
Genotypic Diversity Improves Photosynthetic Traits of Hydrocotyle vulgaris and Alters Soil Organic Matter and N2O Emissions of Wetland Microecosystems

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
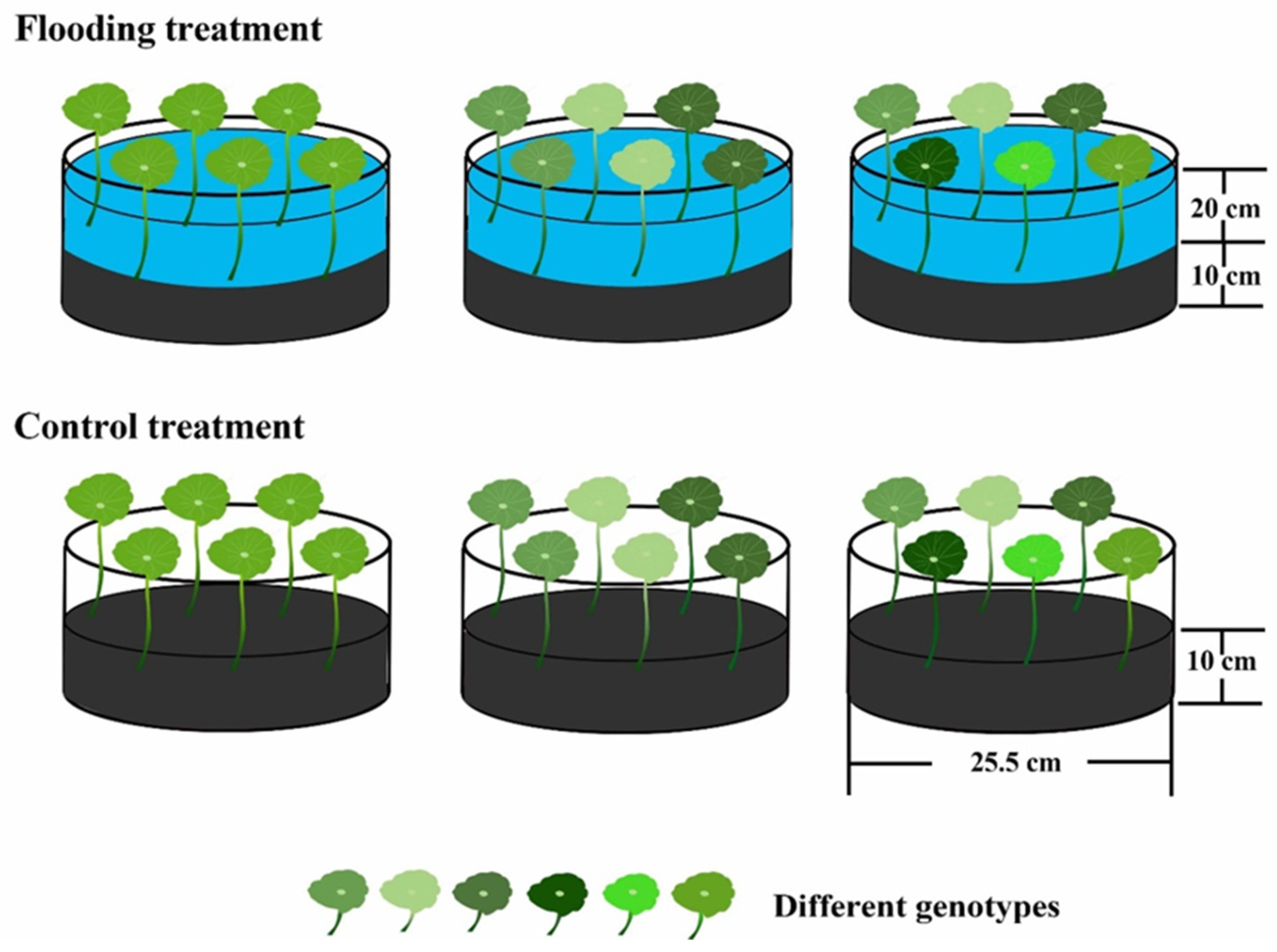
2.2. Experimental Design
2.3. Harvest and Measurements
2.4. Data Analysis
2.4.1. The Net Diversity, Complementarity, and Selection Effects
2.4.2. Greenhouse Gas Emissions
2.4.3. Statistical Analysis
3. Results
3.1. Effects of Genotypic Diversity and Flooding on the Invasiveness of H. vulgaris
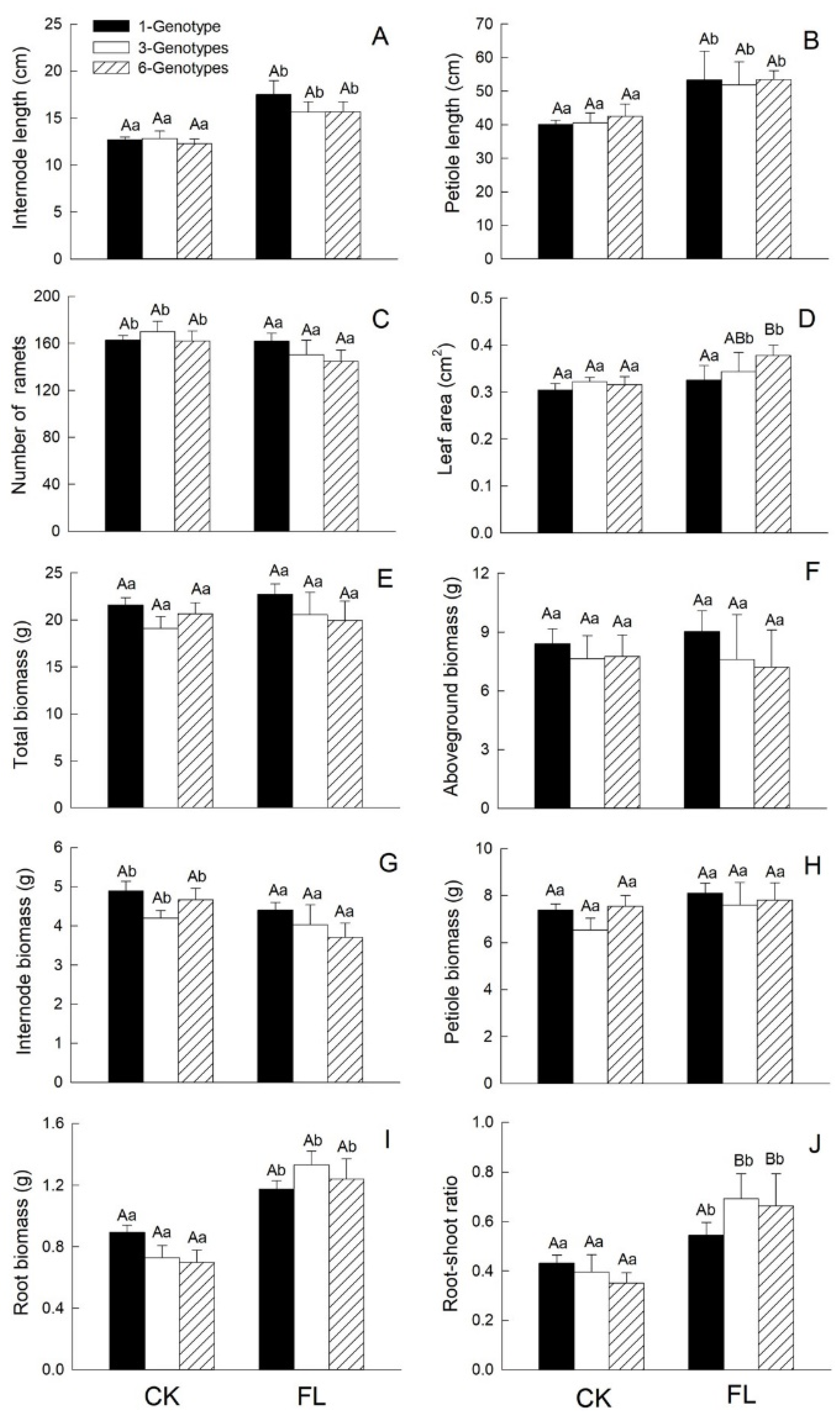
3.1.1. Growth Traits
3.1.2. Photosynthetic Physiological Traits
3.1.3. The Net Diversity, Complementarity, and Selection Effects
3.2. Effects of Genotypic Diversity and Flooding on the Wetland Microecosystem
3.2.1. Soil Properties
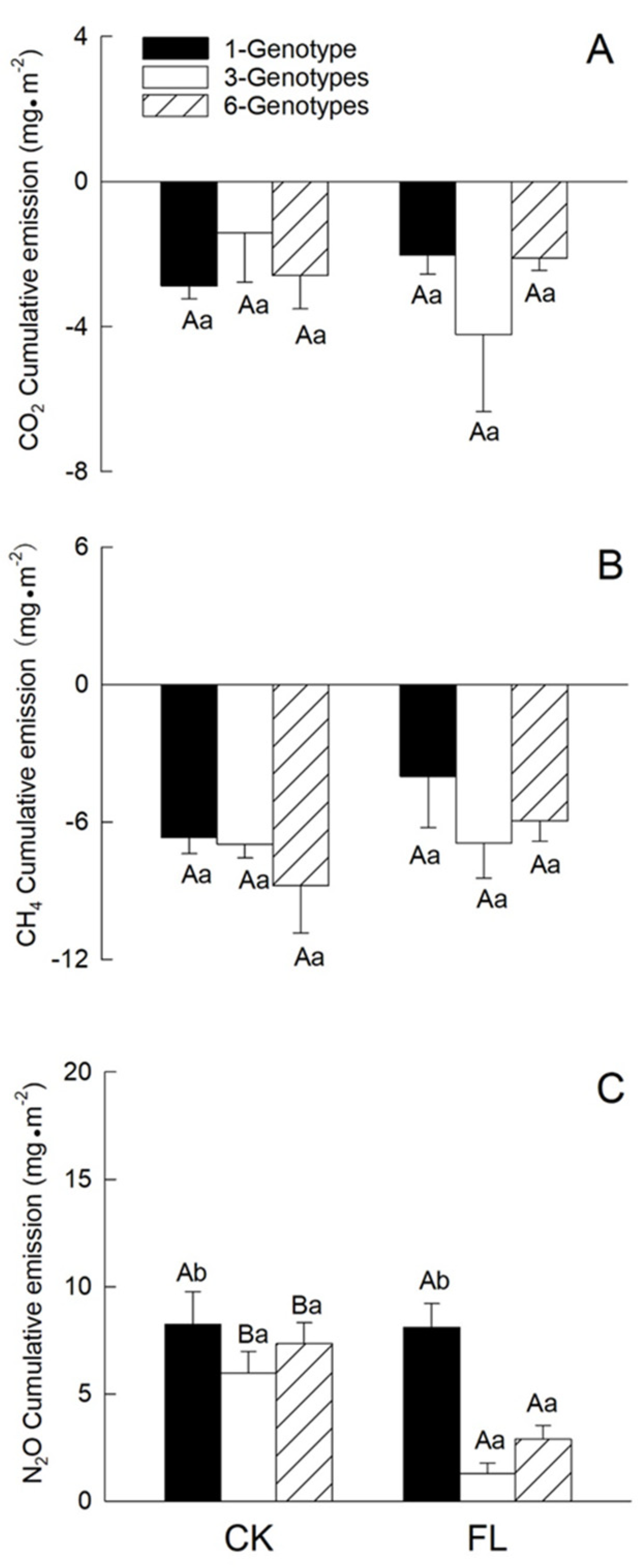
3.2.2. Greenhouse Gas Emissions
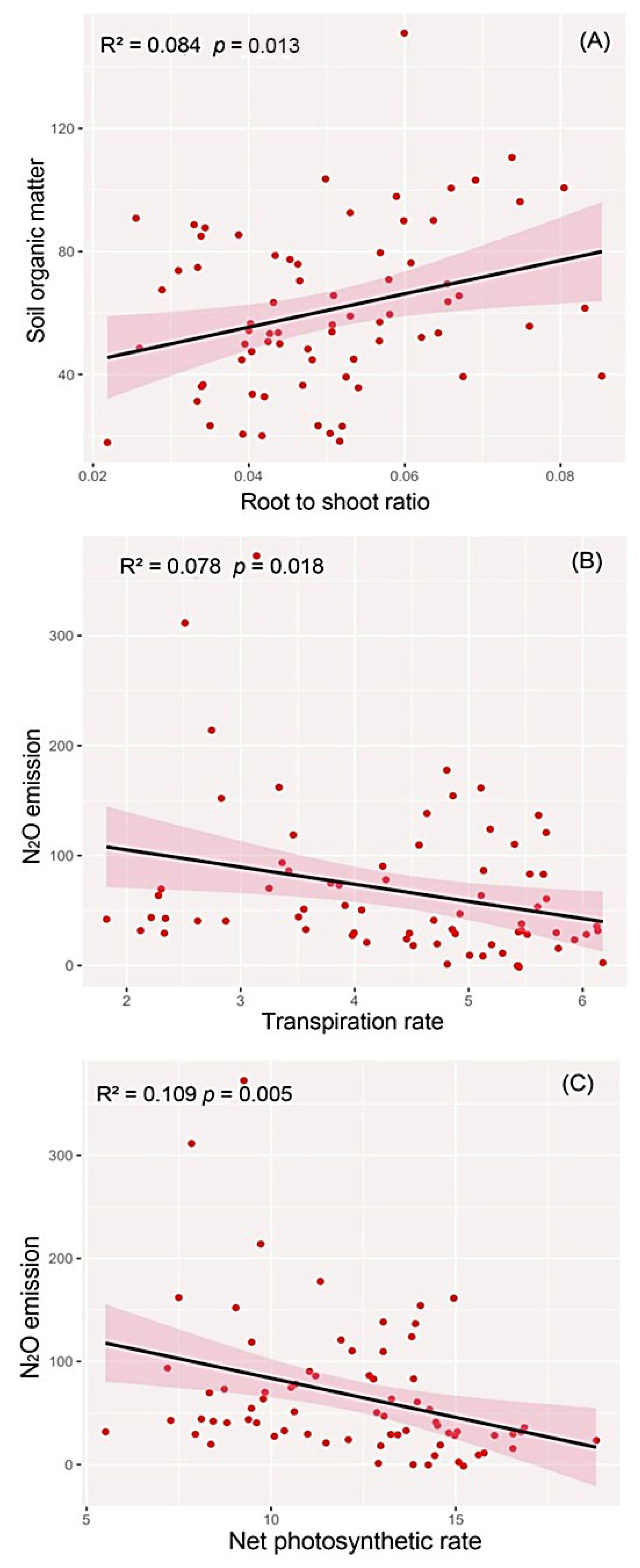
3.3. Relationships between the Invasiveness of H. vulgaris and the Wetland Microecosystem
4. Discussion
4.1. Effects of Genotypic Diversity and Flooding on H. vulgaris Growth
4.2. Effects of Genotypic Diversity and Flooding on the Wetland Microecosystem
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A. AFLP Protocols
References
- Callaway, R.M.; Aschehoug, E.T. Invasive plants versus their new and old neighbors: A mechanism for exotic invasion. Science 2000, 290, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Ren, M.X.; Zhang, Q.G. The relative generality of plant invasion mechanisms and predicting future invasive plants. Weed Res. 2009, 49, 449–460. [Google Scholar] [CrossRef]
- Warren, R.J.; Candeias, M.; Labatore, A.; Olejniczak, M.; Yang, L. Multiple mechanisms in woodland plant species invasion. J. Plant Ecol. 2019, 12, 201–209. [Google Scholar] [CrossRef]
- Williamson, M.H.; Fitter, A. The characters of successful invaders. Biol. Conserv. 1996, 78, 163–170. [Google Scholar] [CrossRef]
- Blossey, B.; Notzold, R. Evolution of increased competitive ability in invasive nonindigenous plants: A hypothesis. J. Ecol. 1995, 83, 887–889. [Google Scholar] [CrossRef]
- Davis, M.; Pelsor, M. Experimental support for a resource-based mechanistic model of invisibility. Ecol. Lett. 2001, 4, 421–428. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pysek, P. The ecology of invasions by animals and plants. London: Methuen. Prog. Phys. Geogr. 2007, 31, 659–666. [Google Scholar] [CrossRef]
- Thomsen, M.A.; D’Antonio, C.M.; Suttle, K.B.; Sousa, W.P. Ecological resistance, seed density and their interactions determine patterns of invasion in a California coastal grassland. Ecol. Lett. 2006, 9, 160–170. [Google Scholar] [CrossRef]
- Zheng, Y.L.; Burns, J.H.; Liao, Z.Y.; Li, W.-T.; Li, L. Nutrient fluctuation has different effects on a tropical invader in communities from the native and non-native range. Environ. Exp. Bot. 2020, 178, 104193. [Google Scholar] [CrossRef]
- Alotaibi, N.M.; Kenyon, E.J.; Cook, K.J.; Borger, L.; Bull, J.C. Low genotypic diversity and long-term ecological decline in a spatially structured seagrass population. Sci. Rep. 2019, 9, 18387. [Google Scholar] [CrossRef] [Green Version]
- Evans, S.M.; Verges, A.; Poore, A.G.B. Genotypic diversity and short-term response to shading stress in a threatened seagrass: Does low diversity mean low resilience? Front. Plant Sci. 2017, 8, 1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazard, C.; Johnson, D. Does genotypic and species diversity of mycorrhizal plants and fungi affect ecosystem function? New Phytol. 2018, 220, 1122–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.W.; Han, G.X. A review on the relationships between plant genetic diversity and ecosystem functioning. Chin. J. Plant Ecol. 2018, 42, 977–989. [Google Scholar] [CrossRef] [Green Version]
- Crawford, K.; Whitney, K. Population genetic diversity influences colonization success. Mol. Ecol. 2010, 9, 1253–1263. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.; Geng, Y.; Ou, X. Genetic diversity and invasion success of alien species: Where are we and where should we go? Biodivers. Sci. 2010, 18, 590–597. [Google Scholar] [CrossRef]
- Genton, B.J.; Shykoff, J.A.; Giraud, T. High genetic diversity in French invasive populations of common ragweed, Ambrosia artemisiifolia, as a result of multiple sources of introduction. Mol. Ecol. 2005, 14, 4275–4285. [Google Scholar] [CrossRef]
- Lavergne, S.; Molofsky, J. Increased genetic variation and evolutionary potential drive the success of an invasive grass. Proc. Natl. Acad. Sci. USA 2007, 104, 3883–3888. [Google Scholar] [CrossRef] [Green Version]
- Pappert, R.A.; Hamrick, J.L.; Donovan, L.A. Genetic variation in Pueraria lobata (Fabaceae), an introduced, clonal, invasive plant of the southeastern United States. Am. J. Bot. 2000, 87, 1240–1245. [Google Scholar] [CrossRef]
- Vellend, M.; Drummond, E.B.; Tomimatsu, H. Effects of genotype identity and diversity on the invasiveness and invisibility of plant populations. Oecologia 2010, 162, 371–381. [Google Scholar] [CrossRef]
- Wang, X.-Y.; Miao, Y.; Yu, S.; Chen, X.-Y.; Schmid, B. Genotypic diversity of an invasive plant species promotes litter decomposition and associated processes. Oecologia 2014, 174, 993–1005. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-Y.; Shen, D.-W.; Jiao, J.; Xu, N.-N.; Yu, S.; Zhou, X.-F.; Shi, M.-M.; Chen, X.-Y. Genotypic diversity enhances invasive ability of Spartina alterniflora. Mol. Ecol. 2012, 21, 2542–2551. [Google Scholar] [CrossRef] [PubMed]
- Perkins, L.B.; Johnson, D.W.; Nowak, R.S. Plant-induced changes in soil nutrient dynamics by native and invasive grass species. Plant Soil 2011, 345, 365–374. [Google Scholar] [CrossRef]
- Sun, F.; Ou, Q.; Yu, H.; Li, N.; Peng, C. The invasive plant Mikania micrantha affects the soil food web and plant-soil nutrient contents in orchards. Soil Biol. Biochem. 2019, 139, 107630. [Google Scholar] [CrossRef]
- Attermeyer, K.; Flury, S.; Jayakumar, R.; Fiener, P.; Steger, K.; Arya, V.; Wilken, F.; van Geldern, R.; Premke, K. Invasive floating macrophytes reduce greenhouse gas emissions from a small tropical lake. Sci. Rep. 2016, 6, 20424. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J. A global synthesis of the effects of biological invasions on greenhouse gas emissions. Glob. Ecol. Biogeogr. 2015, 24, 1351–1362. [Google Scholar] [CrossRef]
- Ai, Z.; Zhang, J.; Xin, Q.; Xue, S.; Liu, H. Soil nutrients influence the photosynthesis and biomass in invasive Panicum virgatum on the Loess Plateau in China. Plant Soil 2017, 418, 153–164. [Google Scholar] [CrossRef]
- He, Z.S.; He, W.M. Asymmetric climate warming does not benefit plant invaders more than natives. Sci. Total Environ. 2020, 742, 140624. [Google Scholar] [CrossRef]
- Nguyen, L.T.T.; Osanai, Y.; Anderson, I.C.; Bange, M.; Tissue, D.T.; Singh, B.K. Flooding and prolonged drought have differential legacy impacts on soil nitrogen cycling, microbial communities and plant productivity. Plant Soil 2018, 431, 371–387. [Google Scholar] [CrossRef]
- Zhou, W.; Chen, F.; Meng, Y.; Chandrasekaran, U.; Luo, X.; Yang, W.; Shu, K. Plant waterlogging/flooding stress responses: From seed germination to maturation. Plant Physiol. Biochem. 2020, 148, 228–236. [Google Scholar] [CrossRef]
- Casanova, M.T.; Brock, M.A. How do depth, duration and frequency of flooding influence the establishment of wetland plant communities? Plant Ecol. 2000, 147, 237–250. [Google Scholar] [CrossRef]
- Lei, T.; Cui, G.F.; Sha, H.F.; Li, F.L.; Yan, J.H. Diversity and community distribution characteristics of alien vascular plant species in wetlands of Beijing. J. Beijing For. Univ. 2010, 32, 51–57. [Google Scholar]
- Gao, L.X. Comparisons of morphological variation and cellular osmotic potential adjustment between invasive species Alternanthera philoxeroides and its native congener A. sessilis under different water treatments. Plant Sci. J. 2015, 33, 195–202. [Google Scholar]
- Chen, Y.; Zhou, Y.; Yin, T.F.; Liu, C.-X.; Luo, F.-L. The invasive wetland plant Alternanthera philoxeroides shows a higher tolerance to waterlogging than its native congener Alternanthera sessilis. PLoS ONE 2013, 8, e81456. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Wang, G.X.; Liu, J.E.; Xu, W.; Ren, L. Effects of waterlogging regulation on growth of Spartina Alterniflora. Res. Environ. Sci. 2011, 24, 1003–1007. [Google Scholar]
- Xie, B.H.; Wang, A.D.; Zhao, Y.J.; Shu, S.Y.; Song, J.B.; Han, G.X.; Guan, B.; Zhang, L.W. Effects of mowing plus waterlogging on germination and seedling growth of Spartina alterniflora. Chin. J. Ecol. 2018, 37, 417–423. [Google Scholar]
- Catford, J.; Downes, B.J.; Gippel, C.J.; Vesk, P.A. Flow regulation reduces native plant cover and facilitates exotic invasion in riparian wetlands. J. Appl. Ecol. 2011, 48, 432–442. [Google Scholar] [CrossRef]
- Dong, B.-C.; Wang, J.-Z.; Liu, R.-H.; Zhang, M.-X.; Luo, F.-L.; Yu, F.-H. Soil heterogeneity affects ramet placement of Hydrocotyle vulgaris. Plant Ecol. 2014, 8, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Chen, Q.; Dong, B.-C.; Yu, F.-H. Effects of vegetative propagule pressure on the establishment of an introduced clonal plant Hydrocotyle vulgaris. Sci. Rep. 2014, 4, 5507. [Google Scholar] [CrossRef] [Green Version]
- Miao, L.H.; Ji, M.C.; Wang, Y.Y.; Qiao, D.-D.; Chen, Y.-C. Study on invasion risk of Hydrocotyle vulgaris as an alien species in wetlands. J. Zhejiang Univ. Agric. Life Sci. 2011, 37, 425–431. [Google Scholar]
- Wang, M.-Z.; Bu, X.-Q.; Li, L.; Dong, B.-C.; Li, H.-L.; Yu, F.-H. Constraints on the evolution of phenotypic plasticity in the clonal plant Hydrocotyle vulgaris. J. Evol. Biol. 2018, 31, 1006–1017. [Google Scholar] [CrossRef]
- Liu, L.; Alpert, P.; Dong, B.C.; Li, J.-M.; Yu, F.-H. Combined effects of soil heterogeneity, herbivory and detritivore on growth of the clonal plant Hydrocotyle vulgaris. Plant Soil 2017, 421, 429–437. [Google Scholar] [CrossRef]
- Qin, T.; Guan, Y.; Quan, H.; Dong, B.; Luo, F.; Zhang, M.; Li, H.; Yu, F. Growth traits of the exotic plant Hydrocotyle vulgaris and the evenness of resident plant communities are mediated by community age, not species diversity. Weed Res. 2019, 59, 377–386. [Google Scholar] [CrossRef]
- Li, Q.-W.; Zhang, X.-Y.; Gao, J.-Q.; Song, M.-H.; Liang, J.-F.; Yue, Y. Effects of N addition frequency and quantity on Hydrocotyle vulgaris growth and greenhouse gas emissions from wetland microcosms. Sustainability 2019, 11, 1520. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Mo, K.; Chen, Q.; Zhang, J.; Xia, J.; Lin, Y. An experimental study on potential changes in plant community evapotranspiration due to the invasion of Hydrocotyle vulgaris. J. Hydro Environ. Res. 2020, 30, 63–70. [Google Scholar] [CrossRef]
- Wang, M.-Z.; Li, H.-L.; Li, J.-M.; Yu, F.-H. Correlations between genetic, epigenetic and phenotypic variation of an introduced clonal herb. Heredity 2020, 124, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; Van De Lee, T.; Hornes, M.; Friters, A.; Pot, J.; Paleman, J.; Kuiper, M.; et al. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef] [Green Version]
- Guy-Haim, T.; Lyons, D.A.; Kotta, J.; Ojaveer, H.; Queiros, A.M.; Chatzinikolaou, E.; Arvanitidis, C.; Como, S.; Magni, P.; Blighy, A.J.; et al. Diverse effects of invasive ecosystem engineers on marine biodiversity and ecosystem functions: A global review and meta-analysis. Glob. Change Biol. 2018, 24, 906–924. [Google Scholar] [CrossRef]
- Han, F.; Ren, L.; Zhang, X.C. Effect of biochar on the soil nutrients about different grasslands in the Loess Plateau. Catena 2016, 137, 554–562. [Google Scholar] [CrossRef]
- Zhang, H.-Y.; Goncalves, P.; Copeland, E.; Qi, S.-S.; Dai, Z.-C.; Li, G.-L.; Wang, C.; Du, D.-L.; Thomas, T. Invasion by the weed Conyza canadensis alters soil nutrient supply and shifts microbiota structure. Soil Biol. Biochem. 2020, 143, 107739. [Google Scholar] [CrossRef]
- Michel, L.; Andy, H. Partitioning selection and complementarity in biodiversity experiments. Nature 2001, 412, 72–77. [Google Scholar]
- Engelhardt, K.A.M.; Ritchie, M.E. The effect of aquatic plant species richness on wetland ecosystem processes. Ecology 2002, 83, 2911–2924. [Google Scholar] [CrossRef]
- Yang, X.; Wang, X.P.; Qu, Y.B.; Wu, M.; Wang, J.L.; Wang, Z.L.; Wang, S.Q.; Zhao, N.X.; Gao, Y.B. Comparing the effects of companion species diversity and the dominant species (Stipa grandis) genotypic diversity on the biomass explained by plant functional trait. Ecol. Eng. 2019, 136, 17–22. [Google Scholar] [CrossRef]
- Wang, J.; Song, C.; Miao, Y.; Meng, H. Greenhouse gas emissions from southward transplanted wetlands during freezing-thawing periods in Northeast China. Wetlands 2013, 33, 1075–1081. [Google Scholar] [CrossRef]
- Begum, N.; Guppy, C.; Herridge, D.; Schwenke, G. Influence of source and quality of plant residues on emissions of N2O and CO2 from a fertile, acidic black vertisol. Biol. Fertil. Soils 2014, 50, 499–506. [Google Scholar] [CrossRef]
- Gao, J.-Q.; Duan, M.-Y.; Zhang, X.-Y.; Li, Q.-W.; Yu, F.-H. Effects of frequency and intensity of drying-rewetting cycles on Hydrocotyle vulgaris growth and greenhouse gas emissions from wetland microcosms. Catena 2018, 164, 44–49. [Google Scholar] [CrossRef]
- Drummond, E.B.M.; Vellend, M. Genotypic diversity effects on the performance of Taraxacum officinale populations increase with time and environmental favorability. PLoS ONE 2012, 7, e30314. [Google Scholar] [CrossRef]
- Tomimatsu, H.; Nakano, K.; Yamamoto, N.; Suyama, Y. Effects of genotypic diversity of Phragmites australis on primary productivity and water quality in an experimental wetland. Oecologia 2014, 175, 163–172. [Google Scholar] [CrossRef]
- Zhang, J.L.; Cao, K.F. The effect of irradiance on photosynthetic capacity, heat dissipation, and antioxidants of seedlings of two tropical rain forest species. Acta Phytoecol. Sin. 2002, 26, 639–646. [Google Scholar]
- Crawford, K.M.; Rudgers, J.A. Plant species diversity and genetic diversity within a dominant species interactively affect plant community biomass. J. Ecol. 2012, 100, 1512–1521. [Google Scholar] [CrossRef]
- Guo, X.; Xu, Z.-W.; Li, M.-Y.; Ren, X.-H.; Liu, J.; Guo, W.-H. Increased soil moisture aggravated the competitive effects of the invasive tree Rhus typhina on the native tree Cotinus coggygria. BMC Ecol. 2020, 20, 17. [Google Scholar] [CrossRef] [Green Version]
- Hooper, D.U.; Dukes, J.S. Overyielding among plant functional groups in a long-term experiment. Ecol. Lett. 2003, 7, 95–105. [Google Scholar] [CrossRef]
- Huang, M.J.; Liu, X.; Cadotte, W.M.; Zhou, S. Functional and phylogenetic diversity explain different components of diversity effects on biomass production. Oikos 2020, 129, 1185–1195. [Google Scholar] [CrossRef]
- Cardinale, B.J. Biodiversity improves water quality through niche partitioning. Nature 2011, 472, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Reusch, T.B.H.; Ehlers, A.; Hammerli, A.; Worm, B. Ecosystem recovery after climatic extremes enhanced by genotypic diversity. Proc. Natl. Acad. Sci. USA 2005, 102, 2826–2831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhang, C.B.; Chen, T.; Li, W.H. From selection to complementarity: The shift along the abiotic stress gradient in a controlled biodiversity experiment. Oecologia 2013, 171, 227–235. [Google Scholar] [CrossRef]
- Forrester, D.I. The spatial and temporal dynamics of species interactions in mixed species forests: From pattern to process. For. Ecol. Manag. 2014, 312, 282–292. [Google Scholar] [CrossRef]
- Hughes, A.R.; Stachowicz, J.J. Seagrass genotypic diversity increases disturbance response via complement and dominance. J. Ecol. 2011, 99, 445–453. [Google Scholar]
- Shen, J.; Ren, H.; Xin, X.; Xu, B.; Gao, Y.B.; Zhao, N.X. Leymus chinensis genotypic diversity increases the response of populations to disturbance. Acta Ecol. Sin. 2015, 35, 7682–7689. [Google Scholar]
- Yang, Q.Q. The Phenotypic Plasticity of Water and the Forecast of Evolution of the Invasive Plant Hydrocotyle vulgaris. Master’s Thesis, Zhejiang A&F University, Hangzhou, China, 2013. [Google Scholar]
- Vitousek, P.M.; Walker, L.R.; Whiteaker, L.D.; Mueller-Dombois, D.; Matson, P.A. Biological invasion by Myrica faya alters ecosystem development in Hawaii. Science 1987, 238, 802–804. [Google Scholar] [CrossRef]
- Tyrer, S.J.; Hild, A.L.; Munn, M.L.C. Establishment of native species in soils from Russian Knapweed (Acroptilon repens) invasions. Rangel. Ecol. Manag. 2007, 60, 604–612. [Google Scholar] [CrossRef]
- Zhang, X.; Gao, G.; Wu, Z.; Wen, X.; Zhong, H.; Zhong, Z.; Yang, C.; Bian, F.; Gai, X. Responses of soil nutrients and microbial communities to intercropping medicinal plants in moso bamboo plantations in subtropical China. Environ. Sci. Pollut. Res. Int. 2020, 27, 2301–2310. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.H.; Tao, X.U.; Fei, H.U.; Shoushan, H. Allelopathy under environmental stress and its induced mechanism. Acta Ecol. Sin. 2000, 20, 849–854. [Google Scholar]
- Peng, S.L. Research advances in the interactions between allelopathy and soil in exotic invasions. Ecol. Environ. 2007, 16, 644–648. [Google Scholar]
- Morris, K.A.; Stark, J.M.; Bugbee, B.; Norton, J.M. The invasive annual cheatgrass releases more nitrogen than crested wheatgrass through root exudation and senescence. Oecologia 2016, 181, 971–983. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.S.; Cai, W.F.; Kohl, K.; Nishimoto, R.K. Plant stress and allelopathy. Acs Symp. Ser. 1995, 582, 142–147. [Google Scholar]
- Liu, X.W.; Zhou, Y.L.; Qi, C.M.; Li, Y.; Wang, Q.X.; Guo, M.X.; Yan, D.D.; Cao, A.C. Effects of Mikania micrantha invasion on soil nutrient contents and enzyme activities. Ecol. Environ. Sci. 2012, 21, 1960–1965. [Google Scholar]
- Zhao, J.; Cheng, C.; Gu, X.Y.; Liu, B. Effects of root exudates from Mirabilis Jalapa on soil microenvironment. J. Henan Norm. Univ. Nat. Sci. Ed. 2014, 42, 95–99. [Google Scholar]
- Chen, H.; Li, Y.; Bo, L.; Chen, J.; Wu, J. Impacts of exotic plant invasions on soil biodiversity and ecosystem processes. Biodivers. Sci. 2005, 13, 555–565. [Google Scholar] [CrossRef]
- Dassonville, N.; Vanderhoeven, S.; Vanparys, V.; Hayez, M.; Gruber, W.; Meerts, P. Impacts of alien invasive plants on soil nutrients are correlated with initial site conditions in NW Europe. Oecologia 2008, 157, 131–140. [Google Scholar] [CrossRef]
- Santoro, R.; Jucker, T.; Carranza, M.L.; Acosta, A. Assessing the effects of Carpobrotus invasion on coastal dune soils. Does the nature of the invaded habitat matter? Community Ecol. 2011, 12, 234–240. [Google Scholar] [CrossRef]
- Yang, H.S.; Tain, K.; Yao, X.; Cao, P.L. The impacts of dam impoundment on the distribution patterns of soil organic matter and total nitrogen in Lashihai plateau wetland. Chin. J. Ecol. 2015, 34, 162–167. [Google Scholar]
- Tian, K.; Chang, F.L.; Lu, M. Impacts of human disturbances on organic carbon carbon and nitrogen in Nanpahai wetlands, northwest Yunnan. Acta Pedol. Sin. 2004, 41, 681–686. [Google Scholar]
- Gao, C.; Zhang, T.L.; Wu, W.D. Phosphorus sorption and release of passy soils: Effect of alternation of oxidized and reduced conditions. Acta Pedol. Sin. 2002, 39, 542–549. [Google Scholar]
- Lenhart, K.; Behrendt, T.; Greiner, S.; Steinkamp, J.; Well, R.; Giesemann, A.; Keppler, F. Nitrous oxide effluxes from plants as a potentially important source to the atmosphere. New Phytol. 2019, 221, 1398–1408. [Google Scholar] [CrossRef]
- Schützenmeister, K.; Meurer, K.H.E.; Gronwald, M.; Hartmann, A.B.D.; Gansert, D.; Jungkunst, H.F. N2O emissions from plants are reduced under photosynthetic activity. Plant Environ. Interact. 2020, 1, 48–56. [Google Scholar] [CrossRef]
- Hu, H.X.; Yuan, D.; Zeng, J.R.; Yao, J.Z.; He, X.D.; Qin, S.P.; Hu, C.S.; Zhou, S.G. Advances in plant nitrous oxide (N2O) emissions. Chin. J. Eco. Agric. 2021, 29, 345–354. [Google Scholar]
- Chen, Q.W.; Zheng, T.; Li, L. Influence Factors of N2O production and accumulation during denitrification. J. Anhui Agric. Sci. 2011, 39, 11667–11668. [Google Scholar]
- Qi, X.; Zhang, S.; Lin, F.; Zhang, L.L.; Wang, H. Effect of Flaveria bidentis invasion on plant community and soil microbial community of different in invaded soil. Ecol. Environ. Sci. 2019, 39, 8472–8482. [Google Scholar]
- Sun, Z.G.; Liu, J.S.; Yu, J.B.; Yi, H.P. Ammonia volatilization, nitrous oxide emission processes in wetland soil and affecting factors. Wetl. Sci. 2008, 6, 429–439. [Google Scholar]
- Yan, X.Y.; Shi, S.L.; Du, L.J.; Xing, G.X. N2O emission from paddy soil as affected by water regime. Acta Pedol. Sin. 2000, 37, 483–489. [Google Scholar]
- Cheng, X.L.; Peng, R.H.; Chen, J.Q.; Luo, Y.; Zhang, Q.; An, S.; Chen, J.; Li, B. CH4 and N2O emissions from Spartina alterniflora and Phragmites australis in experimental mesocosms. Chemosphere 2007, 68, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.N.; Wang, X.K.; OuYang, Z.Y. Effects of vascular plants on methane emissions from natural wetlands. Acta Ecol. Sin. 2005, 25, 3375–3382. [Google Scholar]
- Mer, J.L.; Roger, P. Production, oxidation, emission and consumption of methane by soils: A review. Eur. J. Soil Biol. 2001, 37, 25–50. [Google Scholar] [CrossRef]
- Huang, Q.Q.; Shen, Y.D.; Li, X.X.; Cheng, H.T.; Song, X.; Fan, Z.W. Research progress on the distribution and invasiveness of alien invasive plants in China. Ecol. Environ. Sci. 2012, 21, 977–985. [Google Scholar]
- Wei, Z.S.; Chen, X.W.; Yang, D.L.; Huangfu, C. Comparison of gas exchange characteristics between two invasive plants and a co-occurring native plant in Liaoning Province. Chin. J. Eco. Agric. 2017, 25, 975–982. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotypic Diversity (G) | Flooding (F) | G × F | ||||
|---|---|---|---|---|---|---|
| Indexes | F | p | F | p | F | p |
| Growth traits | ||||||
| Internode length | 0.84 | 0.437 | 8.84 | 0.004 | 0.71 | 0.496 |
| Petiole length | 0.39 | 0.677 | 0.89 | 0.034 | 0.50 | 0.609 |
| Number of ramets | 0.56 | 0.574 | 2.83 | <0.001 | 0.96 | 0.388 |
| Leaf area | 8.41 | 0.005 | 3.56 | 0.034 | 4.80 | 0.011 |
| Total biomass | 5.48 | 0.060 | 0.11 | 0.740 | 1.21 | 0.304 |
| Leaf biomass | 4.78 | 0.110 | 0.03 | 0.846 | 1.02 | 0.366 |
| Internode biomass | 5.68 | 0.055 | 8.22 | 0.006 | 1.18 | 0.314 |
| Petiole biomass | 3.58 | 0.096 | 1.89 | 0.174 | 2.47 | 0.092 |
| Root biomass | 1.37 | 0.261 | 45.46 | <0.001 | 1.55 | 0.220 |
| Root to shoot ratio | 2.43 | 0.033 | 90.41 | 0.097 | 8.01 | 0.001 |
| Photosynthetic physiological traits | ||||||
| Net photosynthetic rate | 30.31 | <0.001 | 75.87 | <0.001 | 6.70 | 0.002 |
| Stomatal conductance | 30.46 | 0.001 | 80.51 | <0.001 | 5.51 | 0.005 |
| Intercellular CO2 concentration | 4.73 | 0.011 | 36.17 | <0.001 | 4.03 | 0.020 |
| Transpiration rate | 24.23 | <0.001 | 8.50 | 0.004 | 1.71 | 0.185 |
| Greenhouse gas emissions | ||||||
| Carbon dioxide cumulative emissions (CO2) | 0.12 | 0.886 | 0.44 | 0.509 | 2.43 | 0.097 |
| Methane flux cumulative emissions (CH4) | 0.71 | 0.499 | 1.15 | 0.289 | 0.27 | 0.767 |
| Nitrous oxide cumulative emissions (N2O) | 3.27 | 0.044 | 11.89 | 0.001 | 2.47 | 0.093 |
| Soil properties | ||||||
| Soil organic matter (SOM) | 15.97 | 0.000 | 4.58 | 0.036 | 3.29 | 0.043 |
| Soil total nitrogen (TN) | 1.25 | 0.295 | 9.03 | 0.004 | 0.84 | 0.435 |
| Soil total phosphorus (TP) | 1.81 | 0.172 | 4.20 | 0.044 | 1.10 | 0.338 |
| Genotypic Diversity (G) | Flooding (F) | G × F | ||||
|---|---|---|---|---|---|---|
| F | p | F | p | F | p | |
| Complementary effect | ||||||
| Internode length | 9.57 | 0.006 | 21.94 | <0.001 | 5.52 | 0.029 |
| Petiole length | 3.22 | 0.088 | 12.13 | 0.002 | 4.363 | 0.050 |
| Leaf area | 11.06 | 0.003 | 116.08 | <0.001 | 50.76 | <0.001 |
| Number of plants | 25.84 | <0.001 | 41.20 | <0.001 | 16.24 | 0.001 |
| Internode biomass | 30.15 | <0.001 | 2.78 | 0.112 | 23.31 | <0.001 |
| Petiole biomass | 23.54 | <0.001 | 74.33 | <0.001 | 55.08 | <0.001 |
| Root biomass | 59.57 | <0.001 | 0.01 | 0.909 | 17.92 | <0.001 |
| Leaf biomass | 31.75 | <0.001 | 35.92 | <0.001 | 20.42 | <0.001 |
| Total biomass | 34.58 | <0.001 | 39.60 | <0.001 | 36.94 | <0.001 |
| Selection effect | ||||||
| Internode length | 8.68 | 0.008 | 15.29 | 0.001 | 5.35 | 0.031 |
| Petiole length | 2.71 | 0.116 | 8.11 | 0.01 | 3.61 | 0.072 |
| Leaf area | 9.78 | 0.005 | 69.67 | <0.001 | 36.67 | <0.001 |
| Number of plants | 15.75 | 0.001 | 22.34 | <0.001 | 11.11 | 0.003 |
| Internode biomass | 16.13 | 0.001 | 1.81 | 0.194 | 9.16 | 0.007 |
| Petiole biomass | 16.56 | 0.001 | 41.44 | <0.001 | 33.22 | <0.001 |
| Root biomass | 30.11 | <0.001 | 0.08 | 0.783 | 7.40 | 0.013 |
| Leaf biomass | 22.43 | <0.001 | 20.15 | <0.001 | 13.26 | 0.002 |
| Total biomass | 22.46 | <0.001 | 21.38 | <0.001 | 20.66 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, J.-F.; Sun, K.; Qin, T.-J.; Bu, X.-Q.; Wang, M.-Z.; Li, H.-L. Genotypic Diversity Improves Photosynthetic Traits of Hydrocotyle vulgaris and Alters Soil Organic Matter and N2O Emissions of Wetland Microecosystems. Water 2022, 14, 872. https://doi.org/10.3390/w14060872
Cai J-F, Sun K, Qin T-J, Bu X-Q, Wang M-Z, Li H-L. Genotypic Diversity Improves Photosynthetic Traits of Hydrocotyle vulgaris and Alters Soil Organic Matter and N2O Emissions of Wetland Microecosystems. Water. 2022; 14(6):872. https://doi.org/10.3390/w14060872
Chicago/Turabian StyleCai, Jing-Fang, Kai Sun, Tian-Jian Qin, Xiang-Qi Bu, Mo-Zhu Wang, and Hong-Li Li. 2022. "Genotypic Diversity Improves Photosynthetic Traits of Hydrocotyle vulgaris and Alters Soil Organic Matter and N2O Emissions of Wetland Microecosystems" Water 14, no. 6: 872. https://doi.org/10.3390/w14060872
APA StyleCai, J. -F., Sun, K., Qin, T. -J., Bu, X. -Q., Wang, M. -Z., & Li, H. -L. (2022). Genotypic Diversity Improves Photosynthetic Traits of Hydrocotyle vulgaris and Alters Soil Organic Matter and N2O Emissions of Wetland Microecosystems. Water, 14(6), 872. https://doi.org/10.3390/w14060872