Potential Submerged Macrophytes to Mitigate Eutrophication in a High-Elevation Tropical Shallow Lake—A Mesocosm Experiment in the Andes

 ,
, 
 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
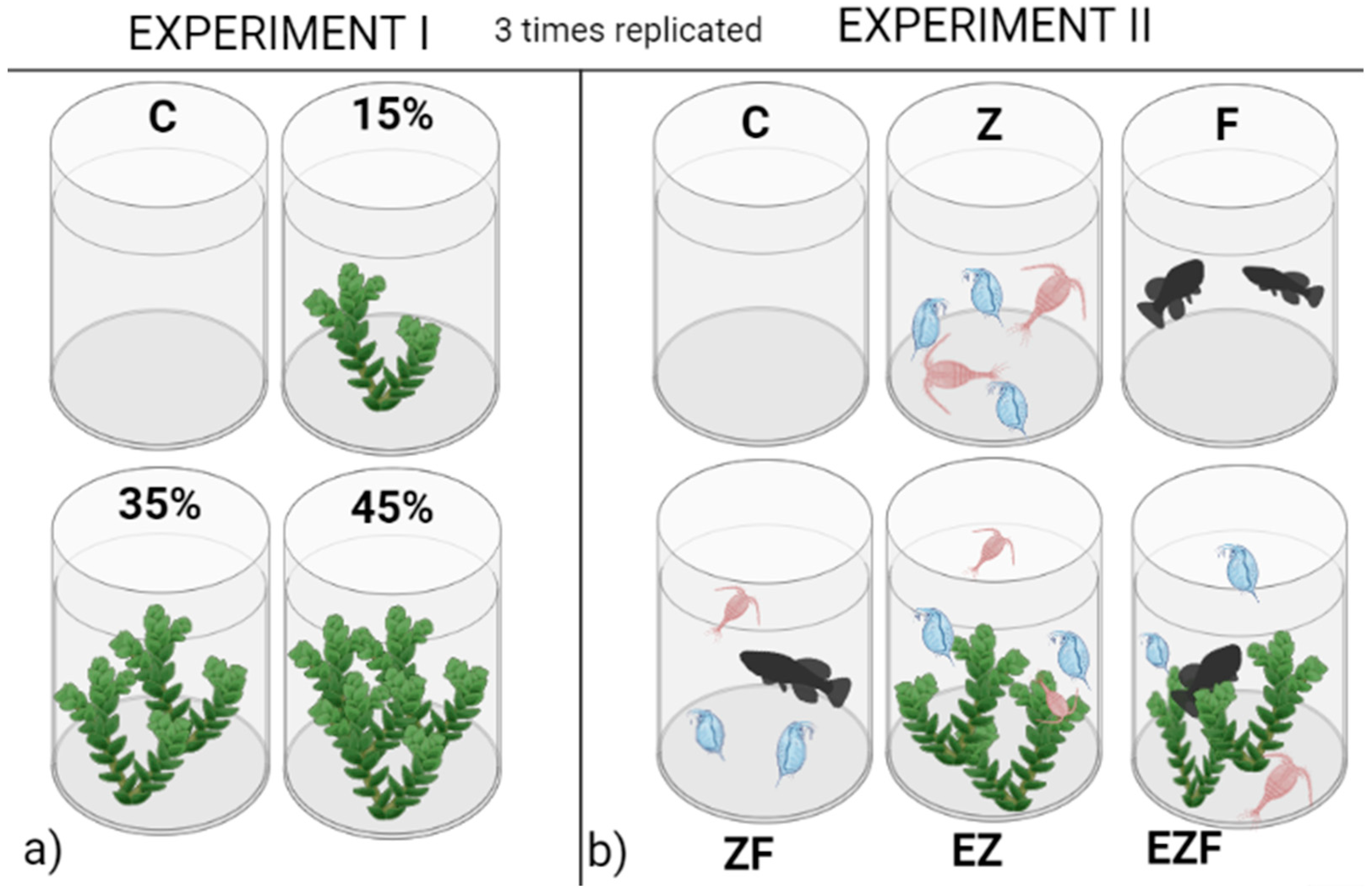
2.2. Experimental Set Up
2.3. Sample Analysis
2.4. Statistical Analyses
3. Results
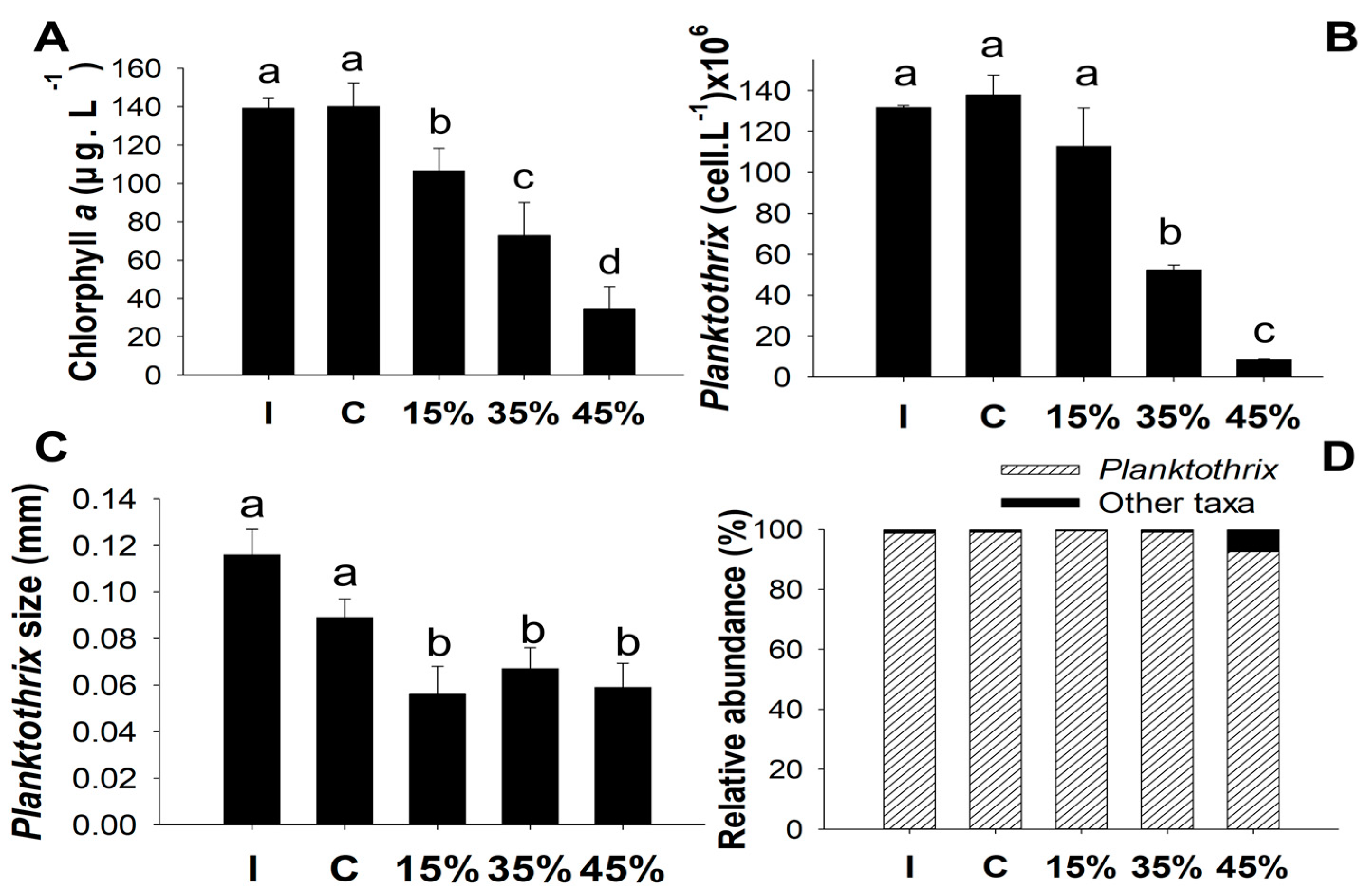
3.1. Experiment 1
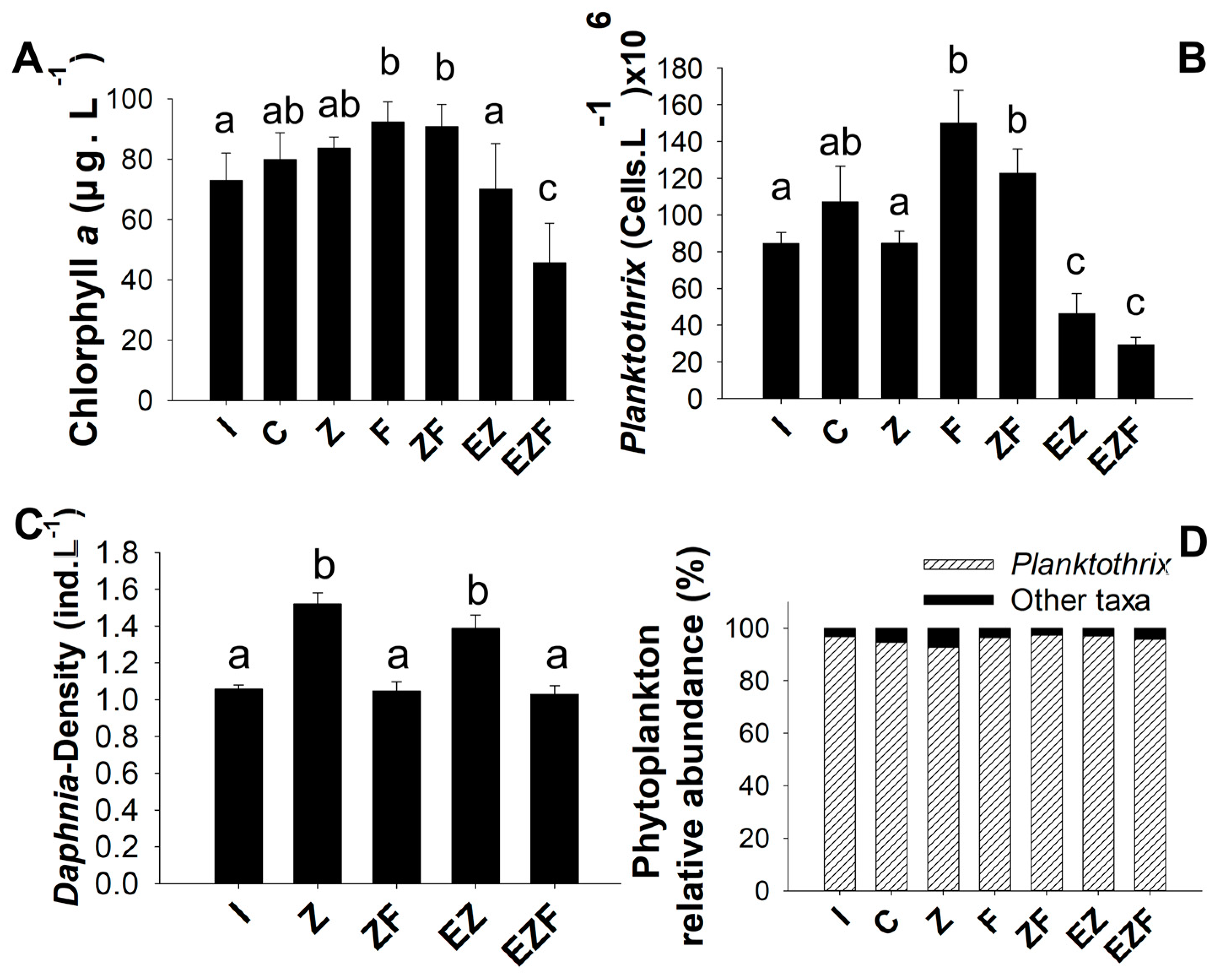
3.2. Experiment 2
4. Discussion
4.1. Phytoplankton Biomass and Total Nutrients with Different Concentrations of Macrophytes
4.2. Effects of Zooplankton on Phytoplankton
4.3. Effects of Fish on Zooplankton and Phytoplankton
4.4. Effects of Fish-Zooplankton and Macrophytes on Phytoplankton
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Experiment I | Experiment II | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CI | 15% | 35% | 45% | CII | F | Z | ZF | EZ | EZF | |
| Phytoplankton biomass | 0.66 ns | −23.6 * | −47.7 *** | −75.2 *** | 9.6 ns | 26.62 *** | 14.76 ns | 24.65 ** | −3.84 ns | −37.34 *** |
| Planktothrix density | 4.4 ns | −14.4 * | −60 *** | −93 *** | 26.7 ns | 77.5 *** | 0.15 ns | 45.18 * | −45.11 * | −65.21 *** |
| Planktothrix relative abundance | 99.12 | 99.53 | 99.10 | 92.55 | 94.57 | 97.04 | 92.68 | 97.28 | 69.93 | 95.76 |
| 30% | −10% | 34 | 9.1 | |||||||
| Daphnia | - | - | - | - | ||||||
| (Chla ug/L) | SD | Growth Rate (u) | SD | Planktothrix (Cell/L) × 106 | SD | Planktothrix Size | SD | TP (µg·L−1) | SD | TN (µg·L−1) | SD | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | 139.14 | 5.24 | 131.7 | 0.9 | 0.113 | 0.011 | 89.3 | 21 | 4866 | 750 | ||
| C | 140.06 | 12.28 | 0.0003 | 0.008 | 137.6 | 9.8 | 0.089 | 0.008 | 99 | 96.1 | 5000 | 1000 |
| 15% | 106.26 | 11.99 | −0.027 | 0.010 | 112.7 | 18.8 | 0.059 | 0.01 | 73.3 | 20.8 | 4066 | 1101 |
| 35% | 72.72 | 17.28 | −0.067 | 0.023 | 52.2 | 2.3 | 0.056 | 0.012 | 41.6 | 34.5 | 5666 | 577 |
| 45% | 34.5 | 11.5 | −0.144 | 0.032 | 8.4 | 0.2 | 0.067 | 0.009 | 52.3 | 15.3 | 3133 | 2759 |
| (Chla ug/L) | SD | Growth Rate (u) | SD | Planktothrix (Cell/L) × 106 | SD | TP (µg·L−1) | SD | TN (µg·L−1) | SD | Daphnia (ind L−1) | SD | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | 72.8 | 9.1 | 84.4 | 6 | 75 | 21.2 | 4250 | 353.55 | 1.164 | 0.048 | ||
| C | 79.8 | 8.9 | 0.006 | 0.008 | 107 | 19.4 | 75.6 | 46.6 | 5066 | 1006.64 | ||
| Z | 83.6 | 3.7 | 0.018 | 0.006 | 149.9 | 17.8 | 133.3 | 66.5 | 5233. | 251.66 | 1.52 | 0.060 |
| F | 92.2 | 6.7 | 0.009 | 0.003 | 84.5 | 6.6 | 116.6 | 47.2 | 5066 | 115.47 | ||
| ZF | 90.8 | 7.3 | 0.015 | 0.006 | 122.6 | 13.2 | 123.3 | 61.1 | 5666 | 577.35 | 1.04 | 0.051 |
| EZ | 70 | 15 | −0.004 | 0.014 | 46.3 | 10.7 | 93.3 | 15.2 | 5166 | 57.735 | 1.56 | 0.07 |
| EZF | 45.6 | 13 | −0.036 | 0.022 | 29.3 | 3.9 | 79.3 | 35.7 | 4800 | 692.82 | 1.27 | 0.02 |
References
- Smith, V.H.; Tilman, G.D.; Nekola, J.C. Eutrophication: Impacts of excess nutrient inputs on freshwater, marine, and terrestrial ecosystems. Environ. Pollut. 1999, 100, 179–196. [Google Scholar] [CrossRef] [PubMed]
- Hilt, S.; Brothers, S.; Jeppesen, E.; Veraart, A.J.; Kosten, S. Translating regime shifts in shallow lakes into changes in ecosystem functions and services. BioScience 2017, 67, 928–936. [Google Scholar] [CrossRef]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Jeppesen, E.; Jensen, J.P.; Jensen, C.; Faafeng, B.; Hessen, D.O.; Søndergaard, M.; Lauridsen, T.; Brettum, P.; Christoffersen, K. The impact of nutrient state and lake depth on top-down control in the pelagic zone of lakes: A study of 466 lakes from the temperate zone to the arctic. Ecosystems 2003, 313–325. [Google Scholar] [CrossRef]
- Hargeby, A.; Blindow, I.; Hansson, L.A. Shifts between clear and turbid states in a shallow lake: Multi-causal stress from climate, nutrients and biotic interactions. Arch. Fur. Hydrobiol. 2004, 161, 433–454. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Boyer, G.L. Health impacts from cyanobacteria harmful algae blooms: Implications for the North American Great Lakes. Harmful. Algae 2016, 54, 194–212. [Google Scholar] [CrossRef] [PubMed]
- de Magalhães, V.F.; Soares, R.M.; Azevedo, S.M. Microcystin contamination in fish from the Jacarepaguá Lagoon (Rio de Janeiro, Brazil): Ecological implication and human health risk. Toxicon 2001, 39, 1077–1085. [Google Scholar] [CrossRef]
- Barros, M.U.; Wilson, A.E.; Leitão, J.I.; Pereira, S.P.; Buley, R.P.; Fernandez-Figueroa, E.G.; Capelo-Neto, J. Environmental factors associated with toxic cyanobacterial blooms across 20 drinking water reservoirs in a semi-arid region of Brazil. Harmful. Algae 2019, 86, 128–137. [Google Scholar] [CrossRef]
- Coltorti, M.; Ollier, C.D. Geomorphic and tectonic evolution of the Ecuadorian Andes. Geomorphology 2000, 32, 1–19. [Google Scholar] [CrossRef]
- Andrade, G.I.; Franco, L.; Delgado, J. Socio-ecological barriers to adaptive management of lake Fuquene, Colombia. Int. J. Des. Nat. Ecodynamics 2012, 7, 251–260. [Google Scholar] [CrossRef]
- Velez, M.I.; Salgado, J.; Brenner, M.; Hooghiemstra, H.; Escobar, J.; Boom, A.; Bird, B.; Curtis, J.H.; Temoltzin-Loranca, Y.; Patino, L.F.; et al. Novel responses of diatoms in neotropical mountain lakes to indigenous and post-European occupation. Anthropocene 2021, 34, 100294. [Google Scholar] [CrossRef]
- Ayala, R.; Acosta, F.; Mooij, W.M.; Rejas, D.; Van Damme, P.A. Management of Laguna Alalay: A case study of lake restoration in Andean valleys in Bolivia. Aquat. Ecol. 2007, 41, 621–630. [Google Scholar] [CrossRef]
- Zolá, R.P.; Bengtsson, L. Long-term and extreme water level variations of the shallow Lake Poopó, Bolivia. Hydrol. Sci. J. 2006, 51, 98–114. [Google Scholar] [CrossRef] [Green Version]
- Mariano, M.; Huaman, P.; Mayta, E.; Montoya, H.; Chanco, M. Contaminación producida por piscicultura intensiva en lagunas andinas de Junín, Perú. Rev. Peru. De Biol. 2010, 17, 137–140. [Google Scholar] [CrossRef] [Green Version]
- Guevara, E.A.; Santander, T.; Espinosa, R.; Graham, C.H. Aquatic bird communities in Andean lakes of Ecuador are increasingly dissimilar over time. Ecol. Indic. 2021, 121, 107044. [Google Scholar] [CrossRef]
- Jacobsen, D.; Dangles, O. Ecology of High Altitude Waters; Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- Salomón, S.; Rivera-Rondón, C.A.; Zapata, Á.M. Floraciones de cianobacterias en Colombia: Estado del conocimiento y necesidades de investigación ante el cambio global. Rev. De La Acad. Colomb. De Cienc. Exactas Físicas Y Nat. 2020, 44, 376–391. [Google Scholar] [CrossRef]
- Jeppesen, E.; Meerhoff, M.; Jacobsen, B.A.; Hansen, R.S.; Søndergaard, M.; Jensen, J.P.; Lauridsen, T.; Mazzeo, N.; Branco, C.W.C. Restoration of shallow lakes by nutrient control and biomanipulation—The successful strategy varies with lake size and climate. Hydrobiologia 2007, 581, 269–285. [Google Scholar] [CrossRef]
- Søndergaard, M.; Jeppesen, E.; Lauridsen, T.L.; Skov, C.; Van Nes, E.H.; Roijackers, R.; Lammens, E.; Portielje, R.O.B. Lake restoration: Successes, failures and long-term effects. J. Appl. Ecol. 2007, 44, 1095–1105. [Google Scholar] [CrossRef]
- Yu, J.; Liu, Z.; Li, K.; Chen, F.; Guan, B.; Hu, Y.; Zhong, P.; Tang, Y.; Zhao, X.; He, H.; et al. Restoration of shallow lakes in subtropical and tropical China: Response of nutrients and water clarity to biomanipulation by fish removal and submerged plant transplantation. Water 2016, 8, 438. [Google Scholar] [CrossRef] [Green Version]
- Søndergaard, M.; Moss, B. Impact of submerged macrophytes on phytoplankton in shallow freshwater lakes. In The Structuring Role of Submerged Macrophytes in Lakes; Springer: New York, NY, USA, 1998; pp. 115–132. [Google Scholar]
- Van Donk, E.; van de Bund, W.J. Impact of submerged macrophytes including charophytes on phyto-and zooplankton communities: Allelopathy versus other mechanisms. Aquat. Bot. 2002, 72, 261–274. [Google Scholar] [CrossRef]
- Jeppesen, E.; Søndergaard, M.; Søndergaard, M.; Christoffersen, K. (Eds.) The Structuring Role of Submerged Macrophytes in Lakes; Springer Science & Business Media: New York, NY, USA, 2012; Volume 131. [Google Scholar]
- Dong, J.; Yang, K.; Li, S.; Li, G.; Song, L. Submerged vegetation removal promotes shift of dominant phytoplankton functional groups in a eutrophic lake. J. Environ. Sci. 2014, 26, 1699–1707. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, X.; Liu, J.; Hong, Y.; He, Q.; Yu, D.; Liu, C.; Dingshanbayi, H. Greater performance of exotic Elodea nuttallii in response to water level may make it a better invader than exotic Egeria densa during winter and spring. Front. Plant Sci. 2019, 10, 144. [Google Scholar] [CrossRef] [PubMed]
- Horppila, J.; Nurminen, L. Effects of submerged macrophytes on sediment resuspension and internal phosphorus loading in Lake Hiidenvesi (southern Finland). Water Res. 2003, 37, 4468–4474. [Google Scholar] [CrossRef] [PubMed]
- Lazzaro, X. Do the trophic cascade hypothesis and classical biomanipulation approaches apply to tropical lakes and reservoirs? Int. Ver. Für Theor. Und Angew. Limnol. Verh. 1997, 26, 719–730. [Google Scholar] [CrossRef]
- Meerhoff, M.; Iglesias, C.; Teixeira de Mello, F.T.; Clemente, J.M.; Jensen, E.; Lauridsen, T.L.; Jeppesen, E. Effects of habitat complexity on community structure and predator avoidance behaviour of littoral zooplankton in temperate versus subtropical shallow lakes. Freshw. Biol. 2007, 52, 1009–1021. [Google Scholar] [CrossRef]
- Iglesias, C.; Goyenola, G.; Mazzeo, N.; Meerhoff, M.; Rodo, E.; Jeppesen, E. Horizontal dynamics of zooplankton in subtropical Lake Bianca (Uruguay) hosting multiple Zooplankton predators and aquatic plant refuges. In Shallow Lakes in a Changing World; Springer: Dordrecht, The Nertherlands, 2007; pp. 179–189. [Google Scholar]
- Teixeira de Mello, F.; de Oliveira, V.A.; Loverde-Oliveira, S.M.; Huszar, V.L.M.; Barquín, J.; Iglesias, C.; Silva, T.S.F.; Duque-Estrada, C.H.; Silió-Calzada, A.; Mazzeo, N. The structuring role of free-floating plants on the fish community in a tropical shallow lake: An experimental approach with natural and artificial plants. Hydrobiologia 2016, 778, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Quirós, R. Fish effects on trophic relationships in the pelagic zone of lakes. Hydrobiologia 1997, 361, 101–111. [Google Scholar] [CrossRef]
- Sinistro, R. Top-down and bottom-up regulation of planktonic communities in a warm temperate wetland. J. Plankton Res. 2010, 32, 209–220. [Google Scholar] [CrossRef]
- Vanderstukken, M.; Mazzeo, N.; Van Colen, W.; Declerck, S.A.; Muylaert, K. Biological control of phytoplankton by the subtropical submerged macrophytes Egeria densa and Potamogeton illinoensis: A mesocosm study. Freshw. Biol. 2011, 56, 1837–1849. [Google Scholar] [CrossRef]
- Yu, J.; Liu, Z.; He, H.; Zhen, W.; Guan, B.; Chen, F.; Li, K.; Zhong, P.; Teixeira de Mello, F.; Jeppesen, E. Submerged macrophytes facilitate dominance of omnivorous fish in a subtropical shallow lake: Implications for lake restoration. Hydrobiologia 2016, 775, 97–107. [Google Scholar] [CrossRef]
- Amorim, C.A.; Moura, A.N. Effects of the manipulation of submerged macrophytes, large zooplankton, and nutrients on a cyanobacterial bloom: A mesocosm study in a tropical shallow reservoir. Environ. Pollut. 2020, 265, 114997. [Google Scholar] [CrossRef] [PubMed]
- Gunkel, G.; Casallas, J. Limnology of an equatorial high mountain lake—Lago San Pablo, Ecuador: The significance of deep diurnal mixing for lake productivity. Limnologica 2002, 32, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Van Colen, W.; Portilla, K.; Oña, T.; Wyseure, G.; Goethals, P.; Velarde, E.; Muylaert, K. Limnology of the neotropical high elevation shallow lake Yahuarcocha (Ecuador) and challenges for managing eutrophication using biomanipulation. Limnologica 2017, 67, 37–44. [Google Scholar] [CrossRef]
- Scheffer, M.; Hosper, S.H.; Meijer, M.L.; Moss, B.; Jeppesen, E. Alternative equilibria in shallow lakes. Trends Ecol. Evol. 1993, 8, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Setubal, R.B.; Riccardi, N. Long-term effects of fish biomanipulation and macrophyte management on zooplankton functional diversity and production in a temperate shallow lake. Limnology 2020, 21, 305–317. [Google Scholar] [CrossRef]
- Cooke, G.D.; Welch, E.B.; Peterson, S.; Nichols, S.A. Restoration and Management of Lakes and Reservoirs; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Hilt, S.; Gross, E.M.; Hupfer, M.; Morscheid, H.; Mählmann, J.; Melzer, A.; Poltz, J.; Sandrock, S.; Scharf, E.M.; Schneider, S.; et al. Restoration of submerged vegetation in shallow eutrophic lakes–A guideline and state of the art in Germany. Limnologica 2006, 36, 155–171. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, S.R.; Lodge, D.M. Effects of submersed macrophytes on ecosystem processes. Aquat. Bot. 1986, 26, 341–370. [Google Scholar] [CrossRef]
- Liu, Z.; Hu, J.; Zhong, P.; Zhang, X.; Ning, J.; Larsen, S.E.; Chen, D.; Gao, Y.; He, H.; Jeppesen, E. Successful restoration of a tropical shallow eutrophic lake: Strong bottom-up but weak top-down effects recorded. Water Res. 2018, 146, 88–97. [Google Scholar] [CrossRef]
- Hussner, A. Alien aquatic plant species in European countries. Weed Res. 2012, 52, 297–306. [Google Scholar] [CrossRef]
- Espinosa-Rodríguez, C.A.; Sarma, S.S.S.; Nandini, S. Effect of the allelochemicals from the macrophyte Egeria densa on the competitive interactions of pelagic and littoral cladocerans. Chem. Ecol. 2017, 33, 247–256. [Google Scholar] [CrossRef]
- Wu, Z.B.; Gao, Y.N.; Wang, J.; Liu, B.Y.; Zhou, Q.H.; Zhang, Y.Y. Allelopathic effects of phenolic compounds present in submerged macrophytes on Microcystis aeruginosa. Allelopath. J. 2009, 23, 403–410. [Google Scholar]
- Patton, C.J.; Kryskalla, J.R. Methods of Analysis by the US Geological Survey National Water Quality Laboratory: Evaluation of Alkaline Persulfate Digestion as an Alternative to Kjeldahl Digestion for Determination of Total and DISSOLVED Nitrogen and Phosphorus in Water; No. 4174; US Department of the Interior, US Geological Survey: Denver, CO, USA, 2003; Volume 3.
- Wang, H.; Zhong, G.; Yan, H.; Liu, H.; Wang, Y.; Zhang, C. Growth control of cyanobacteria by three submerged macrophytes. Environ. Eng. Sci. 2012, 29, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Nakai, S.; Hosomi, M.; Okada, M.; Murakami, A. Control of algal growth by macrophytes and macrophyte-extracted bioactive compounds. Water Sci. Technol. 1996, 34, 227–235. [Google Scholar] [CrossRef]
- Erhard, D.; Gross, E.M. Allelopathic activity of Elodea canadensis and Elodea nuttallii against epiphytes and phytoplankton. Aquat. Bot. 2006, 85, 203–211. [Google Scholar] [CrossRef] [Green Version]
- Mazzeo, N.; Rodríguez-Gallego, L.; Kruk, C.; Meerhoff, M.; Gorga, J.; Lacerot, G.; Quintans, F.; Loureiro, M.; Larrea, D.; García-Rodríguez, F. Effects of Egeria densa Planch. beds on a shallow lake without piscivorous fish. Hydrobiologia 2003, 506, 591–602. [Google Scholar] [CrossRef]
- Mony, C.; Koschnick, T.J.; Haller, W.T.; Muller, S. Competition between two invasive Hydrocharitaceae (Hydrilla verticillata (Lf)(Royle) and Egeria densa (Planch)) as influenced by sediment fertility and season. Aquat. Bot. 2007, 86, 236–242. [Google Scholar] [CrossRef]
- Mohamed, Z.A. Macrophytes-cyanobacteria allelopathic interactions and their implications for water resources management—A review. Limnologica 2017, 63, 122–132. [Google Scholar] [CrossRef]
- Senavirathna, M.D.H.J.; Muhetaer, G.; Atapaththu, K.S.S.; Fujino, T. Egeria densa allelopathy on microcystis aeruginosa under different light intensities and preliminary insight into inter-parameter relationships. Water Air Soil Pollut. 2021, 232, 1–13. [Google Scholar] [CrossRef]
- Hu, H.; Hong, Y. Algal-bloom control by allelopathy of aquatic macrophytes—A review. Front. Environ. Sci. Eng. China 2008, 2, 421–438. [Google Scholar] [CrossRef]
- Vergalli, J.; Fayolle, S.; Combes, A.; Franquet, E.; Comte, K. Persistence of microcystin production by Planktothrix agardhii (Cyanobacteria) exposed to different salinities. Phycologia 2020, 59, 24–34. [Google Scholar] [CrossRef]
- Mulderij, G.; Van Nes, E.H.; Van Donk, E. Macrophyte–phytoplankton interactions: The relative importance of allelopathy versus other factors. Ecol. Model. 2007, 204, 85–92. [Google Scholar] [CrossRef]
- Brönmark, C.; Weisner, S.E. Indirect effects of fish community structure on submerged vegetation in shallow, eutrophic lakes: An alternative mechanism. In The Dynamics and Use of Lacustrine Ecosystems; Springer: Dordrecht, The Nertherlands, 1992; pp. 293–301. [Google Scholar]
- Lampert, W. Laboratory studies on zooplankton-cyanobacteria interactions. N. Z. J. Mar. Freshw. Res. 1987, 21, 483–490. [Google Scholar] [CrossRef]
- Ferrão-Filho, A.S.; Pereira, U.J.; Vilar, M.C.; de Magalhães, L.; Marinho, M.M. Can small-bodied Daphnia control Raphidiopsis raciborskii in eutrophic tropical lakes? A mesocosm experiment. Environ. Sci. Pollut. Res. 2020, 27, 35459–35473. [Google Scholar] [CrossRef] [PubMed]
- Dussault, G.V.; Kramer, D.L. Food and feeding behavior of the guppy, Poecilia reticulata (Pisces: Poeciliidae). Can. J. Zool. 1981, 59, 684–701. [Google Scholar] [CrossRef]
- Canfield, D.E., Jr.; Shireman, J.V.; Colle, D.E.; Haller, W.T.; Watkins II, C.E.; Maceina, M.J. Prediction of chlorophyll a concentrations in Florida lakes: Importance of aquatic macrophytes. Can. J. Fish. Aquat. Sci. 1984, 41, 497–501. [Google Scholar] [CrossRef]
- Pacheco, J.P.; Aznarez, C.; Meerhoff, M.; Liu, Y.; Li, W.; Baattrup-Pedersen, A.; Yu, C.; Jeppesen, E. Small-sized omnivorous fish induce stronger effects on food webs than warming and eutrophication in experimental shallow lakes. Sci. Total Environ. 2021, 797, 148998. [Google Scholar] [CrossRef] [PubMed]
- Teixeira de Mello, F.; Meerhoff, M.; Pekcan-Hekim, Z.; Jeppesen, E. Substantial differences in littoral fish community structure and dynamics in subtropical and temperate shallow lakes. Freshw. Biol. 2009, 54, 1202–1215. [Google Scholar] [CrossRef]
- Burks, R.L.; Lodge, D.M.; Jeppesen, E.; Lauridsen, T.L. Diel horizontal migration of zooplankton: Costs and benefits of inhabiting the littoral. Freshw. Biol. 2002, 47, 343–365. [Google Scholar] [CrossRef]
- Petry, A.C.; Guimarães, T.F.R.; Vasconcellos, F.M.; Hartz, S.M.; Becker, F.G.; Rosa, R.S.; Goyenola, G.; Caramaschi, E.P.; de Astarloa, J.M.D.; Sarmento-Soares, L.M.; et al. Fish composition and species richness in eastern South American coastal lagoons: Additional support for the freshwater ecoregions of the world. J. Fish Biol. 2016, 89, 280–314. [Google Scholar] [CrossRef]
- Meerhoff, M.; Mazzeo, N.; Moss, B.; Rodríguez-Gallego, L. The structuring role of free-floating versus submerged plants in a subtropical shallow lake. Aquat. Ecol. 2003, 37, 377–391. [Google Scholar] [CrossRef]
- Seghers, B.H. Schooling behavior in the guppy (Poecilia reticulata): An evolutionary response to predation. Evolution 1974, 28, 486–489. [Google Scholar] [PubMed]
- Araújo, F.G.; Peixoto, M.G.; Pinto, B.C.T.; Teixeira, T.P. Distribution of guppies Poecilia reticulata (Peters, 1860) and Phalloceros caudimaculatus (Hensel, 1868) along a polluted stretch of the Paraíba do Sul River, Brazil. Braz. J. Biol. 2009, 69, 41–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilt, S.; Elisabeth Gross, M. Can allelopathically active submerged macrophytes stabilize clear-water states in shallow lakes? Basic Appl. Ecol. 2008, 9, 422–432. [Google Scholar] [CrossRef]



| Treatments | Variables | ||||
|---|---|---|---|---|---|
| Phytoplankton Biomass | Planktothrix Density | Planktothrix Size | Nutrients (NP) | Daphnia Density | |
| C | + | + | + | + | − |
| 15% | + | + | + | + | − |
| 35% | + | + | + | + | − |
| 45% | + | + | + | + | − |
| C | + | + | − | + | + |
| Z | + | + | − | + | + |
| F | + | + | − | + | + |
| ZF | + | + | − | + | + |
| EZ | + | + | − | + | + |
| EZF | + | + | − | + | + |
| | | Experiment I | Experiment II | ||||
|---|---|---|---|---|---|---|
| df | F | P | df | F | P | |
| Chlorophyll a | 4 | 106.6 | <0.001 | 6 | 23.67 | <0.001 |
| Planktothrix density | 4 | 47.94 | <0.001 | 6 | 34.15 | <0.001 |
| Planktothrix size | 4 | 18.07 | <0.001 | - | - | - |
| TN | 4 | 1.32 | 0.333 | 6 | 1.5 | 0.25 |
| TP | 4 | 0.75 | 0.575 | 6 | 0.77 | 0.6 |
| Daphnia density | - | - | - | 4 | 45.91 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Portilla, K.; Velarde, E.; Decaestecker, E.; Teixeira de Mello, F.; Muylaert, K. Potential Submerged Macrophytes to Mitigate Eutrophication in a High-Elevation Tropical Shallow Lake—A Mesocosm Experiment in the Andes. Water 2023, 15, 75. https://doi.org/10.3390/w15010075
Portilla K, Velarde E, Decaestecker E, Teixeira de Mello F, Muylaert K. Potential Submerged Macrophytes to Mitigate Eutrophication in a High-Elevation Tropical Shallow Lake—A Mesocosm Experiment in the Andes. Water. 2023; 15(1):75. https://doi.org/10.3390/w15010075
Chicago/Turabian StylePortilla, Karen, Elizabeth Velarde, Ellen Decaestecker, Franco Teixeira de Mello, and Koenraad Muylaert. 2023. "Potential Submerged Macrophytes to Mitigate Eutrophication in a High-Elevation Tropical Shallow Lake—A Mesocosm Experiment in the Andes" Water 15, no. 1: 75. https://doi.org/10.3390/w15010075
APA StylePortilla, K., Velarde, E., Decaestecker, E., Teixeira de Mello, F., & Muylaert, K. (2023). Potential Submerged Macrophytes to Mitigate Eutrophication in a High-Elevation Tropical Shallow Lake—A Mesocosm Experiment in the Andes. Water, 15(1), 75. https://doi.org/10.3390/w15010075