
Study on the Algae Lysis Method of White Rot Fungi Algae Control System



Abstract
:1. Introduction
2. Materials and Methods
2.1. Algal Cultures
2.2. Fungal Strains and Culture
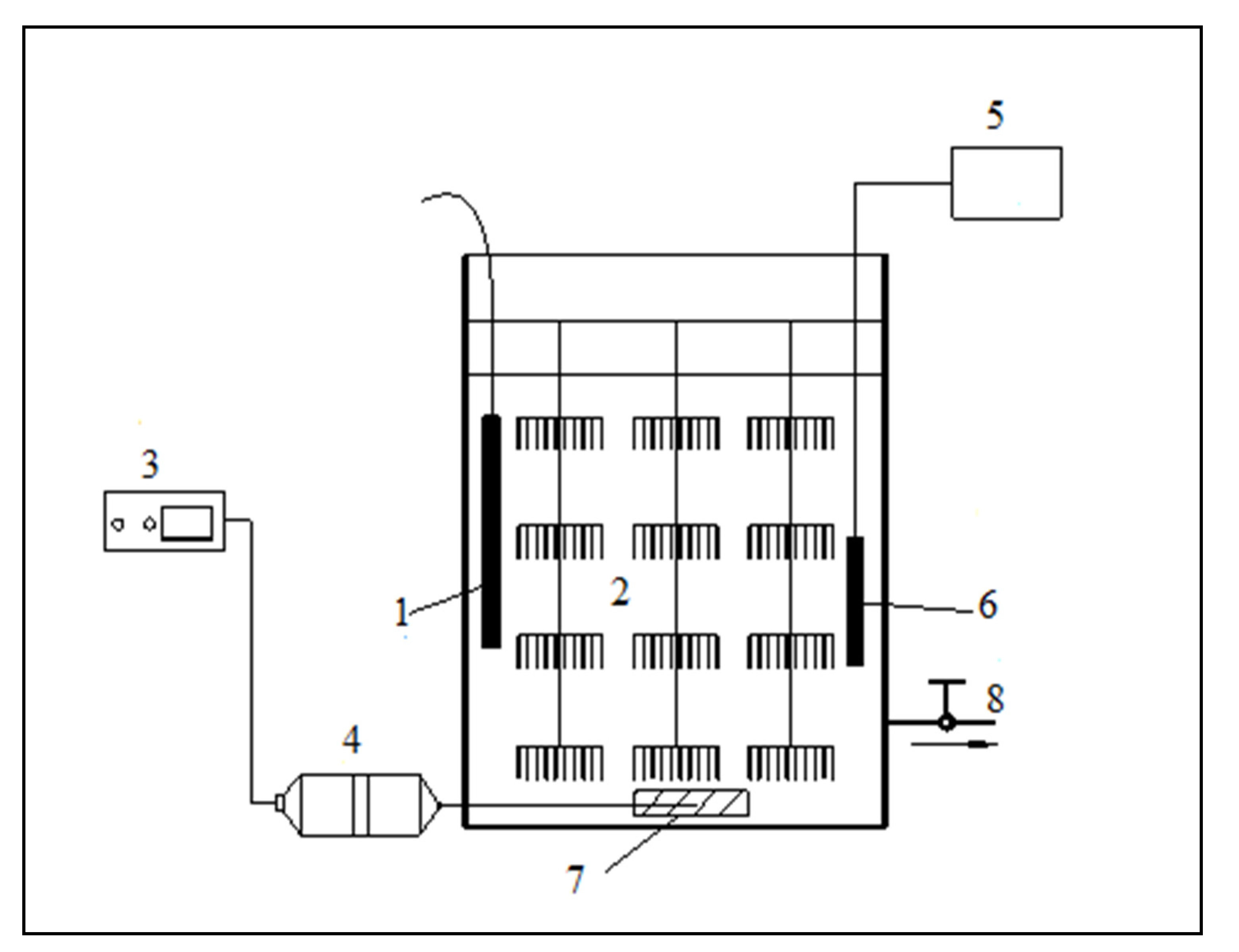
2.3. Experimental Device
2.4. Experimental Methods
2.5. Experimental Measurement Index
2.5.1. Measurement of Chlorophyll-a
2.5.2. Determination of Dehydrogenase Activity in Algal Cells
- (1)
- Firstly, 15 mL of the solution was taken to be tested and was filtered through a 0.25 μm microporous filter membrane. After suction filtration, the filter membrane was placed into a centrifuge tube, and 2 mL of Tris-HCl (pH = 7.5) and 1 mL of 0.8% (w/v) TTC were successively added until the algal cells were thoroughly mixed. Finally, the sample was placed in a dark place and the reaction was carried out in a constant-temperature water bath at 35 ± 1 °C;
- (2)
- The reaction was terminated after 60 min by adding 3–6 drops of 40% (w/w) formaldehyde solution to the centrifuge tube;
- (3)
- The triphenyl formazone (TPF) was extracted by adding 4 mL of acetone and 5 mL of petroleum ether, and shaken with a vortex mixer for 3 min. All the red extract was placed on the upper layer, and the absorbance of the petroleum ether extract was measured at 492 nm (using petroleum ether as a reference). The dehydrogenase activity (DHA) of algal cells was calculated according to the TPF value. DHA was represented by the TPF produced within 1 h in 1 mL of the test solution (μg TPF/mL.h). All experiments were repeated 3 times.
2.5.3. Determination of Soluble Protein Content
- (1)
- Preparation of 100 mg/L Coomassie brilliant blue reagent: Firstly, 100 mg of G-250 Coomassie brilliant blue was accurately weighed and dissolved in 50 mL of 95% ethanol solution, then 100 mL of 85% (w/v) H3PO4 solution was added, using distilled water to make the total volume of solution 1 L.
- (2)
- Preparation of 0.1 mg/mL standard protein: 10 mg of bovine serum albumin was accurately weighed, then dissolved in 100 mL of distilled water to prepare a 0.1 mg/mL standard protein solution, and stored at 4 °C for later use.
- (3)
- Preparation of 0.05 mol/L phosphate buffer solution with pH value of 7.8: 1.52 g of KH2PO4·2H2O and 14.33 g of K2HPO4·12H2O were accurately weighed, dissolved in distilled water, and made up to 1 L.
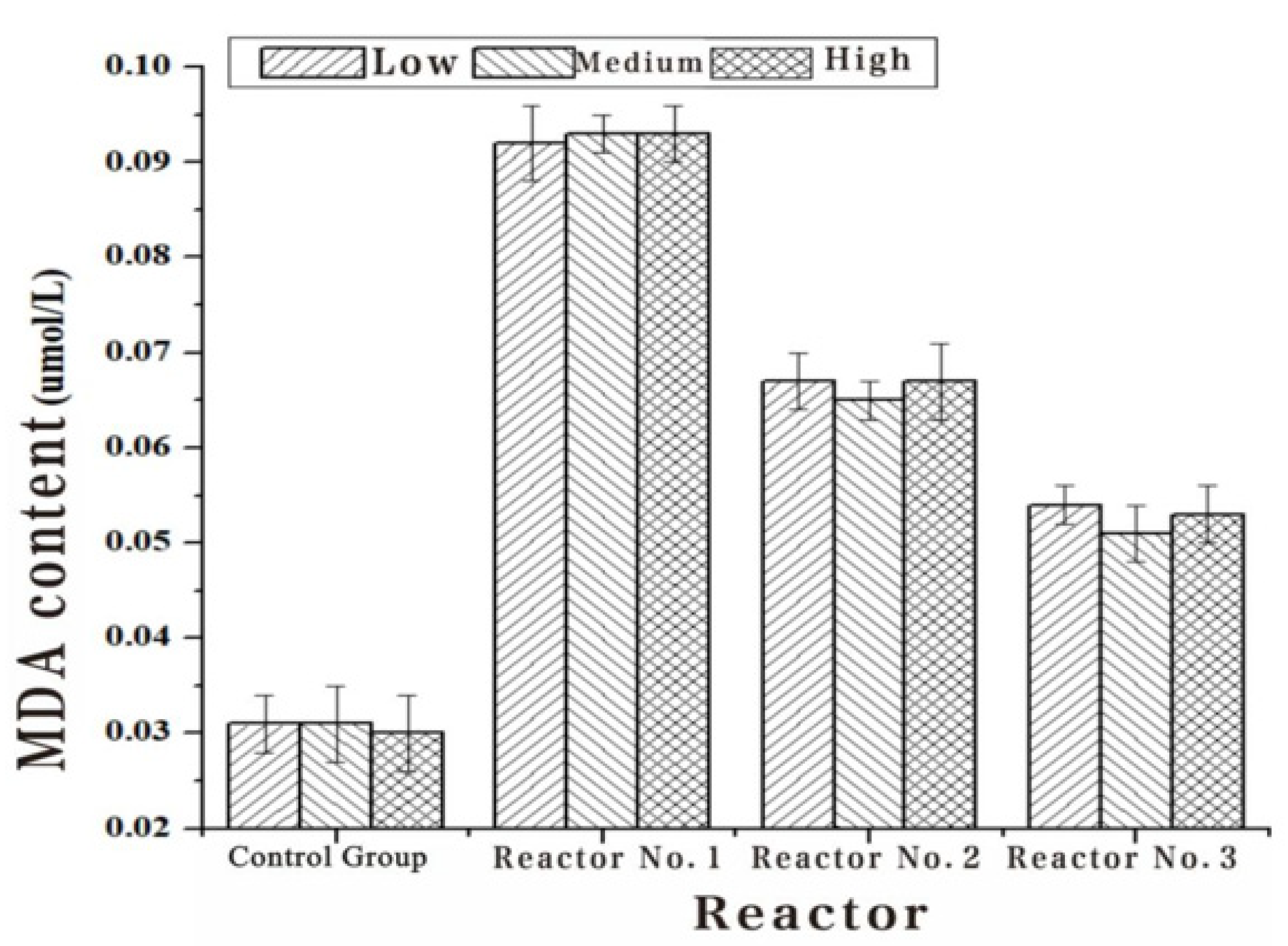
2.5.4. Determination of Malondialdehyde Content
- (1)
- Firstly, 2 mL of crude enzyme solution was drawn (2 mL of distilled water for the control group), 2 mL of 0.5% (w/v) trichloroacetic acid solution was added, and then 2 mL of 0.67% (w/v) TBA solution was added. This solution was shaken well;
- (2)
- The solution was put in a boiling water bath and boiled for 15 min to make it fully react;
- (3)
- After 15 min., the solution was immediately taken out of the test tube and put into cold water for cooling. After cooling, the solution was centrifuged at 4000 r/min for 20 min. The supernatant absorbance was measured at 600 nm, 532 nm and 450 nm wavelengths, respectively. The MDA concentration and content were calculated according to Equations (2) and (3), as follows:
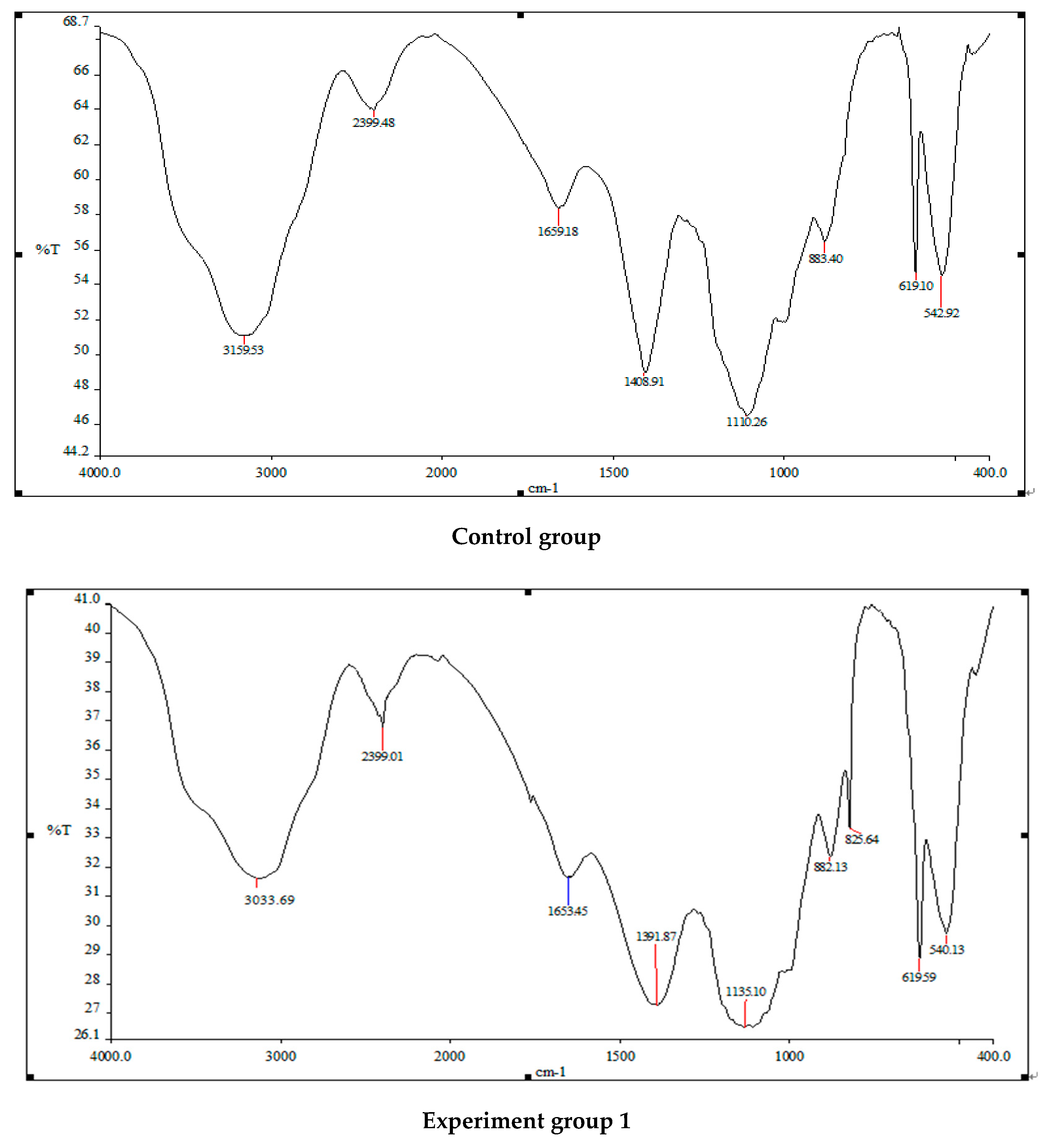
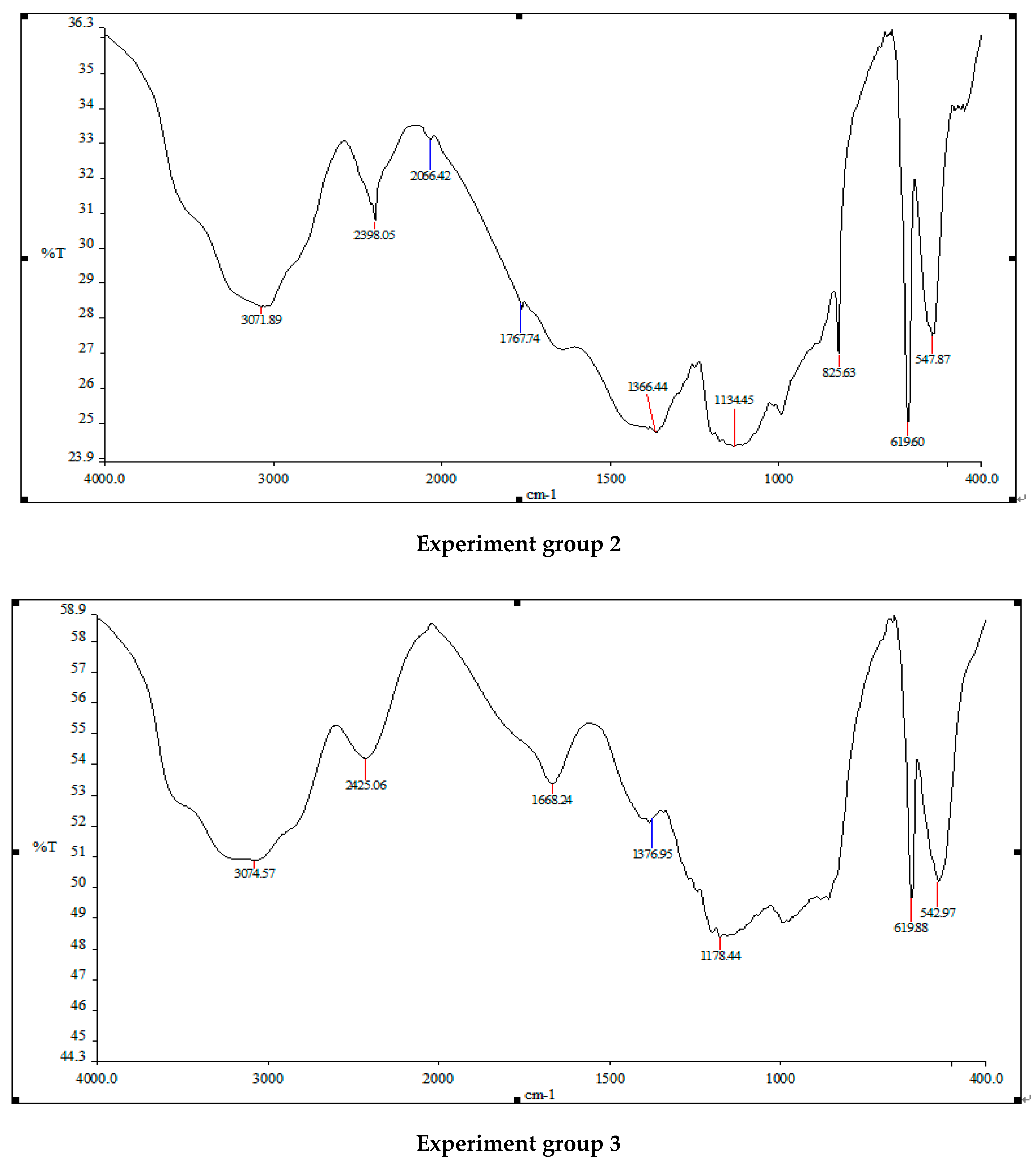
2.5.5. Fourier Transform Infrared Absorption Spectrometer Detection
2.6. Statistical Analysis Methods
3. Results
3.1. Effect of Three Different Ways of Adding White Rot Fungi on the Treatment of Turpin
3.2. Changes in Algal Cell Functional Groups before and after Treatment with Three Different Addition Methods of White Rot Fungi
4. Discussion
4.1. Effects of Three Addition Methods of White Rot Fungi on the Physiological Metabolism of Algae
4.2. The Effect of Three Addition Methods of White Rot Fungi on the Functional Groups of Algae
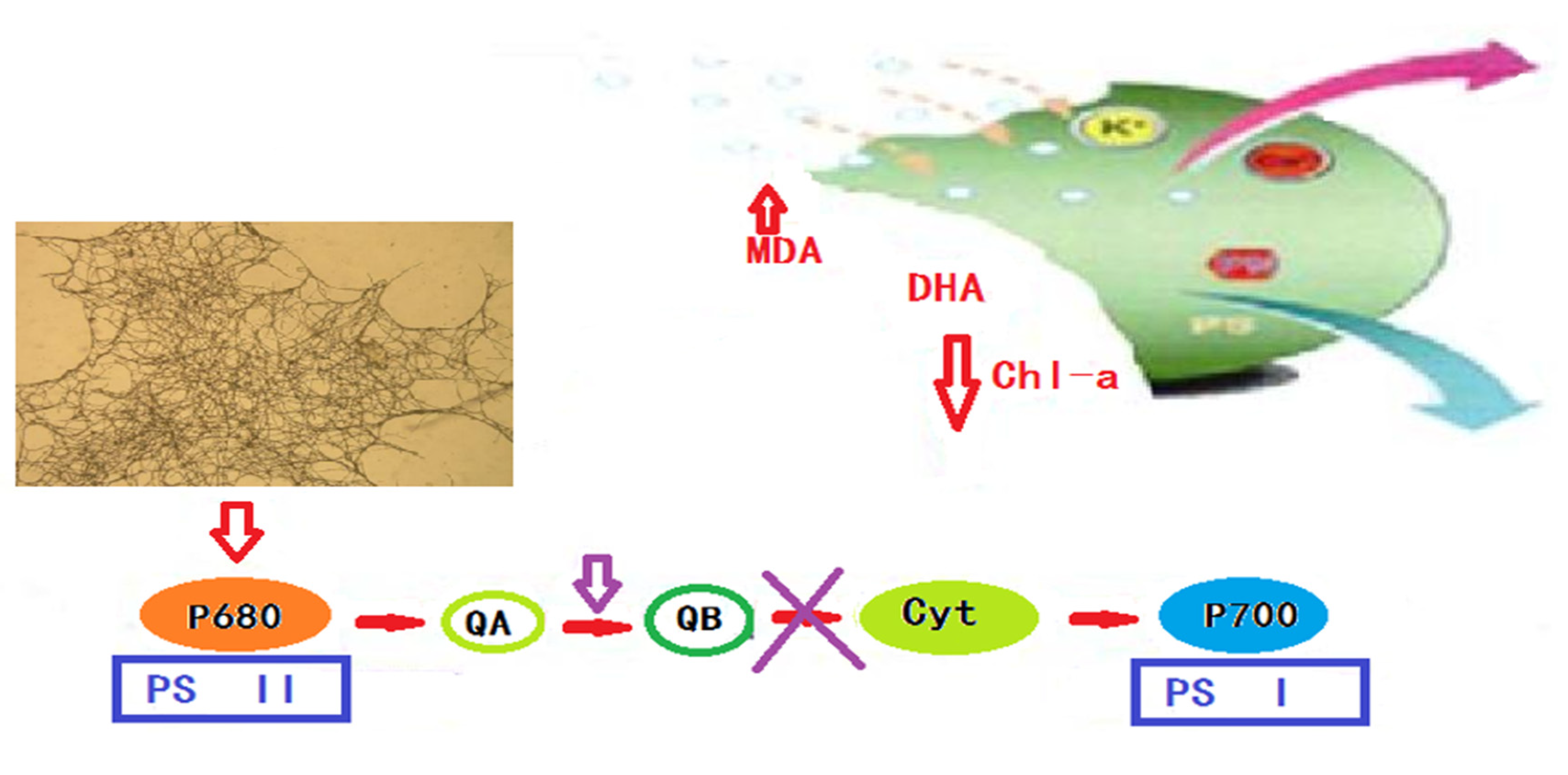
4.3. Discussion on the Algae Lysis Mechanism of White Rot Fungi
5. Conclusions
- (1)
- Based on the above physiological and biochemical indicators, and the results of the Fourier infrared spectroscopy experiment, it is found that the method of algae lysis for white rot fungi may mainly occur through direct algae lysis, supplemented by indirect algae lysis, and the algae-lysing substance contains a large amount of protein substances, and, at the same time, contains a small amount of polysaccharides or lipids.
- (2)
- The algae-dissolving mechanism of the white rot fungi algae control system is as follows: after the white rot fungi make contact with the algal cells, they begin to directly destroy the integrity of the algal cell membrane, and then the algae-lysing substances secreted by the white rot fungi pass through the damaged cell membrane and gradually destroy the algal body. A large number of photosynthetic pigments inhibit the normal physiological metabolism of the algal cells and the occurrence of photosynthesis. Finally, under the continuous action of white rot fungi, the nucleic acid substances, nutrients and genetic materials in algal cells are also severely damaged, which inhibits or kills the algal cells.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marsalek, B.; Jancula, D.; Marsalkova, E. Multimodal Action and Selective Toxicity of Zerovalent Iron Nanoparticles against Cyanobacteria. Environ. Sci. Technol. 2012, 46, 2316–2323. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.R.; Hong, Y.; Xie, X. Research progress on prevention and control methods of algal blooms and new technologies for inactivation and capture. Environ. Sci. Technol. 2021, 44, 171–185. [Google Scholar]
- Xie, S.L.; Wang, J.; Liu, Q. Research Progress of Plant Allelopathic Algae Control. J. Shanxi Univ. Nat. Sci. Ed. 2017, 40, 652–660. [Google Scholar]
- Ye, X.; Yi, C.; Li, T.L.; Yang, L.; Zhuang, X.C. Current status of research on the application of microbial agents in the water environment. Sichuan Environ. 2021, 40, 240–245. [Google Scholar]
- Yang, L.L.; Pan, W.B.; Li, Y. Composition and algal lysis mechanism of an active algae solubilizing substance from a strain of algae solubilizing bacteria. Environ. Sci. Technol. 2010, 33, 72–75. [Google Scholar]
- He, J.Y.; Pan, W.B.; Lin, M. Effects of algicidal bacteria on algal community structure in eutrophic water. Environ. Pollut. Prev. 2008, 30, 70–74. [Google Scholar]
- Mu, R.; Fan, Z.; Pei, H. Isolation and algae-lysing characteristics of the algicidal bacterium B5. J. Environ. Sci. 2007, 19, 1336–1340. [Google Scholar] [CrossRef]
- Ren, J.; Ma, Q.W.; Huang, H.H. Oxidative degradation of microcystin-LR by combination of UV/H2O2. Fresen Environ. Bull. 2010, 12, 3037–3044. [Google Scholar]
- Hu, H.J.; Wei, Y.X. Freshwater Algae of China—Systems, Taxonomy and Ecology; Science Press: Beijing, China, 2006; pp. 67–68. [Google Scholar]
- Wu, G.; Xi, Y.; Zhao, Y.J. The latest progress in the research of algicidal bacteria. Environ. Sci. Res. 2002, 15, 43–46. [Google Scholar]
- Kim, J.D.; Kim, J.Y.; Park, J.K.; Lee, C.G. Selective Control of the Prorocentrum minimum Harmful Algal Blooms by a Novel Algal-Lytic Bacterium Pseudoalteromonas haloplanktis AFMB-008041. Mar. Biotechnol. 2009, 11, 463–472. [Google Scholar] [CrossRef]
- Kim, M.J.; Jeong, S.Y.; Lee, S.J. Isolation, identification, and algicidal activity of marine bacteria against Cochlodinium polykrikoides. J. Appl. Phycol. 2008, 20, 1069–1078. [Google Scholar] [CrossRef]
- Lawton, L.A.; Welgamage, A.; Manage, P.M.; Edwards, C. Novel bacterial strains for the removal of microcystins from drinking water. Water Sci. Technol. 2011, 63, 1137–1142. [Google Scholar] [CrossRef] [PubMed]
- Lemes, G.A.F.; Kersanach, R.; Pinto, L.D.; Dellagostin, O.A.; Yunes, J.S.; Matthiensen, A. Biodegradation of microcystins by aquatic Burkholderia sp from a South Brazilian coastal lagoon. Ecotoxicol. Environ. Saf. 2008, 69, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Lewis, W.M.; Wurtsbaugh, W.A.; Paerl, H.W. Rationale for Control of Anthropogenic Nitrogen and Phosphorus to Reduce Eutrophication of Inland Waters. Environ. Sci. Technol. 2011, 45, 10300–10305. [Google Scholar] [CrossRef]
- Zeng, G.M.; Wang, P.; Huang, T. Extraction of Chlorophyll-a from Eutrophic Water by Repeated Freezing and Thawing-Extraction Method. Asian J. Chem. 2014, 26, 2289–2292. [Google Scholar] [CrossRef]
- Xie, J.; Hu, W.R.; Pei, H.Y. Detection of amount and activity of living algae in fresh water by dehydrogenase activity (DHA). Environ. Monit. Assess. 2008, 146, 473–478. [Google Scholar] [CrossRef]
- Ma, H.; Cui, F.Y.; Fan, Z.Q.; Liu, Z.Q.; Zhao, Z.W. Efficient control of Microcystis blooms by promoting biological filter-feeding in raw water. Ecol. Eng. 2012, 47, 71–75. [Google Scholar] [CrossRef]
- Manage, P.M.; Edwards, C.; Singh, B.K.; Lawton, L.A. Isolation and Identification of Novel Microcystin-Degrading Bacteria. Appl. Environ. Microbiol. 2009, 75, 6924–6928. [Google Scholar] [CrossRef] [Green Version]
- Matthijs, H.C.P.; Visser, P.M.; Reeze, B.; Meeuse, J.; Slot, P.C.; Wijn, G.; Talens, R.; Huisman, J. Selective suppression of harmful cyanobacteria in an entire lake with hydrogen peroxide. Water Res. 2012, 46, 1460–1472. [Google Scholar] [CrossRef]
- Paerl, H.W.; Xu, H.; McCarthy, M.J.; Zhu, G.W.; Qin, B.Q.; Li, Y.P.; Gardner, W.S. Controlling harmful cyanobacterial blooms in a hyper-eutrophic lake (Lake Taihu, China): The need for a dual nutrient (N & P) management strategy. Water Res. 2011, 45, 1973–1983. [Google Scholar]
- Park, J.H.; Yoshinaga, I.; Nishikawa, T.; Imai, I. Algicidal bacteria in particle-associated form and in free-living form during a diatom bloom in the Seto Inland Sea, Japan. Aquat. Microb. Ecol. 2010, 60, 151–161. [Google Scholar] [CrossRef] [Green Version]
- Park, S.C.; Lee, J.K.; Kim, S.W.; Park, Y. Selective Algicidal Action of Peptides against Harmful Algal Bloom Species. PLoS ONE 2011, 6, e26733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, H.F.; Chen, W.; Sun, L.W.; Jin, Y.X.; Liu, W.P.; Fu, Z.W. Inhibitory effects of paraquat on photosynthesis and the response to oxidative stress in Chlorella vulgaris. Ecotoxicology 2009, 18, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.H.; Fan, M.H. The Current State of Water Quality and Technology Development for Water Pollution Control in China. Crit. Rev. Environ. Sci. Technol. 2010, 40, 519–560. [Google Scholar] [CrossRef]
- Imamura, N.; Motoike, I.; Shimada, N. An efficient screening approach for anti-Microcystis compounds based on knowledge of aquatic microbial ecosystem. J. Antibiot. 2001, 54, 582–587. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.O.; Kato, J.; Takiguchi, N. Involvement of an extracellular protease in algicidal activity of the marine fungi Pseudoalteromonas sp strain A28. Appl. Environ. Microbiol. 2000, 66, 4334–4339. [Google Scholar] [CrossRef] [Green Version]
- Banin, E.; Khare, S.K.; Naider, F. Proline-rich peptide from the coral pathogen Vibrio shiloi that inhibits photosynthesis of zooxanthellae. Appl. Environ. Microbiol. 2001, 67, 1536–1541. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Content (g/L) | Soil Nutrient Solution, Content (g/L) | Ingredient |
|---|---|---|---|
| NaNO3 | 0.025 | H3BO3 | 0.286 |
| NaCl | 0.0025 | MnCl2·4H2O | 0.181 |
| K2HPO4·3H2O | 0.0075 | ZnSO4·7H2O | 0.0022 |
| MgSO4·7H2O | 0.0075 | CuSO4·5H2O | 0.0079 |
| CaCl2·2H2O | 0.0025 | Na2MoO4·2H2O | 0.0039 |
| KH2PO4 | 0.0175 | CoCl2·6H2O | 0.0020 |
| FeCl3·6H2O | 0.0005 | ||
| EDTA-Na | 0.0002 | ||
| Soil extract solution | 40 mL−L |
| Inlet Water (ug/L) | 145.58 (Low) | 151.67 (Medium) | 161.64 (High) |
|---|---|---|---|
| 1#Out of the water (ug/L) | 21.82 | 23.48 | 26.38 |
| 2#Out of the water (ug/L) | 71.15 | 73.34 | 78.56 |
| 3#Out of the water (ug/L) | 102.38 | 109.58 | 119.26 |
| 4#Comparison of water output (ug/L) | 149.53 | 154.67 | 165.79 |
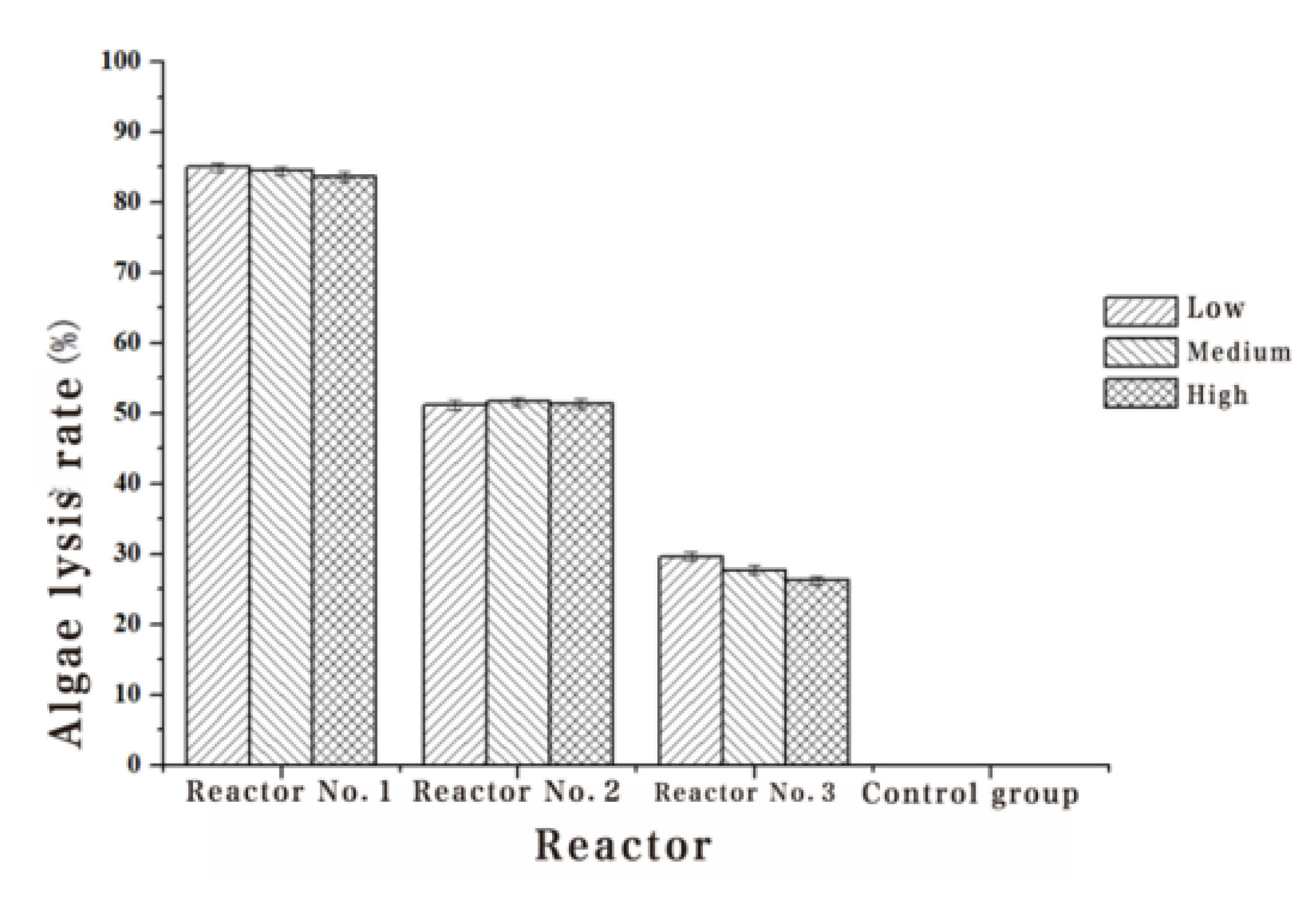
| 1#Algae lysis rate (%) | 85.01 | 84.52 | 83.68 |
| 2#Algae lysis rate (%) | 51.13 | 51.65 | 51.40 |
| 3#Algae lysis rate (%) | 29.67 | 27.75 | 26.22 |
| 4#Algae lysis rate (%) | - | - | - |
| Inlet Water (ug TPF/mL.h) | 0.83 | 0.87 | 0.89 |
|---|---|---|---|
| 1#Out of the water (ug TPF/mL.h) | 0.34 | 0.36 | 0.37 |
| 2#Out of the water (ug TPF/mL.h) | 0.47 | 0.50 | 0.49 |
| 3#Out of the water (ug TPF/mL.h) | 0.61 | 0.64 | 0.66 |
| 4#Comparison of water output (ug TPF/mL.h) | 0.86 | 0.90 | 0.95 |
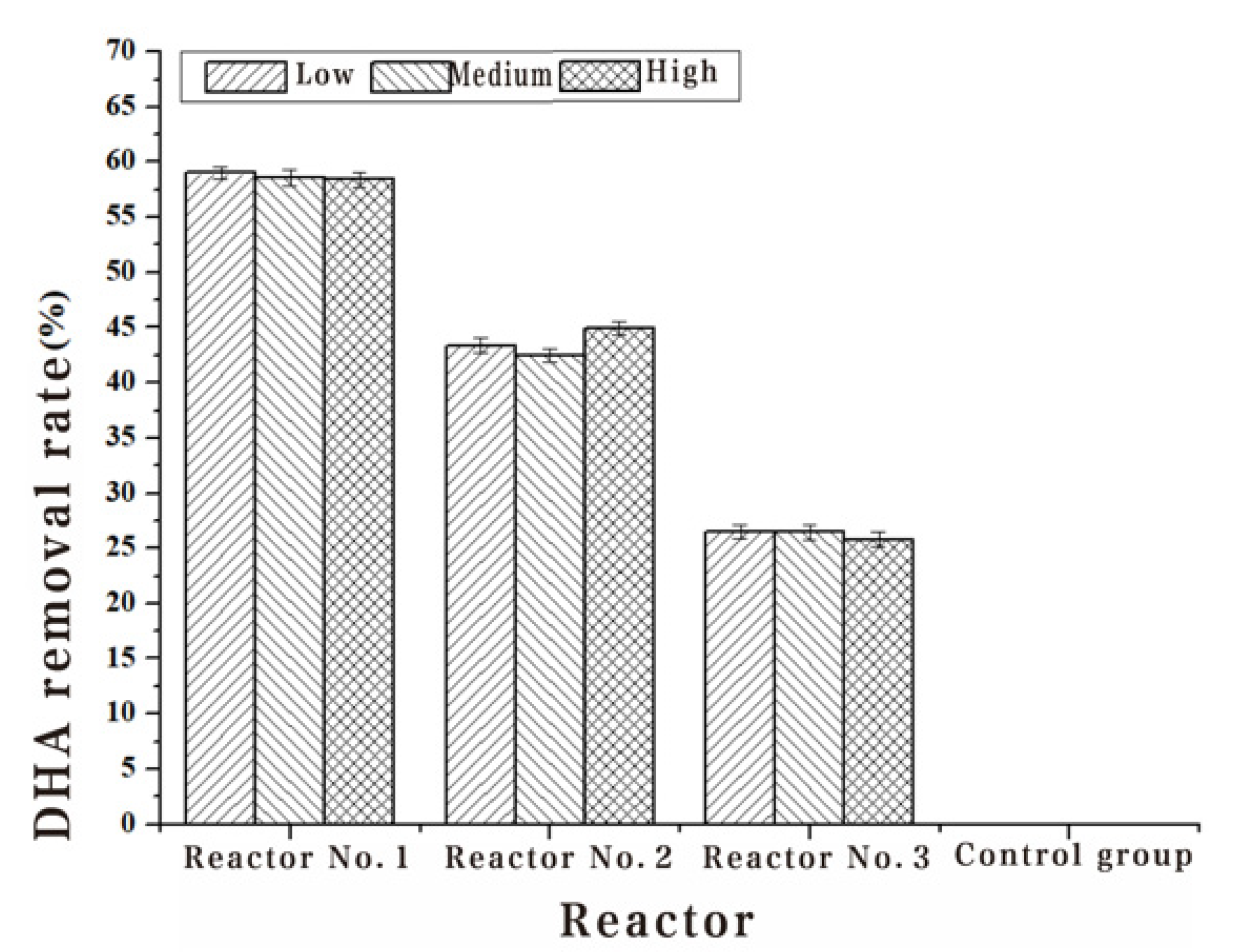
| 1#Removal rate (%) | 59.04 | 58.62 | 58.43 |
| 2#Removal rate (%) | 43.37 | 42.53 | 44.94 |
| 3#Removal rate (%) | 26.51 | 26.44 | 25.84 |
| 4#Removal rate (%) | - | - | - |
| Inlet Water (ug/mL) | 10.76 | 10.84 | 10.87 |
|---|---|---|---|
| 1#Out of the water (ug/mL) | 3.73 | 3.71 | 3.74 |
| 2#Out of the water (ug/mL) | 4.63 | 4.67 | 4.69 |
| 3#Out of the water (ug/mL) | 8.17 | 8.21 | 8.25 |
| 4#Comparison of water output (ug/mL) | 10.81 | 10.89 | 10.94 |
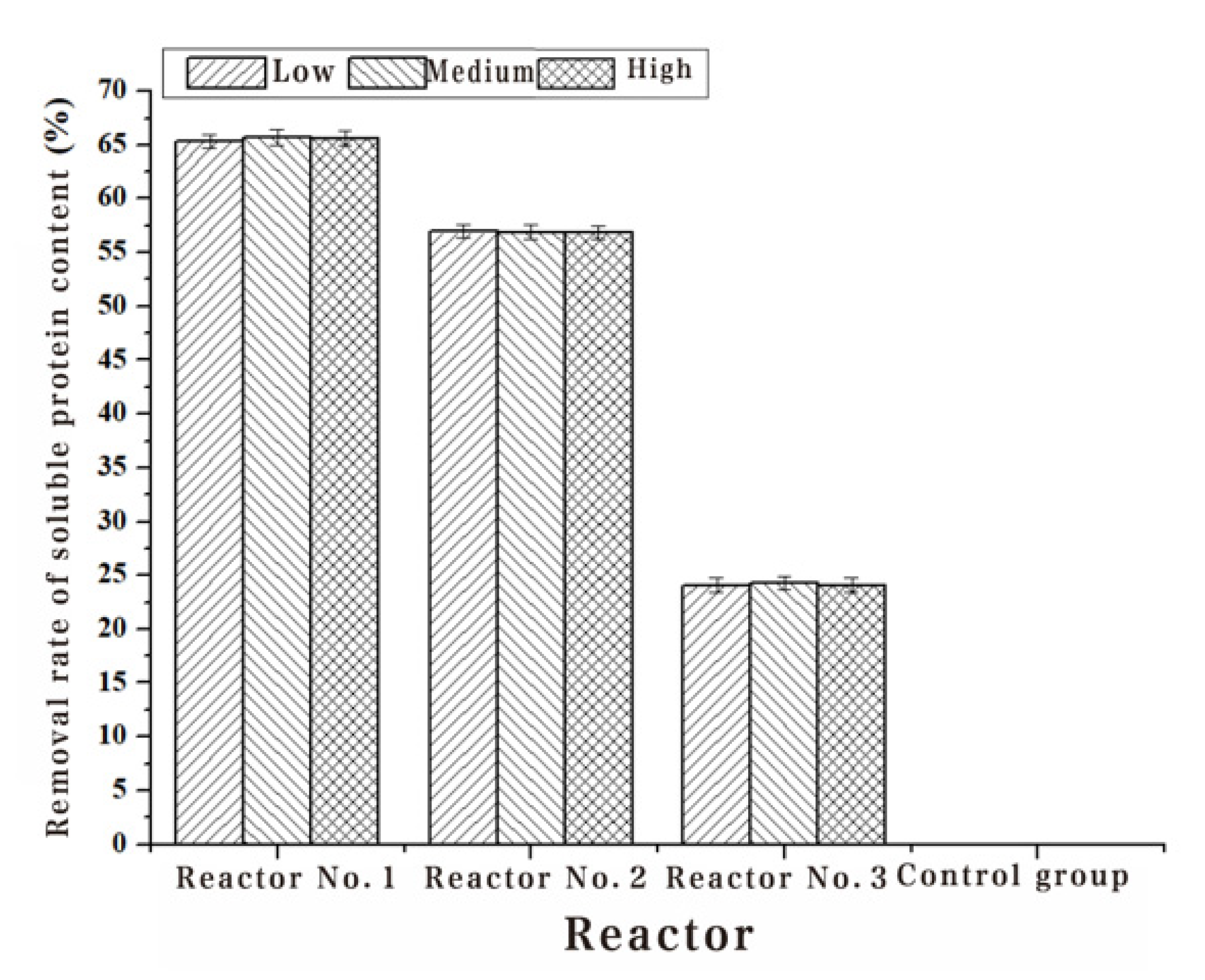
| 1#Removal rate (%) | 65.33 | 65.77 | 65.59 |
| 2#Removal rate (%) | 56.97 | 56.92 | 56.85 |
| 3#Removal rate (%) | 24.07 | 24.26 | 24.10 |
| 4#Removal rate (%) | - | - | - |
| Inlet Water (umol/L) | 0.031 | 0.031 | 0.030 |
|---|---|---|---|
| 1#Out of the water (umol/L) | 0.092 | 0.093 | 0.093 |
| 2#Out of the water (umol/L) | 0.067 | 0.065 | 0.067 |
| 3#Out of the water (umol/L) | 0.054 | 0.051 | 0.053 |
| 4#Out of the water (umol/L) | 0.031 | 0.031 | 0.030 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Zeng, G.; Wang, F.; Huang, X.; Li, Y.; Liang, D.; Zhang, M.; Sun, D. Study on the Algae Lysis Method of White Rot Fungi Algae Control System. Water 2022, 14, 903. https://doi.org/10.3390/w14060903
Wang J, Zeng G, Wang F, Huang X, Li Y, Liang D, Zhang M, Sun D. Study on the Algae Lysis Method of White Rot Fungi Algae Control System. Water. 2022; 14(6):903. https://doi.org/10.3390/w14060903
Chicago/Turabian StyleWang, Jun, Guoming Zeng, Fei Wang, Xin Huang, Yan Li, Dong Liang, Maolan Zhang, and Da Sun. 2022. "Study on the Algae Lysis Method of White Rot Fungi Algae Control System" Water 14, no. 6: 903. https://doi.org/10.3390/w14060903
APA StyleWang, J., Zeng, G., Wang, F., Huang, X., Li, Y., Liang, D., Zhang, M., & Sun, D. (2022). Study on the Algae Lysis Method of White Rot Fungi Algae Control System. Water, 14(6), 903. https://doi.org/10.3390/w14060903