

Possible Origin and Distribution of an Invasive Diatom Species, Skeletonema potamos, in Yangtze River Basin (China)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
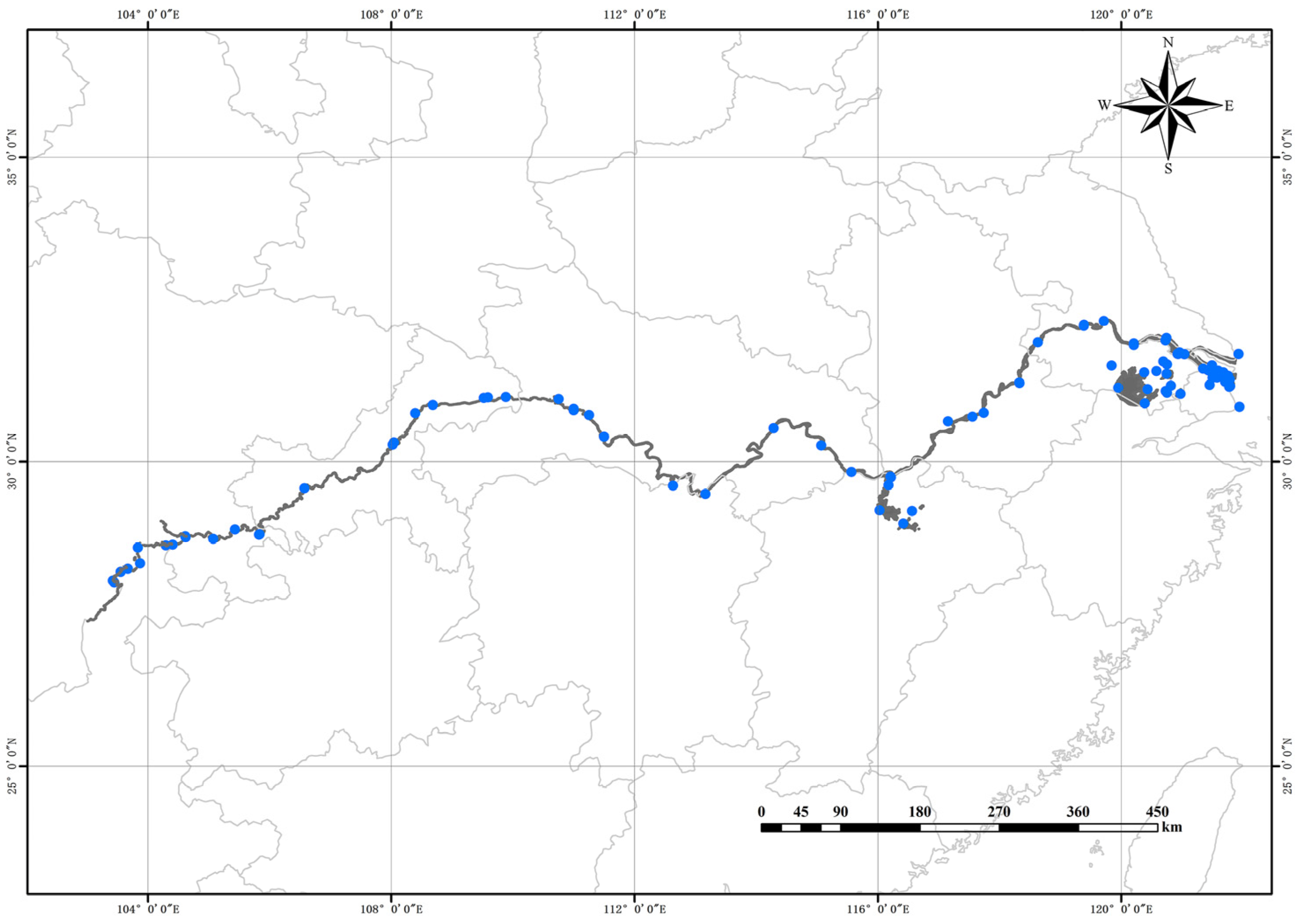
2.1. Sample Examination and Analysis
2.2. Sample Isolation and Cultivation
2.3. DNA Extraction, PCR Amplification, and Sequencing
2.4. Statistical Analysis
3. Results
3.1. Morphological Observations of S. potamos
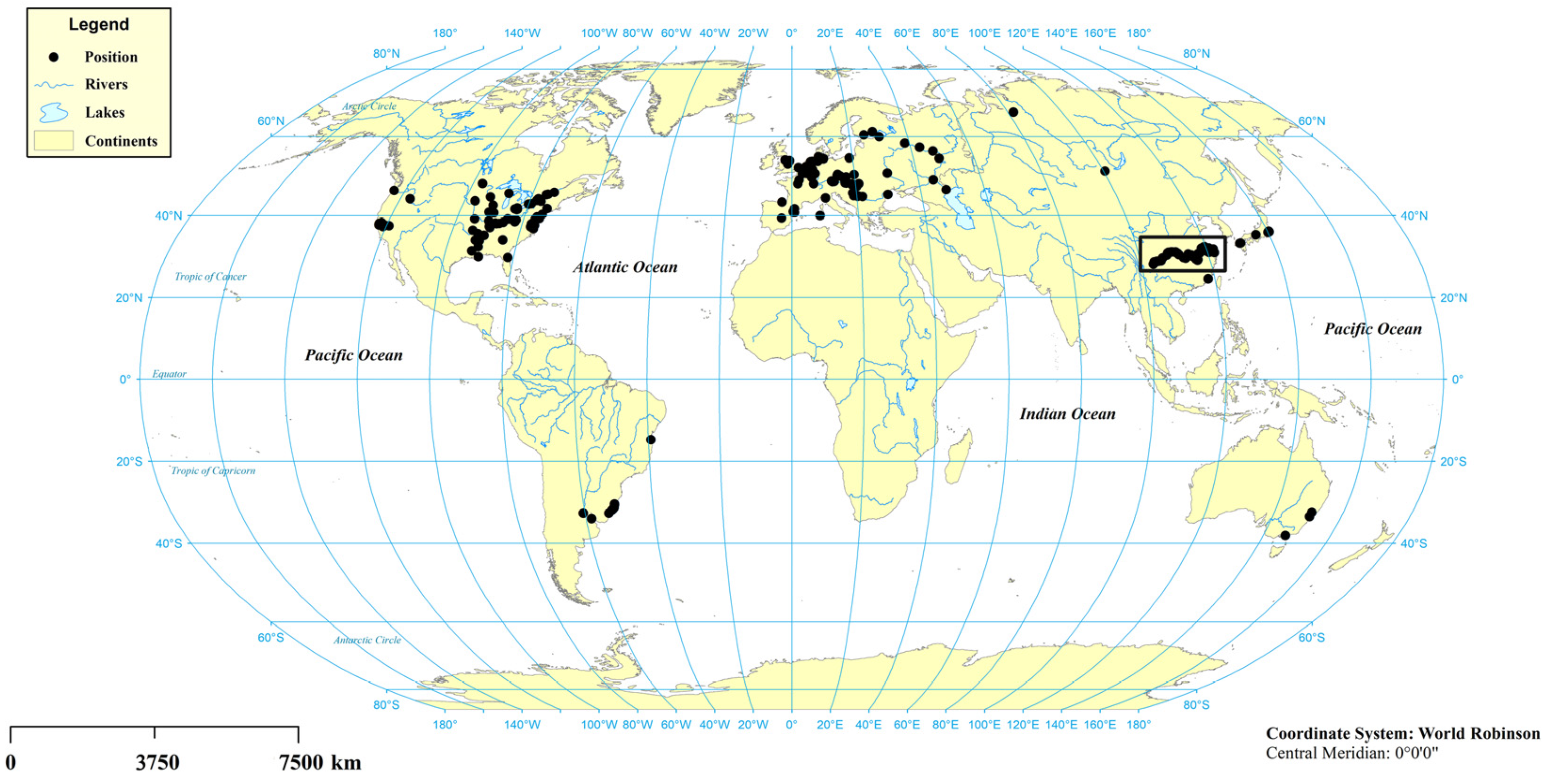
3.2. World Distribution of S. potamos
3.3. Genetic Diversity
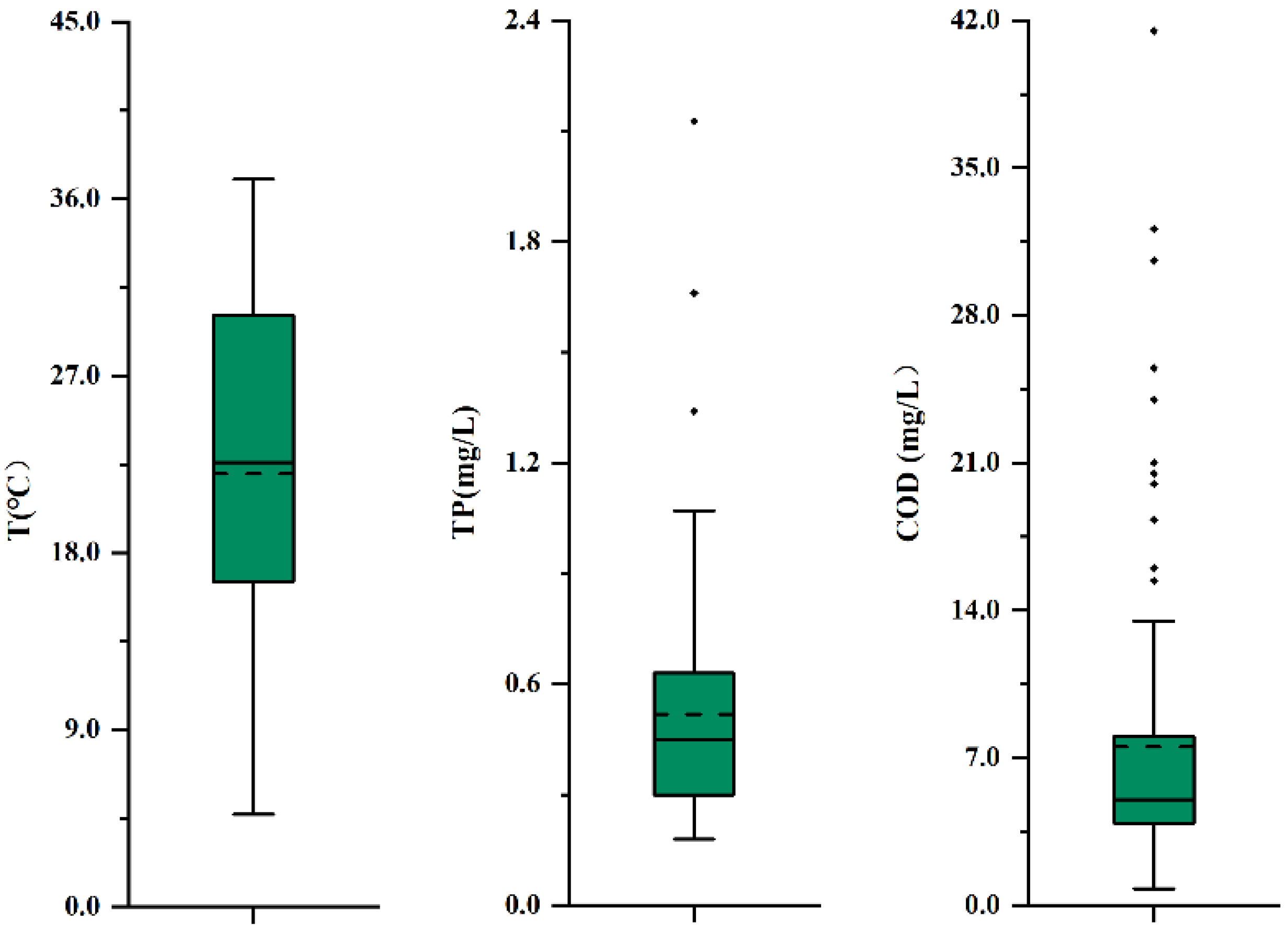
3.4. Ecology Analysis
4. Discussion
4.1. Morphological Comparison
4.2. Possible Origin of S. potamos in China
4.3. The Diffusion Pattern and Dominant Mechanism of S. potamos in China
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Duleba, M.; Ector, L.; Horváth, Z.; Kiss, K.T.; Molnar, L.; Pohner, Z.; Szilágyi, Z.; Tóth, B.; Vad, C.F.; Várbíró, G.; et al. Biogeography and phylogenetic position of a warm-stenotherm centric diatom, Skeletonema potamos (C.I. Weber) Hasle and its long-term dynamics in the River Danube. Protist 2014, 1655, 715–729. [Google Scholar] [CrossRef]
- Cheng, J.F.; Gao, Y.H.; Liang, J.R.; Chen, C.P. Research progress on species and gene diversity of Skeletonema. Prog. Nat. Sci. 2007, 17, 586–594. (In Chinese) [Google Scholar]
- Kiss, K.T.; Klee, R.; Ector, L.; Ács, É. Centric diatoms of large rivers and tributaries in Hungary: Morphology and biogeographic distribution. Acta Bot. Croat. 2012, 71, 311–363. [Google Scholar] [CrossRef] [Green Version]
- Poulin, M. A Multidisciplinary, Community-Based Study of the Environmental Health of the Rideau River: Final Report; Canadian Museum of Nature: Montréal, ON, Canada; Ottawa, ON, Canada, 2001; pp. 1–41. [Google Scholar]
- Hustedt, F. Die Kieselalgen Deutschlands, Österreichs und der Schweiz unter Berücksichtigung der Übrigen Länder Europas Sowie der Angrenzenden Meeresgebiete; Akademische Verlagsgesellschaft m.b.h. Leipzig: Leipzig, Germany, 1928. [Google Scholar]
- Hasle, G.R.; Evensen, D.L. Brackish water and freshwater species of the diatom genus Skeletonema. II. Skeletonema potamos comb. nov. J. Phycol. 1976, 12, 73–82. [Google Scholar] [CrossRef]
- Weber, C.I. A new freshwater centric diatom Microsiphona potamos gen. et sp. nov. J. Phycol. 1970, 6, 149–153. [Google Scholar] [CrossRef]
- Heudre, D.; Wetzel, C.E.; Vijver, B.V.D.; Moreau, L.; Ector, L. Brackish diatom species (Bacillariophyta) from rivers of Rhin-Meuse basin in France. Bot. Lett. 2020, 168, 56–84. [Google Scholar] [CrossRef]
- Torgan, L.C.; Becker, V.; Santos, C.B.D. Skeletonema potamos (Bacillariophyta) in Patos Lagoon, southern Brazil: Taxonomy and distribution. Rev. Peru. Biol. 2011, 16, 93–96. [Google Scholar]
- Cheng, Z.D.; Liu, S.C. A taxonomy study of Skeletonema Greville from Xiamen Harbour, China. J. Xiamen Univ. (Nat. Sci.) 1992, 31, 295–297. (In Chinese) [Google Scholar]
- Lin, G.; Yang, Q. Species diversity and the distribution of micro-phytoplankton in the Taiwan Strait. Biodivers. Sci. 2007, 15, 31–45. (In Chinese) [Google Scholar]
- Zhang, L.J.; Yang, J.H.; Zhang, Y.; Shi, J.Z.; Yu, H.X.; Zhang, X.W. eDNA biomonitoring revealed the ecological effects of water diversion projects between Yangtze River and Tai Lake. Water Res. 2022, 210, 117994. [Google Scholar] [CrossRef]
- Zhao, K.; Cao, Y.; Pang, W.T.; Wang, L.Z.; Song, K.; You, Q.M.; Wang, Q.X. Long-term plankton community dynamics and influencing factors in a man-made shallow lake, Lake Dishui, China. Aquat. Sci. 2021, 83, 1–14. [Google Scholar] [CrossRef]
- Hu, H.J.; Wei, Y.X. The Freshwater Algae of China Systematics, Taxonomy and Ecology; Sciences Press: Beijing, China, 2006. (In Chinese) [Google Scholar]
- Zhao, K.; Wang, L.Z.; You, Q.M.; Pan, Y.D.; Liu, T.T.; Zhou, Y.D.; Zhang, J.Y.; Pang, W.T.; Wang, Q.X. Influence of Cyanobacterial blooms and environmental variation on zooplankton and eukaryotic phytoplankton in a Large, Shallow, Eutrophic Lake in China. Sci. Total Environ. 2021, 773, 145421. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Yu, P.; You, Q.M.; Li, G.S.; Wang, Q.X. Morphological and phylogenetic analysis of a new Melosira species and revision of freshwater Melosira in China. J. Oceanol. Limnol. 2021, 40, 712–728. [Google Scholar] [CrossRef]
- Luo, F.; You, Q.M.; Wang, Q.X. A new species of Genkalia (Bacillariophyta) from mountain lakes within the Sichuan Province of China. Phytotaxa 2018, 372, 236–242. [Google Scholar] [CrossRef]
- Yu, P.; You, Q.M.; Kociolek, J.P.; Lowe, R.L.; Wang, Q.X. Nupela major sp. nov., a new diatom species from Maolan Nature Reserve, central-south of China. Phytotaxa 2017, 311, 245–254. [Google Scholar] [CrossRef]
- Jiang, X.D.; Chen, X.; Pang, W.T.; Wang, Q.X. Phylogeny of Trachelomonas and Strombomonas (Euglenaceae) based on morphological and molecular data. Diversity 2022, 14, 623. [Google Scholar] [CrossRef]
- Richlen, M.L.; Barber, P.H. A technique for the rapid extraction of microalgal DNA from single live and preserved cells. Mol. Ecol. Notes 2005, 5, 688–691. [Google Scholar] [CrossRef]
- Orsini, L.; Sarno, D.; Procaccini, D.; Poletti, R.; Dahlmann, J.; Montresor, M. Toxic Pseudo-nitzschia multistriata (Bacillariophyceae) from the Gulf of Naples: Morphology, toxin analysis and phylogenetic relationships with other Pseudo-nitzschia species. Eur. J. Phycol. 2002, 37, 247–257. [Google Scholar] [CrossRef]
- Theriot, E.C.; Ruck, E.C.; Ashworth, M.; Nakov, T.; Jansen, R.K. Status of the pursuit of the diatom phylogeny: Are traditional views and new molecular paradigms really that different? Cell. Orig. Life Extrem. Habitats Astrobiol. 2011, 19, 119–142. [Google Scholar]
- Yamada, M.; Otsubo, M.; Tsutsumi, Y.; Mizota, C.; Nakamura, Y.; Takahashi, K.; Iwataki, M. Utility of mitochondrial-encoded cytochrome coxidase I gene for phylogenetic analysis and species identification of the planktonic diatom genus Skeletonema. Physiol. Res. 2017, 66, 217–225. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The Clustal_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. Bioedit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/nt. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Belcher, J.H.; Swale, E.M.F. Skeletonema potamos (Weber) Hasle and Cyclotella atomus Hustedt (Bacillariophyceae) in the plankton of rivers in England and France. Br. Phycol. J. 1978, 13, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Steinberg, C.; Heindel, B.; Tille-Backhaus, R. Phytoplanktonstudien an langsamfließenden Gewässern: Donau und Vils. Arch. Für Hydrobiol. 1987, 68, 437–456. [Google Scholar]
- Bergesch, M.; Garcia, M.; Odebrecht, C. Diversity and morphology of Skeletonema species in Southern Brazil, Southwestern Atlantic Ocean. J. Phycol. 2009, 45, 1348–1352. [Google Scholar] [CrossRef]
- Chessman, B.C. Phytoplankton of the La Trobe River, Victoria. Aust. J. Mar. Freshw. Res. 1985, 36, 115–122. [Google Scholar] [CrossRef]
- Kiss, K.T.; Ács, É.; Kovács, A. Ecological observations on Skeletonema potamos (Weber) Hasle in the River Danube, near Budapest (1991–92, daily investigations). Hydrobiologia 1994, 289, 163–170. [Google Scholar]
- Cavalcante, K.P.; Tremarin, P.I.; Ludwig, T.A. Taxonomic studies of centric diatoms (Diatomeae): Unusual nanoplanktonic forms and new records for Brazil. Acta Bot. Bras. 2013, 27, 327–351. [Google Scholar] [CrossRef] [Green Version]
- Descy, J.; Leitão, M.; Everbecq, E.; Smitz, J.; Deliege, J. Phytoplankton of the River Loire, France: A biodiversity and modelling study. J. Plankton Res. 2012, 34, 120–135. [Google Scholar] [CrossRef] [Green Version]
- Pfannkuchen, D.M.; Godrijan, J.; Tanković, M.S.; Baričević, A.; Kužat, N.; Djakovac, T.; Pustijanac, E.; Jahn, R.; Pfannkuchen, M. The ecology of one cosmopolitan, one newly introduced and one occasionally advected species from the Genus Skeletonema in a highly structured ecosystem, the Northern Adriatic. Environ. Microbiol. 2018, 75, 674–687. [Google Scholar] [CrossRef]
- Liu, D.Y.; Jiang, J.J.; Wang, Y.; Zhang, Y.; Di, B.P. Large scale northward expansion of warm water species Skeletonema tropicum (Bacillariophyceae) in China seas. Chin. J. Oceanol. Limnol. 2012, 30, 519–527. [Google Scholar] [CrossRef] [Green Version]
- Du, L.N.; Zhang, J.B. The cooperation relationship study on the Golden Waterway of the Yangtze River and economic development along the Yangtze River. AIP Conf. Proc. 2017, 1890, 040021. [Google Scholar]
- Yamada, M.; Otsubo, M.; Tsutsumi, Y.; Mizota, C.; Iida, N.; Okamura, K.M.; Umehara, A. Species diversity of the marine diatom genus Skeletonema in Japanese brackish water areas. Fish. Sci. 2013, 79, 923–934. [Google Scholar] [CrossRef]
- Numazawa, A. Relationship between Productivity and Transparency Fall Discovered in Water Quality Monitoring and Plankton Analysis in Lake Kasumigaura Ecosystem. In Proceedings of the 11th World Lake Conference, Nairobi, Kenya, 31 October–4 November 2005; Odada, E.O., Olago, D.O., Ochola, W., Ntiba, M., Wandiga, S., Gichuki, N., Oyieke, H., Eds.; Ministry of Water and Irrigation, Kenya, ILEC: Nairobi, Kenya, 2005; pp. 297–300. [Google Scholar]
- Main, S.P. Benthic diatom distribution in the Cedar River Basin, Iowa. Proc. Iowa Acad. Sci. 1977, 84, 23–29. [Google Scholar]
- Sabater, S.; Muñoz, I. Successional dynamics of the phytoplankton in the lower part of the river Ebro. J. Plankton Res. 1990, 12, 573–592. [Google Scholar] [CrossRef]
- Chang, T.P.; Steinberg, C. Seasonal changes in the diatom flora in a small reservoir with special reference to Skeletonema potamos. Diatom Res. 1988, 3, 191–201. [Google Scholar] [CrossRef]
- Mihaljević, M.; Špoljarić, D.; Stević, F.; Pfeiffer, T.Ž. Assessment of flood-induced changes of phytoplankton along a river–floodplain system using the morpho-functional approach. Environ. Monit. Assess. 2013, 185, 8601–8619. [Google Scholar] [CrossRef] [PubMed]
- Pérez, M.C.; Maidana, N.I.; Comas, A. Phytoplankton composition of the Ebro River estuary, Spain. Acta Bot. Croat. 2009, 68, 11–27. [Google Scholar]
- Hamilton, P.B.; Lavoie, I.; Ley, L.M.; Poulin, M. Factors contributing to the spatial and temporal variability of phytoplankton communities in the Rideau River (Ontario, Canada). River Syst. 2011, 19, 189–205. [Google Scholar] [CrossRef]
- Admiraal, W.; Breebaart, L.; Tubbing, G.M.; Zanten, B.V.; Steveninck, E.D.; Bijkerk, R. Seasonal variation in composition and production of planktonic communities in the lower River Rhine. Freshw. Biol. 1994, 32, 519–531. [Google Scholar] [CrossRef]
- Lomas, M.W.; Glibert, P.M. Temperature regulation of nitrate uptake: A novel hypothesis about nitrate uptake and reduction in cool-water diatoms. Limnol. Oceanogr. 1999, 44, 556–572. [Google Scholar] [CrossRef]
- Cloern, J.E.; Dufford, R. Phytoplankton community ecology: Principles applied in San Francisco Bay. Mar. Ecol. Prog. 2005, 285, 11–28. [Google Scholar] [CrossRef]
- Gosselain, V.; Descy, J.P.; Everbecq, E. The phytoplankton community of the River Meuse, Belgium:seasonal dynamics (year 1992) and the possible incidence of zooplankton grazing. Hydrobiologia 1994, 289, 179–191. [Google Scholar] [CrossRef]
- Devercelli, M.; Farrell, I.O. Factors affecting the structure and maintenance of phytoplankton functional groups in a nutrient rich lowland river. Limnologica 2013, 43, 67–78. [Google Scholar] [CrossRef]
- Devercelli, M. Phytoplankton of the Middle Paraná River during an anomalous hydrological period: A morphological and functional approach. Hydrobiologia 2006, 563, 465–478. [Google Scholar] [CrossRef]
- Lehman, P.W.; The, S.J.; Boyer, G.L.; Nobriga, M.L.; Bass, E.; Hogle, C. Initial impacts of Microcystis aeruginosa blooms on the aquatic food web in the San Francisco Estuary. Hydrobiologia 2010, 637, 229–248. [Google Scholar] [CrossRef] [Green Version]
- Brown, L.R.; May, J.T. Periphyton and Macroinvertebrate Communities at Five Sites in the San Joaquin River Basin, California, during June and September, 2001; US Geological Survey: Reston, Virginia, 2004. [Google Scholar]
- Lehman, P.W. The influence of phytoplankton community composition on primary productivity along the riverine to freshwater tidal continuum in the San Joaquin River, California. Estuar Coast 2007, 30, 82–93. [Google Scholar] [CrossRef]
- Tavernini, S.; Pierobon, E.; Viaroli, P. Physical factors and dissolved reactive silica affect phytoplankton community structure and dynamics in a lowland eutrophic river (Po river, Italy). Hydrobiologia 2011, 669, 213–225. [Google Scholar] [CrossRef]
- Čađo, S.; Miletić, A.; Dopuđa-Glišić, T.; Denić, L. Physical-Chemical Characteristics and Phytoplankton Composition of the Sava River on Its Lower Flow Stretch through Serbia. In Proceedings of the 36th International Conference of IAD, Klosterneuburg, Austria, 4–8 September 2006; Austrian Committee DanubeResearch/IAD Vienna: Vienna, Austria, 2006; pp. 184–188. [Google Scholar]
- Marshall, H.G.; Alden, R.W. A comparison of phytoplankton assemblages and environmental relationships in three estuarine rivers of the Lower Chesapeake Bay. Estuaries 1990, 13, 287–300. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Markers | Primers Names | Primers Sequences (5′–3′) |
|---|---|---|
| cox1 | Fd | GGAACTTTATATTTAATCTTTGGTGC |
| 1047R | AATACCAGAATTAGCAAGAACAAC | |
| 424F | GCGTCTTCTATTTTAGGTG | |
| 644R | GGATCACCTCCACCACCAGC | |
| nLSU rDNA | D1R | ACCCGCTGAATTTAAGCATA |
| D2C | CCTTGGTCCGTGTTTCAAGA | |
| nSSU rDNA | 11F | TGATCCTGCCAGTAGTCATACGCT |
| 1147R | AGTTTCAGCCTTGCGACCATAC | |
| rbcL | 66F | TTAAGGAGAAATAAATGTCTCAATCTG |
| 1255R | TTGGTGCATTTGACCACAGT | |
| psbC | 22F | CGTGGTGATACATAGTTA |
| 1154R | GCDCATGCTGGTTTAATGG |
| cox1 | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| 1 LC192721 FCH102 (Japan) | 0.000 | 0.001 | 0.001 | 0.001 | 0.000 | 0.000 | |
| 2 LC192723 FCH106 (Japan) | 0.000 | 0.001 | 0.001 | 0.001 | 0.000 | 0.000 | |
| 3 AB706249 FCH024 (Japan) | 0.001 | 0.001 | 0.000 | 0.002 | 0.001 | 0.001 | |
| 4 LC192720 FCH101 (Japan) | 0.001 | 0.001 | 0.000 | 0.002 | 0.001 | 0.001 | |
| 5 LC192731 FCH159 (Japan) | 0.002 | 0.002 | 0.003 | 0.003 | 0.001 | 0.001 | |
| 6 LC192732 FIS101 (Japan) | 0.000 | 0.000 | 0.001 | 0.001 | 0.002 | 0.000 | |
| 7 OP699718 cjh2 (China) | 0.000 | 0.000 | 0.001 | 0.001 | 0.002 | 0.000 | |
| rbcL | 1 | 2 | 3 | ||||
| 1 KJ081746 AJA010-19 (USA) | 0.001 | 0.000 | |||||
| 2 KF621301 (Hungary) | 0.001 | 0.001 | |||||
| 3 OP819055 cjh2 (China) | 0.000 | 0.001 |
| Chloroplast | Frustule Diameter (µm) | Pervalvar Axis (µm) | Distance between Cells (µm) | Number of FPPs | RP | |
|---|---|---|---|---|---|---|
| Observed | 1–2 | 1.3–3 | 4–12 | 0.2–0.4 | 3–6 | 1 |
| Weber (1970) [7] | several | 3–4 | 4–8 | — | 5–8 | — |
| Hasle and Evensen (1976) [6] | 1–2 (4) | 3–4 | — | — | 6–8 | 1 |
| Belcher and Swale (1978) [25] | 1–2 | 3–4 | 6–10 | 0, >1 | 5–6 | 1 |
| Torgan et al. (2009) [9] | 1–2 | 3–4, 5 | 6–10 | — | 5–7 | 1 |
| Kiss et al. (2012) [30] | — | 3–6.5 | 5–18 | — | 3–8 | 1 |
| Cavalcante et al. (2013) [31] | — | 2.4–3.6 | 4.4–7.3 | short | 4–8 | 1 |
| Location | WT (°C) | Sal (psu) | TP (mg L−1) | COD (mg L−1) | Reference |
|---|---|---|---|---|---|
| Tidal area of the Chikugo River (Japan) | 28.2 | 13.0 | — | — | Yamada et al., 2013 [36] |
| Lake Kasumigaura (Japan) | 11.0–18.0 (D) | — | 0.05–0.175 (D) | 5–7 (D) | Numazawa, 2005 [37] |
| Cedar River Basin (USA) | 11.0–16.0 | — | 0.9–1.9 | — | Main, 1977 [38] |
| Patos Lagoon (Brazil) | 19.0 | 4.0 | — | — | Bergesch et al., 2009 [28] |
| River Ebro (Spain) | 28.0 | — | — | — | Sabater and Muñoz, 1990 [39] |
| River Rott-Postmünster (Germany) | 17.7–19.8 | — | — | — | Chang and Steinberg, 1988 [40] |
| River Cachoeira (Brazil) | 24.6 (M) | — | — | — | Cavalcante et al., 2013 [31] |
| Patos Lagoon (southern Brazil) | 14.0–24.0 | 0.0–7.2 | — | — | Lezilda et al., 2011 [9] |
| River Danube, near Budapest (Hungary) | 14.0–25.0 | — | — | — | Kiss et al., 1994 [30] |
| River Danube at Göd (Hungary) | 14.9–27.0 | — | — | — | Duleba et al., 2014 [1] |
| River Danube (Croatia) | 8.0–23.0 | — | — | — | Mihaljević et al., 2013 [41] |
| Little Miami River (OH, USA) | 6.0–29.0 | — | 0.455 | — | Weber, 1970 [7] |
| Estuary of the River Ebro (Spain) | 9.8–27.3 | 2.3–5.1 | — | — | Pérez et al., 2009 [42] |
| Rideau River (ON, Canada) | 11.9–26.8 | — | 0.037 | — | Hamilton et al., 2011 [43] |
| River Rhine (Netherlands) | — | 0.2–0.9 | — | — | Admiraal et al., 1994 [44] |
| Chesapeake Bay (Virginia, USA) | 14.1 | — | — | — | Lomas and Glibert, 1999 [45] |
| San Francisco Bay (CA, USA) | 17.0–22.0 | 0.0–5.0 | — | — | Cloern and Dufford, 2005 [46] |
| River Meuse at Namur (Belgium) | 12.0–22.0 (D) | — | — | — | Gosselain et al., 1994 [47] |
| Salado River, El Vado stream (Argentina) | 11.0–30.0 | — | 0.52–0.63 | — | Devercelli and Farrell., 2013 [48] |
| Parana’ River (Argentina) | 12.0–29.0 | — | — | — | Devercelli, 2006 [49] |
| Suisun Bay (USA) | 21.5 | — | — | — | Lehman et al., 2010 [50] |
| San Joaquin River (CA, USA) | 24.4–25.6 | — | — | — | Brown and May, 2004 [51] |
| San Joaquin River (CA, USA) | 21.0–25.5 | — | 0.14 | — | Lehman, 2007 [52] |
| Po River (Italy) | — | — | 0.302 | — | Tavernini et al., 2011 [53] |
| Sava River (Republic of Serbia) | — | — | 0.067–0.092 | 2.45–3.45 | Čađo et al., 2006 [54] |
| Moselle River (France) | — | — | 0.1 | 19.6 | Heudre et al., 2020 [8] |
| Estuary of River York (VA, USA) | — | — | 0.01–0.13 | — | Marshall et al., 1990 [55] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Yang, Z.; Yi, Y.; Shu, Z.; Yu, P.; You, Q.; Wang, Q. Possible Origin and Distribution of an Invasive Diatom Species, Skeletonema potamos, in Yangtze River Basin (China). Water 2023, 15, 2875. https://doi.org/10.3390/w15162875
Hu J, Yang Z, Yi Y, Shu Z, Yu P, You Q, Wang Q. Possible Origin and Distribution of an Invasive Diatom Species, Skeletonema potamos, in Yangtze River Basin (China). Water. 2023; 15(16):2875. https://doi.org/10.3390/w15162875
Chicago/Turabian StyleHu, Jingwen, Zhengxin Yang, Yuxin Yi, Zhaoqing Shu, Pan Yu, Qingmin You, and Quanxi Wang. 2023. "Possible Origin and Distribution of an Invasive Diatom Species, Skeletonema potamos, in Yangtze River Basin (China)" Water 15, no. 16: 2875. https://doi.org/10.3390/w15162875
APA StyleHu, J., Yang, Z., Yi, Y., Shu, Z., Yu, P., You, Q., & Wang, Q. (2023). Possible Origin and Distribution of an Invasive Diatom Species, Skeletonema potamos, in Yangtze River Basin (China). Water, 15(16), 2875. https://doi.org/10.3390/w15162875