete Denitrification as a Remediation Option. Water. 2022; 14(5):799. https://doi.org/10.3390/w14050799
Chicago/Turabian StyleMoloantoa, Karabelo M., Zenzile P. Khetsha, Esta van Heerden, Julio C. Castillo, and Errol D. Cason. 2022. "Nitrate Water Contamination from Industrial Activities and Complete Denitrification as a Remediation Option" Water 14, no. 5: 799. https://doi.org/10.3390/w14050799
APA StyleMoloantoa, K. M., Khetsha, Z. P., van Heerden, E., Castillo, J. C., & Cason, E. D. (2022). Nitrate Water Contamination from Industrial Activities and Complete Denitrification as a Remediation Option. Water, 14(5), 799. https://doi.org/10.3390/w14050799