
Effects of the Invasive Fish Species Ameiurus nebulosus on Microbial Communities in Peat Pools

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
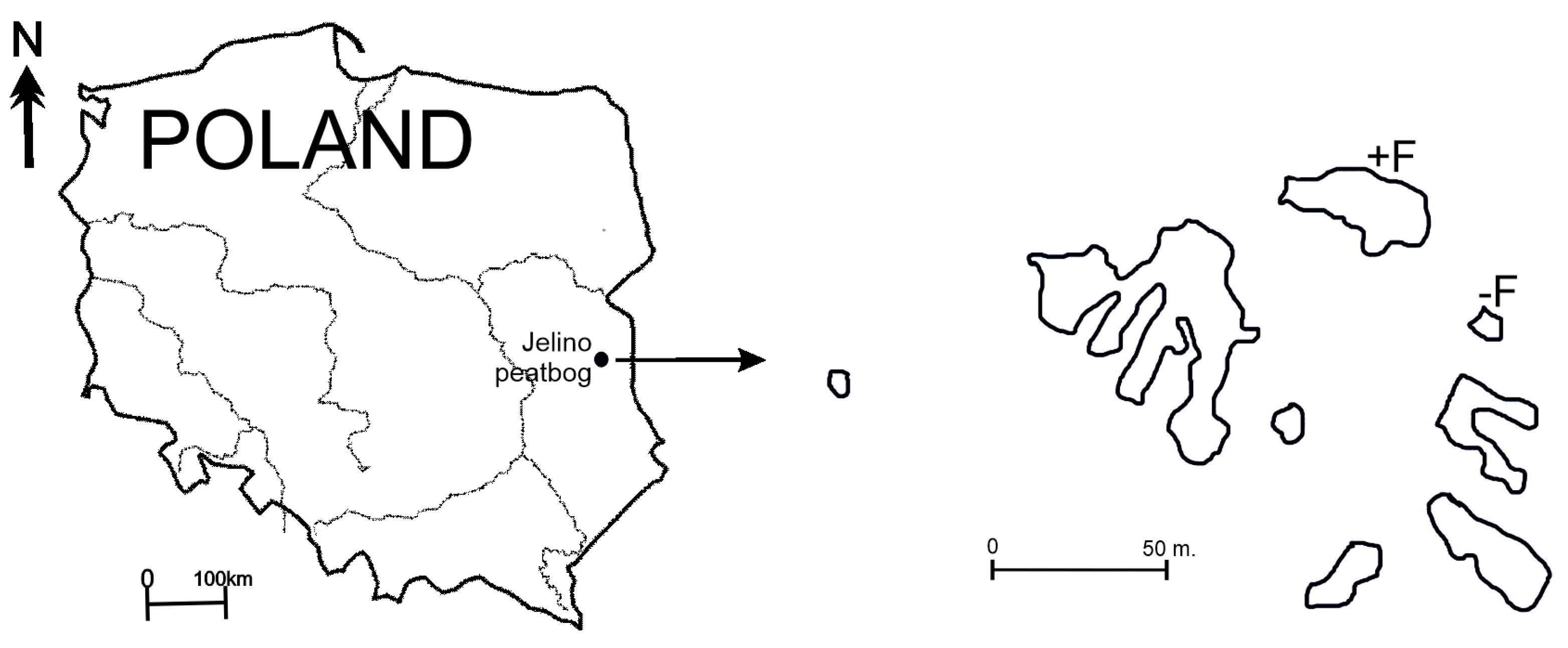
2.1. Study Area
2.2. Microcosm Experiments
2.3. Laboratory Analyses
2.4. Physical and Chemical Variables
2.5. Numerical Analyses
3. Results
3.1. Environmental Variables
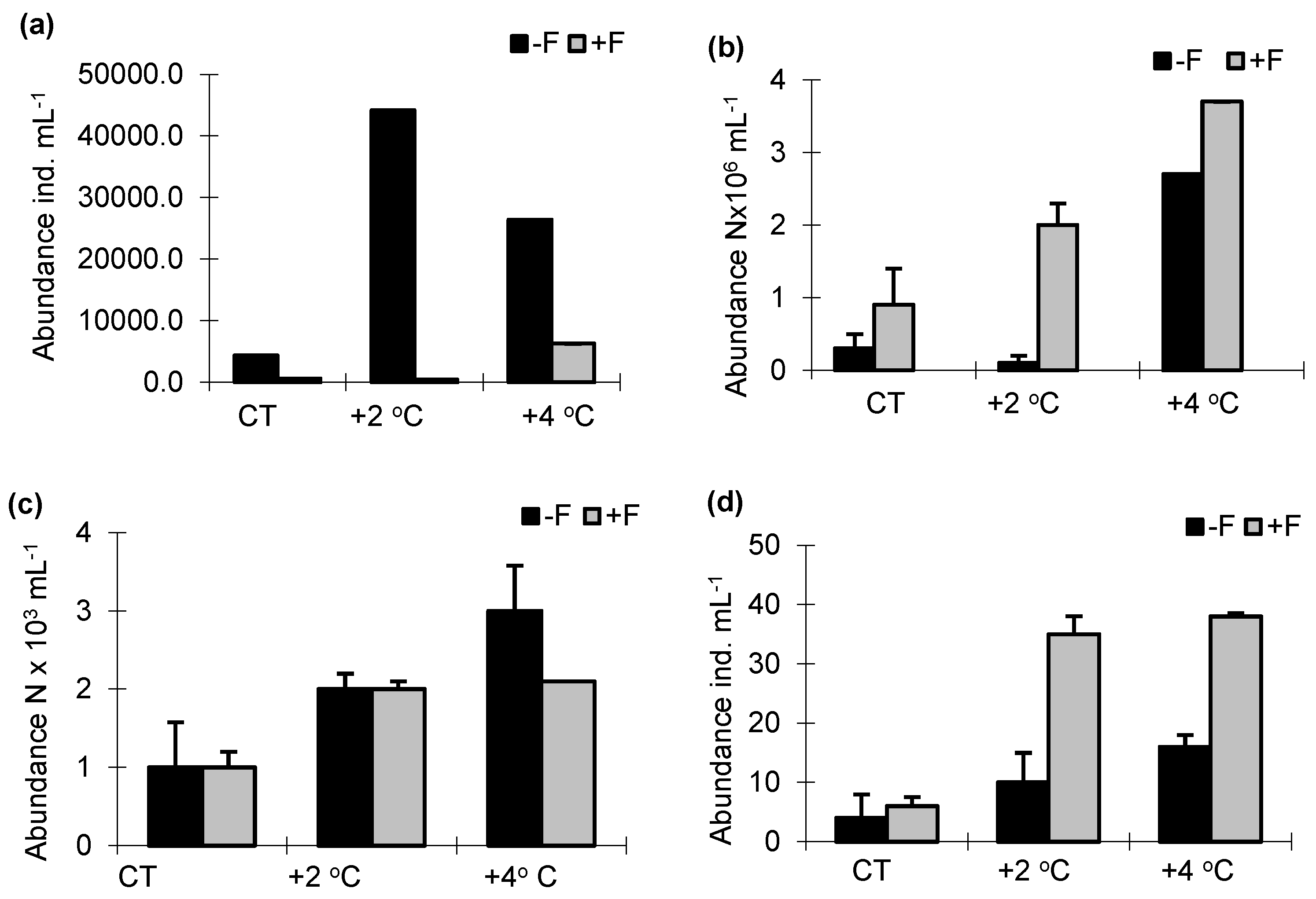
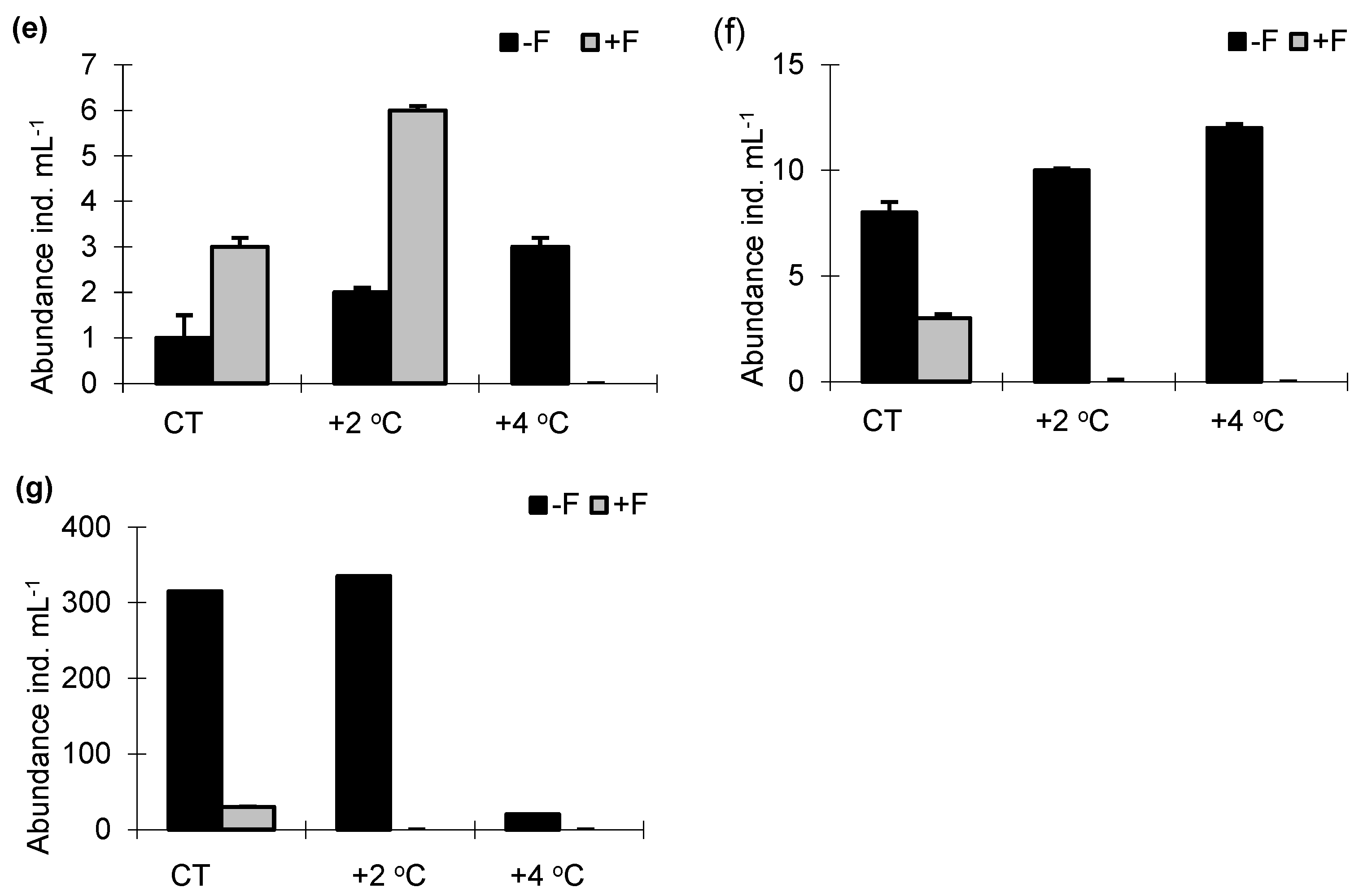
3.2. Abundance and Size Structure
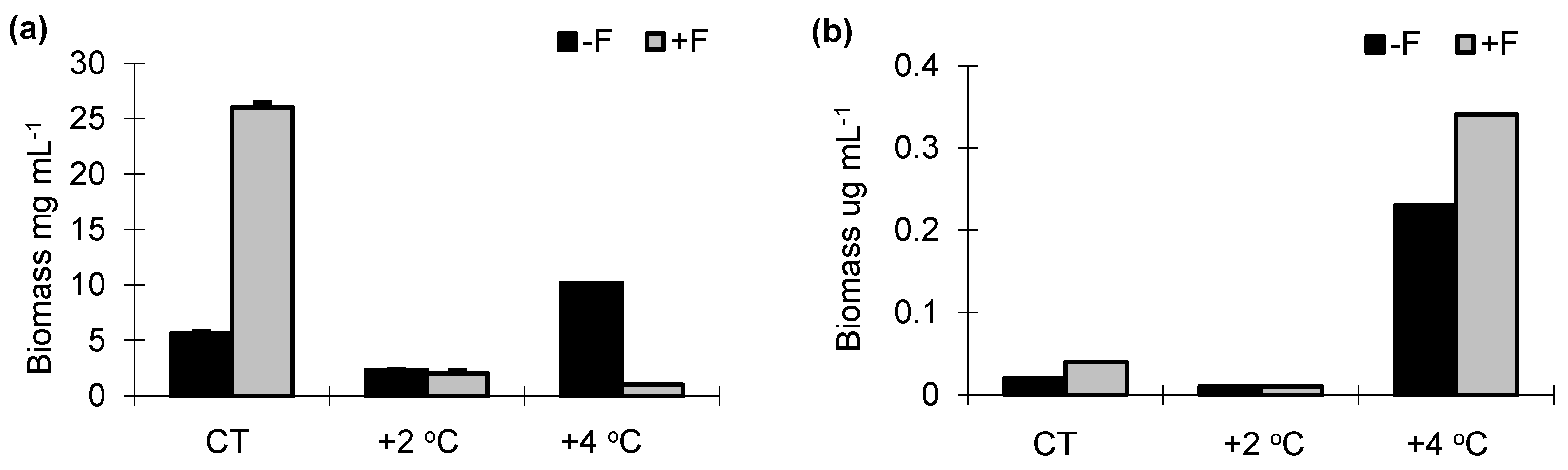
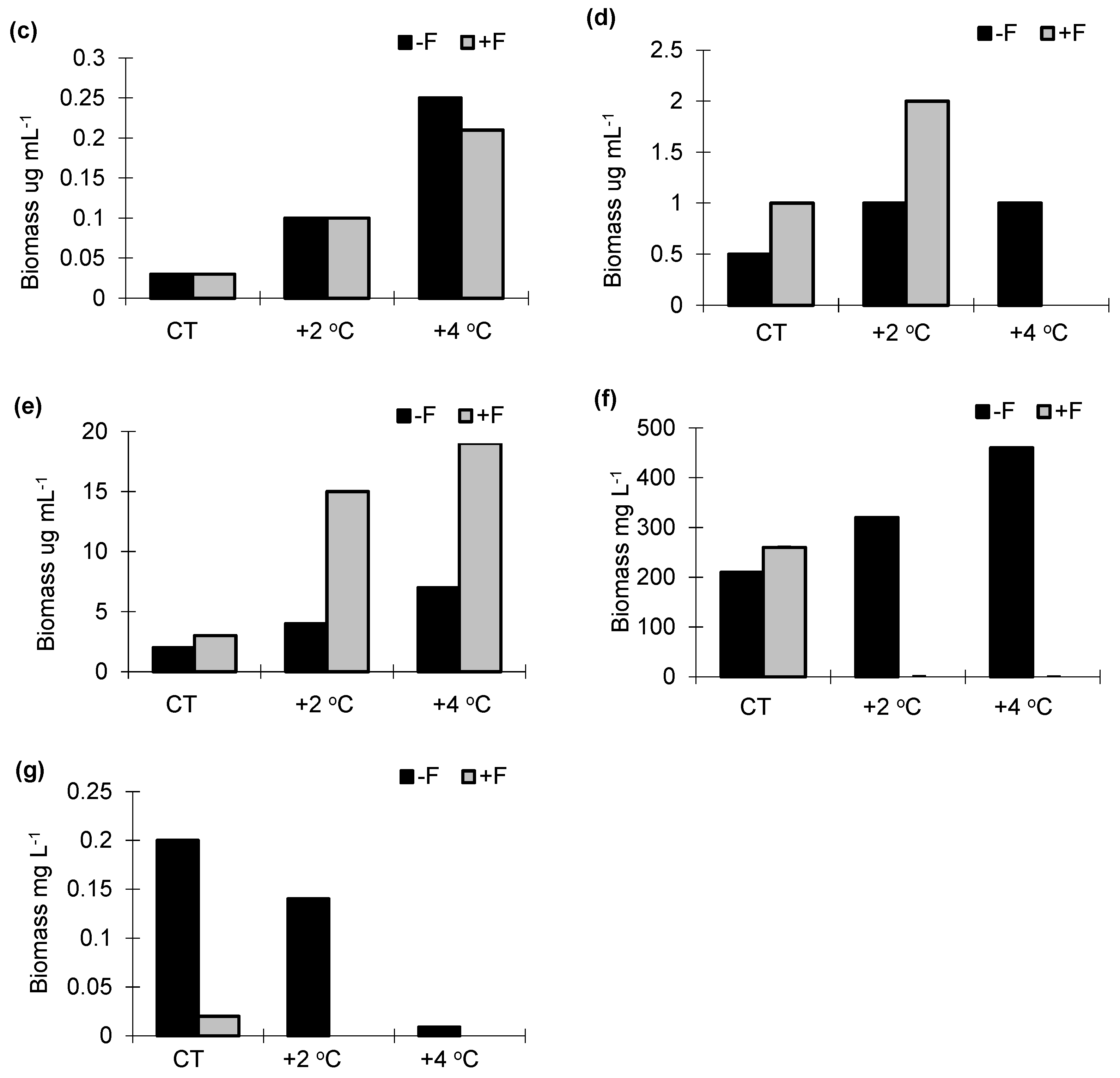
3.3. Biomass
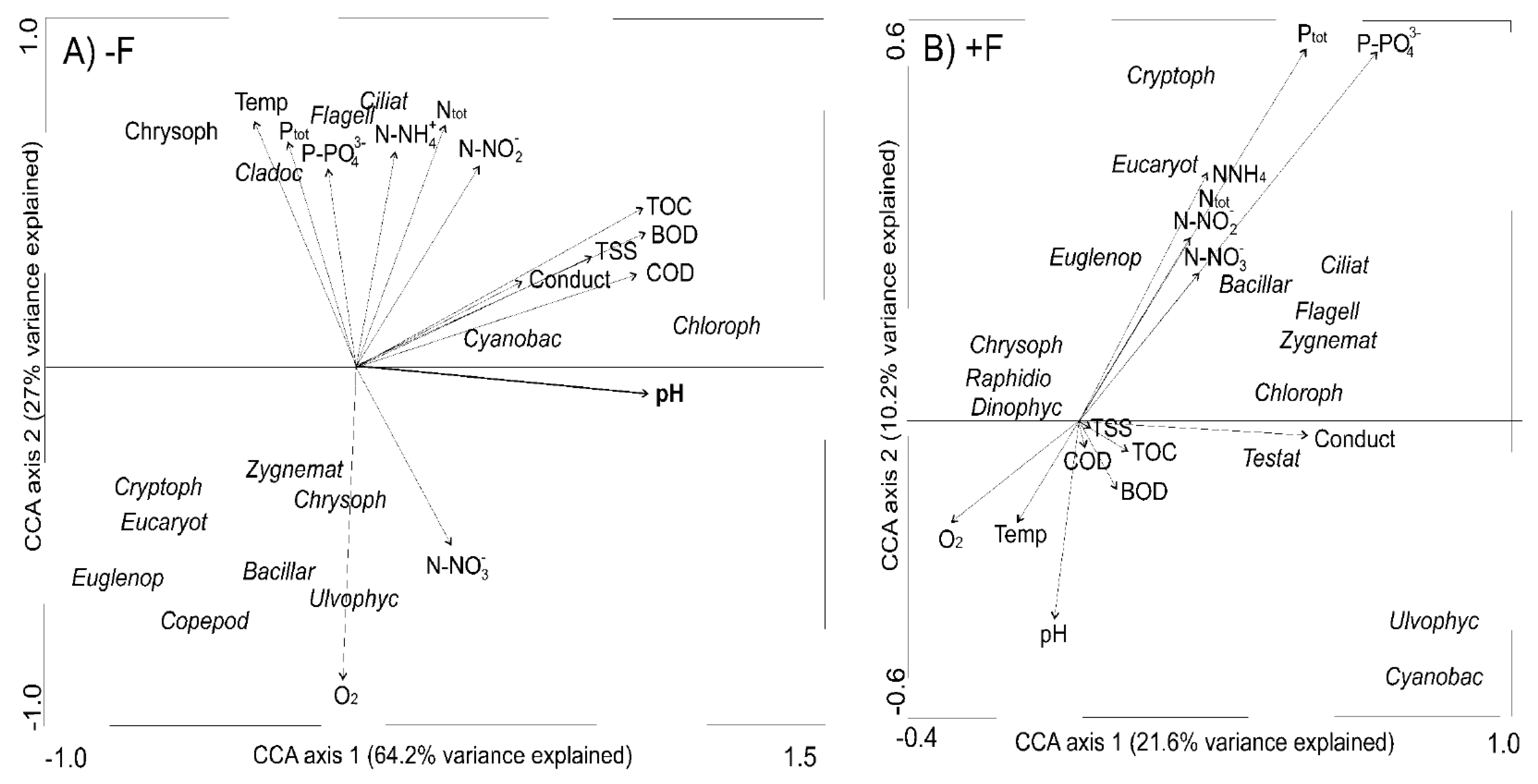
3.4. Microbial Communities vs. Environmental Variables
3.5. Relationships between Food Web Components
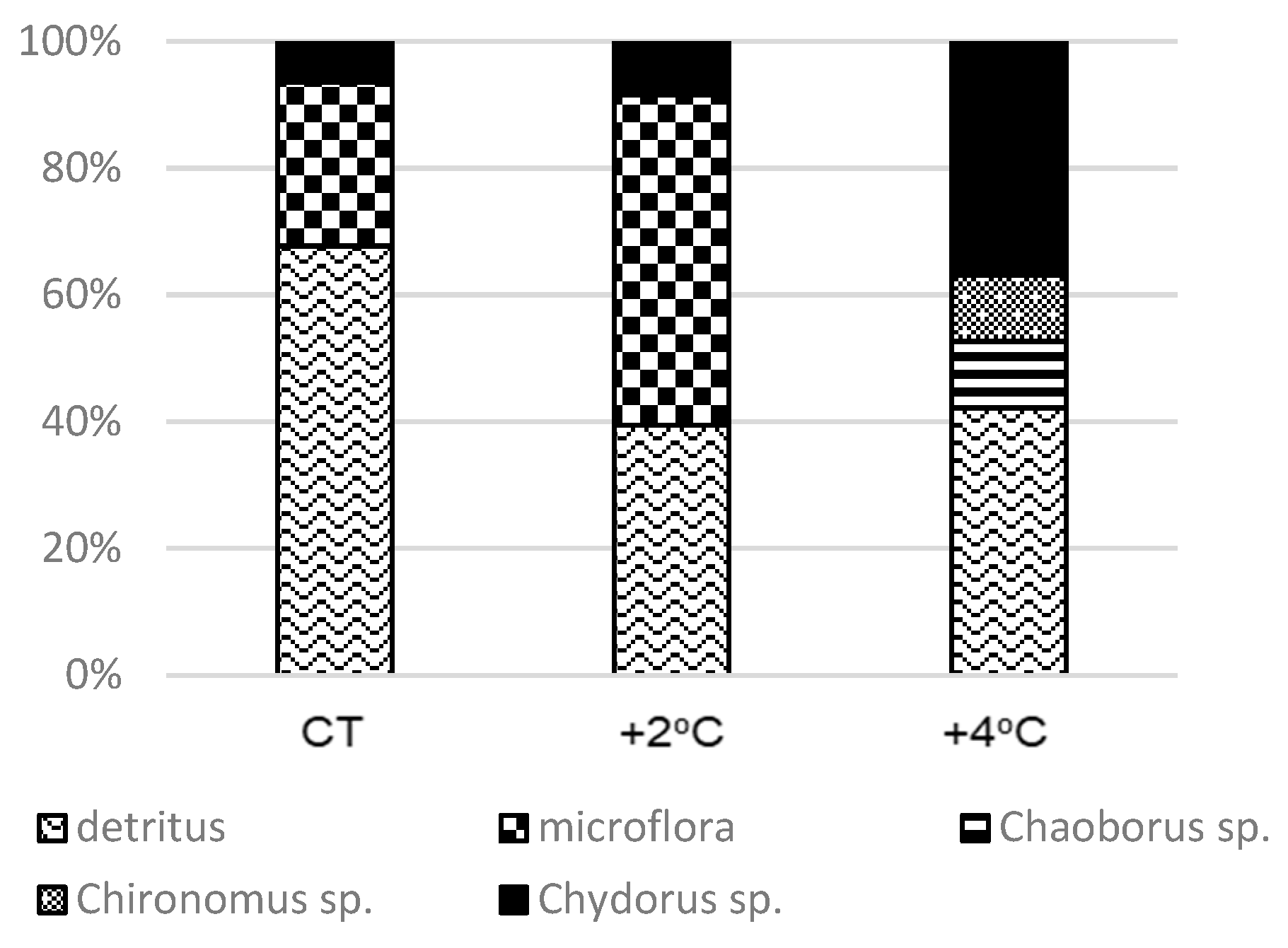
3.6. Stomach Contents of Ameiurus nebulosus
4. Discussion
4.1. Diet of Ameiurus nebulosus
4.2. Functioning of the Food Webs in Treatments with Ameiurus nebulosus
4.3. Functioning of the Food Webs in Fishless Treatments
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bragazza, L.; Freeman, C.; Jones, T.; Rydin, H.; Limpens, J.; Fenner, N.; Ellis, T.; Gerdol, R.; Hàjek, M.; Iacumin, P.; et al. Atmospheric nitrogen deposition promotes carbon loss from peat bogs. Proc. Natl. Acad. Sci. USA 2006, 103, 19386–19389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robson, T.M.; Pancotto, V.A.; Scopel, A.L.; Flint, S.D.; Caldwell, M. Solar UV-B influences microfaunal community composition in a Tierra del Fuego peatland. Soil Biol. Biochem. 2005, 37, 2205–2215. [Google Scholar] [CrossRef]
- Burford, M.A.; Thompson, P.J.; McIntosh, R.P.; Bauman, R.H.; Pearson, D.C. Nutrient and microbial dynamics in high-intensity, zero-exchange shrimp ponds in Belize. Aquaculture 2003, 219, 393–411. [Google Scholar] [CrossRef] [Green Version]
- Tranvik, L.J.; Downing, J.A.; James, B.; Cotner, J.B.; Steven, A.; Loiselle, S.A.; Striegl, R.G.; Ballatore, T.J.; Dillon, P.; Finlay, K.; et al. Lakes and reservoirs as regulators of carbon cycling and climate. Limnol. Oceanogr. 2009, 54, 2298–2314. [Google Scholar] [CrossRef] [Green Version]
- Copp, G.H.; Bianco, P.G.; Bogutskaya, N.G.; Eros, T.; Falka, I.; Ferreira, M.T.; Fox, M.G.; Freyhof, J.; Gozlan, R.E.; Grabowska, J.; et al. To be, or not to be, a non-native freshwater fish? J. Appl. Ichthyol. 2005, 21, 242–262. [Google Scholar] [CrossRef]
- Rabitsch, W.; Milasowszky, N.; Nehring, S.; Wiesner, C.; Wolter, C.; Essl, F. The times are changing temporal shifts in patterns of fish invasions in central European fresh waters. J. Fish Biol. 2013, 82, 17–33. [Google Scholar] [CrossRef]
- Rechulicz, J.; Płaska, W. The diet of non-indigenous Ameiurus nebulosus of varying size and its potential impact on native fish in shallow lakes. Glob. Ecol. Conserv. 2021, 1, e01881. [Google Scholar] [CrossRef]
- Leunda, P.M.; Oscoz, J.; Elvira, B.; Agorreta, A.; Perea, S.; Miranda, R. Feeding habits of the exotic black bullhead Ameiurus melas (Rafinesque) in the Iberian Peninsula: First evidence of direct predation on native fish species. J. Fish Biol. 2008, 73, 96–114. [Google Scholar] [CrossRef]
- Gunn, J.M.; Qadri, S.U.; Mortimer, D.C. Filamentous algae as a food source for the brown bullhead (Ictalurus nebulosus). J. Fish. Res. Board. Can. 1977, 34, 396–401. [Google Scholar] [CrossRef]
- Hill, T.; Duffy, W.G.; Thomson, M.R. Food habits channel catfish in Lake Oahe, South Dakota. J. Freshwat. Ecol. 1995, 10, 319–323. [Google Scholar] [CrossRef]
- Declerck, S.; Louette, G.; De Bie, T.; De Meester, L. Patterns of diet overlap between populations of non-indigenous and native fishes in shallow ponds. J. Fish Biol. 2002, 61, 1182–1197. [Google Scholar] [CrossRef]
- Jeppesen, E.; Christoffersen, K.; Landkilehus, F.; Lauridsen, T.; Amsinck, S.L.; Riget, F.; Söndergaard, M. Fish and crustaceans in northeast Greenland lakes with special emphasis on interactions between Arctic charr (Salvelinus alpinus), Lepidurus arcticus and benthic chydorids. Hydrobiologia 2001, 442, 329–337. [Google Scholar] [CrossRef]
- Riemann, B. Potential importance of fish pre-dation and zooplankton grazing on natural populations of freshwater bacteria. Appl. Environ. Microbiol. 1985, 50, 187–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Foster, J.; Hirst, A.G.; Esteban, G.F. Achieving temperature-size changes in unicellular organism. ISME 2013, 7, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Zingel, P.; Cremona, F.; Noges, T.; Cao, Y.; Neif, E.; Coppens, J.; Iskin, U.; Lauridsen, L.; Davidson, T.A.; Sondergaard, M.; et al. Effect of warming and nutrients on the microbial food web in shallow lake mesocosms. Eur. J. Protistol. 2018, 64, 1–12. [Google Scholar] [CrossRef]
- Jassey, V.E.J.; Chiapusio, G.; Binet, P.; Buttler, A.; Laggoun-Defarge, F.; Delarue, F.; Gilbert, D. Above- and belowground linkages in Sphagnum peatland: Climate warming affects plant-microbial interactions. Glob. Chang. Biol. 2013, 19, 811–823. [Google Scholar] [CrossRef] [Green Version]
- Mieczan, T.; Tarkowska-Kukuryk, M. Diurnal dynamics of the microbial loop in peatlands: Structure, function, and relationship to environmental parameters. Hydrobiologia 2013, 717, 189–201. [Google Scholar] [CrossRef] [Green Version]
- Hillebrand, H.; Dürselen, C.D.; Kirschtel, D.; Pollingher, U.; Zohary, T. Biovolume calculation for pelagic and benthic microalgae. J. Phycol. 1999, 35, 403–424. [Google Scholar] [CrossRef]
- Komárek, J.; Fott, B. Chlorophyceae (Grünalgen) Ordnung: Chlorococcales; Schweizerbart: Stuttgart, Germany, 1983; pp. 1–1001. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota. Chroococcales Teil 1; Spektrum Akademischer Verlag: Berlin, Germany, 1999; pp. 1–548. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Chroococcales Teil 1; Spektrum akademischer Verlag: Berlin, Germany, 2000; pp. 1–548. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Oscillatoriales. Teil 2; Spektrum Akademischer Verlag: München, Germany, 2005; pp. 1–759. [Google Scholar]
- Komárek, J. Cyanoprokaryota Teil 3: Heterocytous Genera.; Springer: Berlin, Germany, 2013; pp. 1–1131. [Google Scholar]
- Starmach, K. Süßwasserflora von Mitteleuropa. Chrysophyceae und Haptophyceae Teil 1.; Spektrum Akademischer Verlag: Stuttgart, Germany, 1985; pp. 1–515. [Google Scholar]
- Cox, E.J. Identification of Freshwater Diatoms from Live Material; Chapman and Hall: London, UK, 1996; pp. 1–156. [Google Scholar]
- Van den Hoek, C.; Mann, D.G.; Jahns, H.M. Algae. An Introduction to Phycology; Cambridge University Press: Cambridge, NY, USA, 1995; pp. 1–623. [Google Scholar]
- Utermöhl, H. Zur vervollkommung der quantative phytoplankton methodic. Mitt. Internat. Verein. Limnol. 1958, 9, 1–38. [Google Scholar]
- Foissner, W.; Berger, H. A user-friendly guide to the ciliates (Protozoa, Ciliophora) commonly used by hydrobiologists as bioindicators in rivers, lakes, and waste waters, with notes on their ecology. Freshw. Biol. 1996, 35, 375–482. [Google Scholar] [CrossRef]
- Foissner, W.; Berger, H.; Schaumburg, J. Identification and Ecology of Limnetic Plankton Ciliates; Bayerisches Landesamt für Wasserwirtschaft: München, Germany, 1999; pp. 1–793. [Google Scholar]
- Charman, D.J.; Hendon, D.; Woodland, W. The identification of testate amoebae (Protozoa: Rhizopoda) in peats. Quat. Res. Tech. Guide 2000, 9, 1–147. [Google Scholar]
- Dumont, H.J.; Van de Velde, I.; Dumont, S. The dry weight estimate of biomass in selection of Cladocera, Copepoda and Rotifera from the plankton, periphiton and benthos of continental waters. Oecologia 1975, 19, 75–97. [Google Scholar] [CrossRef] [PubMed]
- Bottrell, H.H.; Duncan, A.; Gliwicz, Z.M.; Grygierek, E.; Herzig, A.; Hillbricht–Ilkowska, A.; Kurasawa, H.; Larsson, P.; Węgleńska, T. Review of some problems in zooplankton production studies. Norw. J. Zool. 1976, 24, 419–456. [Google Scholar]
- Culver, D.A.; Boucherle, M.M.; Bean, D.J.; Fletcher, W.J. Biomass of freshwater crustacean zooplankton from lenght–weight regression. Can. J. Fish. Aquat. Sci. 1985, 42, 1380–1390. [Google Scholar] [CrossRef]
- Hyslop, E.J. Stomach contents analysis—A review of methods and their application. J. Fish Biol. 1980, 17, 411–429. [Google Scholar] [CrossRef] [Green Version]
- Kołodziejczyk, A.; Koperski, P. Polish Invertebrates of Freshwaters. Manual to Determination and the Basics of the Biology and Ecology of Macrofauna; UW: Warsaw, Poland, 2000; pp. 1–250. [Google Scholar]
- Rybak, J.I. Freshwater Invertebrate Animals. Recognition Guide; PWN: Warsaw, Poland, 2000; pp. 1–87. [Google Scholar]
- Golterman, H.L. Methods for Chemical Analysis of Fresh Waters; Blackwell Scientific: Oxford, UK, 1969; pp. 1–166. [Google Scholar]
- Ter Braak, C.J.F. CANOCO–FORTRAN Program for Canonical Community Ordination; Microcomputer Power: Ithaca, NY, USA, 1988; pp. 1–96. [Google Scholar]
- Gilbert, D.; Amblard, C.; Bourdier, G.; Francez, A.J. The microbial loop at the surface of a peatland: Structure, functioning and impact of nutrients inputs. Microb. Ecol. 1998, 35, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.R.; Krogman, R.M.; Quist, M.C. Influences of native and non-native benthivorous fishes on aquatic ecosystem degradation. Hydrobiologia 2013, 711, 187–199. [Google Scholar] [CrossRef]
- Buosi, P.R.B.; Pauleto, G.M.; Lansac-Toha, F.A.; Velho, L.F.M. Ciliate community associated with aquatic macrophyte roots: Effects of nutrient enrichment on the community composition and species richness. Eur. J. Protistol. 2011, 47, 86–102. [Google Scholar] [CrossRef]
- Zingel, P.; Huitu, E.; Mäkelä, S.; Arvola, L. The abundance and diversity of planktonic ciliates in 12 boreal lakes of varying trophic state. Arch Hydrobiol. 2002, 155, 315–332. [Google Scholar] [CrossRef]
- Andersen, R.; Wells, C.; Macrae, M.; Price, J. Nutrient mineralization and microbial functional diversity in a restored bog approach natural conditions 10 years post restoration. Soil Biol. Biochem. 2013, 64, 37–47. [Google Scholar] [CrossRef]
- Šimek, K.; Bobkova, J.; Macek, M.; Nedoma, J. Ciliate grazing on picoplankton in a eutrophic reservoir during the summer phytoplankton maximum: A study at the species and community level. Limnol. Oceanogr. 1995, 40, 1077–1090. [Google Scholar] [CrossRef]
- Kisand, V.; Zingel, P. Dominance of ciliate grazing on bacteria during spring in a shallow eutrophic lake. Aquat. Microb. Ecol. 2000, 22, 135–142. [Google Scholar] [CrossRef]
- Auer, B.; Arndt, H. Taxonomic composition, and biomass of heterotrophic flagellates in relation to lake trophy and season. Freshw. Biol. 2001, 46, 959–972. [Google Scholar] [CrossRef]
- Mieczan, T.; Bielańska-Grajner, I.; Tarkowska-Kukuryk, M. Hydrochemical and microbiological distinction and function of ombrotrophic peatland lagg as ecotone between Sphagnum peatland and forest catchment (Poleski National Park, eastern Poland). Ann. Limnol. Internat. J. Limnol. 2012, 48, 323–336. [Google Scholar] [CrossRef] [Green Version]
- Marcisz, K.; Lamentowicz, Ł.; Słowińska, S.; Słowiński, M.; Muszak, W.; Lamentowicz, M. Seasonal changes in Sphagnum peatland testate amoebae communities along hydrological gradient. Europ. J. Protistol. 2014, 50, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Laybourn, G.; Whymant, L. The effect of diet and temperature on reproductive rate in Arcella vulgaris Ehrenberg (Sarcodina: Testacida). Oecologia 1980, 45, 282–284. [Google Scholar] [CrossRef] [PubMed]
- Hahn, M.W.; Höfle, M.G. Grazing of protozoa and its effect on populations of aquatic bacteria. FEMS Microbiol. Eco. 2001, 35, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Heckmann, L.; Drossel, B.; Brose, U.; Guill, C. Interactive effects of body-size structure and adaptive foraging on food-web stability. Ecol. Lett. 2012, 15, 243–250. [Google Scholar] [CrossRef]
- Mieczan, T.; Adamczuk, M.; Pawlik-Skowrońska, B.; Toporowska, M. Eutrophication of peatbogs: Consequences of P and N enrichment for microbial and metazoan communities in mesocosm experiments. Aquat. Microb. Ecol. 2015, 74, 121–141. [Google Scholar] [CrossRef]
- Kazanjian, G.; Velthuis, M.; Aben, R.; Stephan, S.; Peeters, E.T.H.M.; Frenken, T.; Touwen, J.; Xue, F.; Kosten, S.; Van de Waal, D.B.; et al. Impacts 638 of warming on top-down and bottom-up controls of periphyton production. Sci. Rep. 2018, 8, 9901. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters/Treatment | −FCT | −FT + 2 °C | −FT + 4 °C | +FCT | +F + 2 °C | +F + 4 °C |
|---|---|---|---|---|---|---|
| Temperature °C | 22 | 24 | 26 | 22 | 24 | 26 |
| pH | 3.78 | 5.42 | 5.03 | 4.13 | 6.48 | 6.39 |
| Conductivity µS cm−1 | 31 | 57 | 76 | 35 | 94 | 65 |
| O2 mg L−1 | 4.03 | 5.16 | 5.32 | 6.89 | 4.88 | 9.98 |
| N-NH4+ mg L−1 | 0.025 | 3.797 | 5.678 | 0.036 | 8.384 | 10.855 |
| N-NO2− mg L−1 | 0.062 | 0.042 | 0.053 | 0.009 | 0.075 | 0.152 |
| N-NO3− mg L−1 | 0.007 | 0.311 | 0.036 | 0.017 | 0.048 | 0.089 |
| P-PO43− mg L−1 | 0.005 | 0.010 | 0.528 | 0.023 | 0.892 | 0.156 |
| Ptot mg L−1 | 0.060 | 0.084 | 0.637 | 0.046 | 1.116 | 0.250 |
| Ntot mg L−1 | 0.984 | 5.241 | 6.579 | 0.927 | 8.443 | 15.750 |
| TSS mg L−1 | 180.0 | 385.0 | 440.0 | 130.0 | 485.0 | 940.0 |
| TOC mg L−1 | 32.0 | 55.5 | 53.0 | 27.0 | 62.0 | 104.0 |
| COD mg L−1 | 72.0 | 126.0 | 126.0 | 59.0 | 147.0 | 236.0 |
| BOD mg L−1 | 43.0 | 73.5 | 70.5 | 36.0 | 83.0 | 136.0 |
| −FCT | −FT + 2 °C | −FT + 4 °C | ||||||||||||||||
| Phy | B | HNF | TA | C | Cr | Phy | B | HNF | TA | C | Cr | Phy | B | HNF | TA | C | Cr | |
| Phy | - | 0.71 | - | - | ||||||||||||||
| B | - | −0.82 | 0.79 | - | - | |||||||||||||
| HNF | −0.78 | - | - | - | - | |||||||||||||
| TA | −0.69 | - | −0.71 | - | −0.77 | - | ||||||||||||
| C | −0.57 | 0.35 | - | 0.51 | - | - | ||||||||||||
| Cr | 0.43 | - | - | 0.53 | - | |||||||||||||
| +FCT | +FT + 2 °C | +FT + 4 °C | ||||||||||||||||
| Phy | - | 0.74 | 0.65 | - | - | |||||||||||||
| B | - | 0.71 | −0.73 | - | 0.5 | −0.73 | - | −0.75 | ||||||||||
| HNF | - | −0.65 | - | 0.37 | - | |||||||||||||
| TA | - | - | - | |||||||||||||||
| C | - | - | - | |||||||||||||||
| Cr | 0.43 | - | 0.5 | - | 0.42 | - | ||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mieczan, T.; Płaska, W.; Adamczuk, M.; Toporowska, M.; Bartkowska, A. Effects of the Invasive Fish Species Ameiurus nebulosus on Microbial Communities in Peat Pools. Water 2022, 14, 815. https://doi.org/10.3390/w14050815
Mieczan T, Płaska W, Adamczuk M, Toporowska M, Bartkowska A. Effects of the Invasive Fish Species Ameiurus nebulosus on Microbial Communities in Peat Pools. Water. 2022; 14(5):815. https://doi.org/10.3390/w14050815
Chicago/Turabian StyleMieczan, Tomasz, Wojciech Płaska, Małgorzata Adamczuk, Magdalena Toporowska, and Aleksandra Bartkowska. 2022. "Effects of the Invasive Fish Species Ameiurus nebulosus on Microbial Communities in Peat Pools" Water 14, no. 5: 815. https://doi.org/10.3390/w14050815
APA StyleMieczan, T., Płaska, W., Adamczuk, M., Toporowska, M., & Bartkowska, A. (2022). Effects of the Invasive Fish Species Ameiurus nebulosus on Microbial Communities in Peat Pools. Water, 14(5), 815. https://doi.org/10.3390/w14050815