

Elucidating the Potential of Dye-Degrading Enterobacter cloacae ZA14 for Cultivation of Solanum lycopersicum Plants with Textile Effluents


 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Dye Decolorization Assay
2.2. Growth and Biochemical Characterization of Selected Strains
2.3. Sand Culture Experiment on Tomato Plants
2.3.1. Estimation of Plant Growth and Physiological Parameters
2.3.2. Oxidative Stress Markers
2.3.3. Osmoprotectants
2.3.4. Secondary Metabolites
2.3.5. Antioxidant Production
2.4. Statistical Analysis
3. Results
3.1. Dye Decolorization Assay for Isolation of Bacterial Endophytes
3.2. Growth and Biochemical Characterization of Bacterial Strains
3.3. Minimum Inhibitory Concentration (MIC)
3.4. Identification of Bacterial Strain
3.5. Sand Culture Experiment
3.5.1. Estimation of Plant Growth and Physiological Parameters
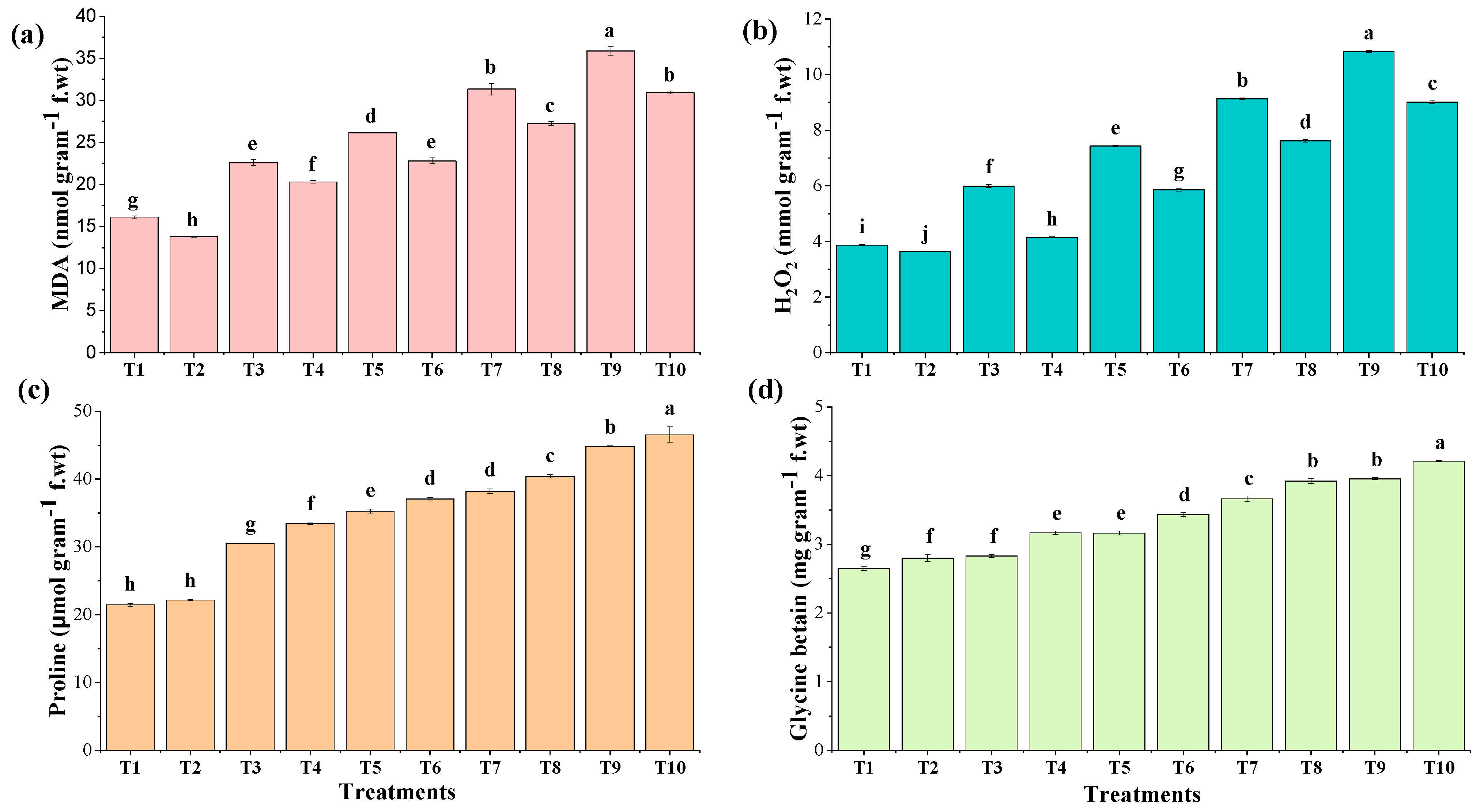
3.5.2. Oxidative Stress Markers
3.5.3. Osmoprotectants
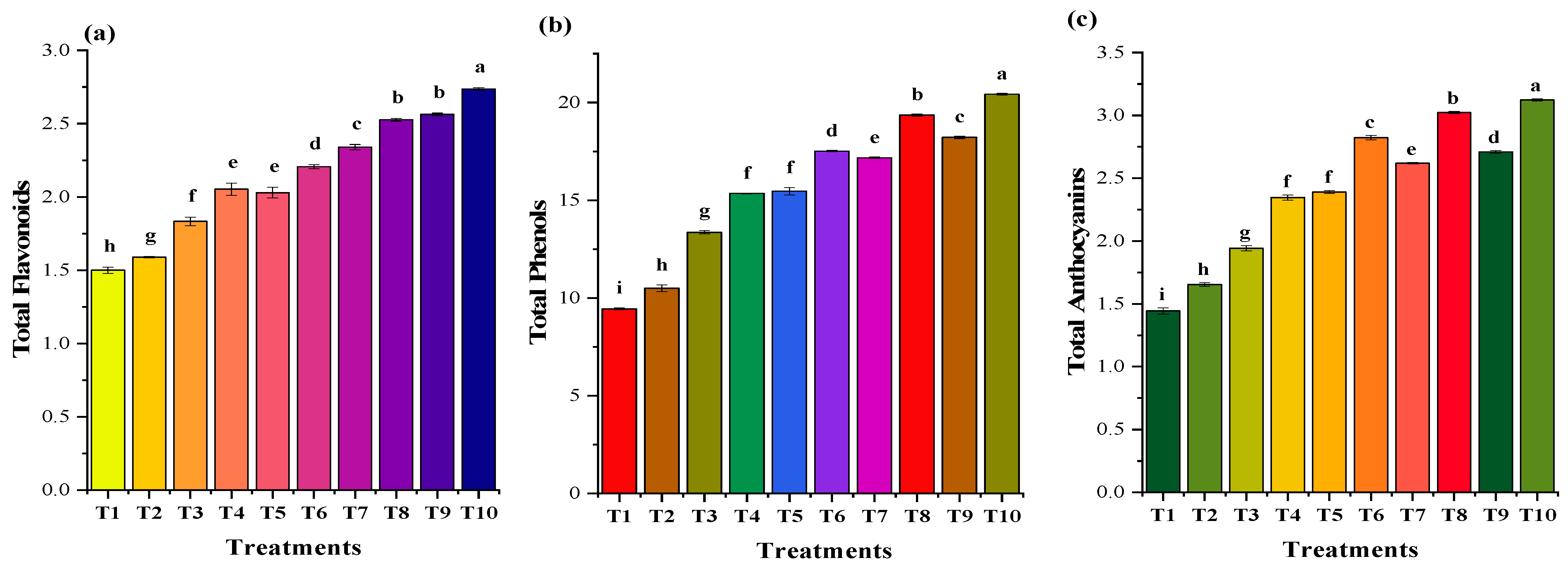
3.5.4. Secondary Metabolites
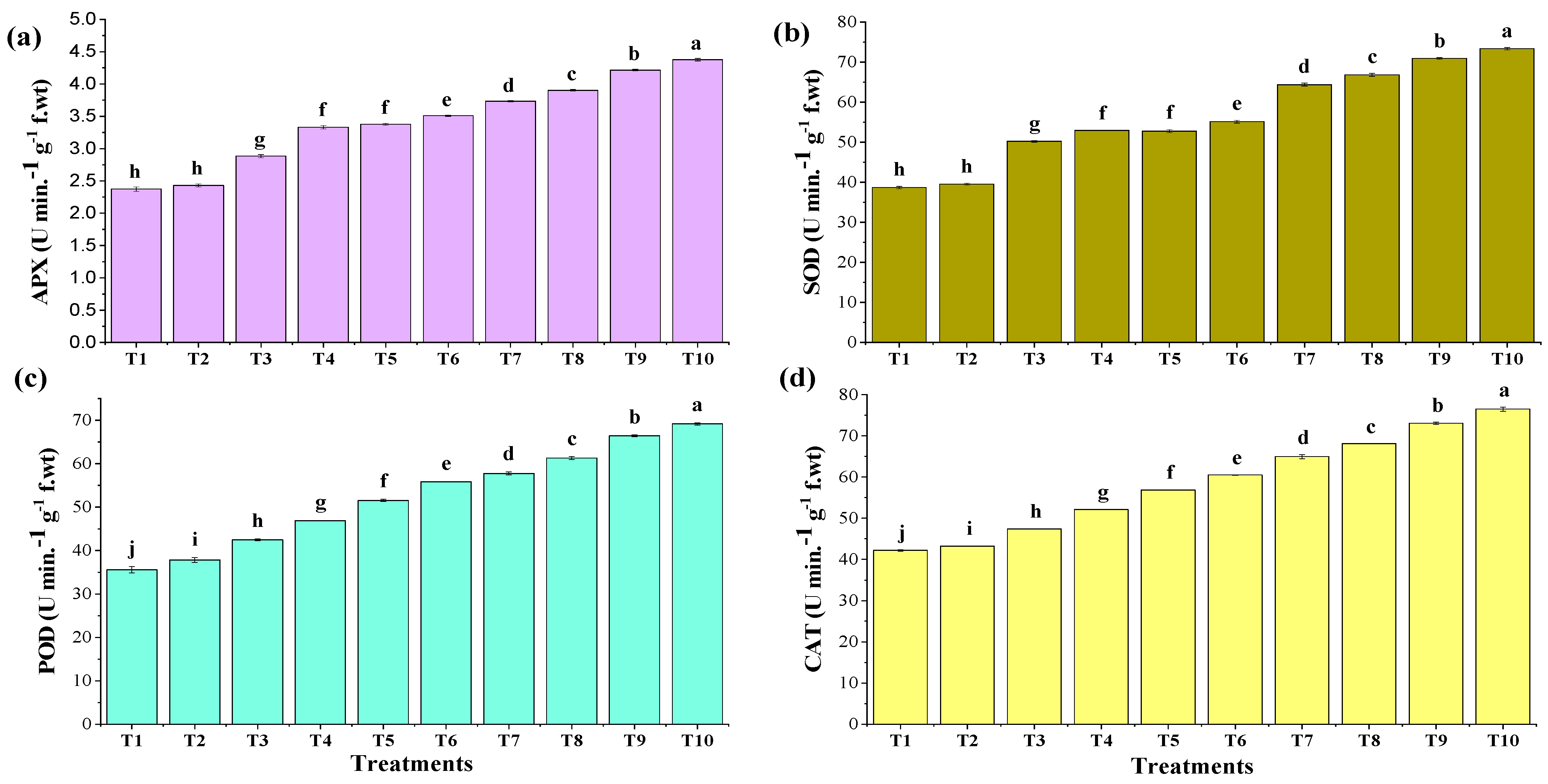
3.5.5. Antioxidant Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eslami, H.; Shariatifar, A.; Rafiee, E.; Shiranian, M.; Salehi, F.; Hosseini, S.S.; Eslami, G.; Ghanbari, R.; Ebrahimi, A.A. Decolorization and biodegradation of reactive Red 198 Azo dye by a new Enterococcus faecalis–Klebsiella variicola bacterial consortium isolated from textile wastewater sludge. World J. Microbiol. Biotechnol. 2019, 35, 38. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Sharma, R.K.; Agrawal, M.; Marshall, F.M. Health risk assessment of heavy metals via dietary intake of foodstuffs from the wastewater irrigated site of a dry tropical area of India. Food Chem Toxicol. 2010, 8, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Najam-us-Sahar, H.A.; Mustafa, A.; Waqas, R.; Ashraf, I.; Akhtar, M.F.U.Z. Effect of textile wastewater on growth and yield of wheat (Triticum aestivum L.). Soil Environ. 2017, 36, 28–34. [Google Scholar]
- Chandanshive, V.V.; Rane, N.R.; Tamboli, A.S.; Gholave, A.R.; Khandare, R.V.; Govindwar, S.P. Co-plantation of aquatic macrophytes Typha angustifolia and Paspalum scrobiculatum for effective treatment of textile industry effluent. J. Hazard. Mater. 2017, 338, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Javaid, S. Heavy metals stress, mechanism and remediation techniques in rice (Oryza sativa L.): A review. Pure Appl. Biol. 2020, 9, 403–426. [Google Scholar] [CrossRef]
- Sabir, M.; Naseem, Z.; Ahmad, W.; Usman, M.; Nadeem, F.; Saifullah; Ahmad, H.R. Alleviation of adverse effects of nickel on growth and concentration of copper and manganese in wheat through foliar application of ascorbic acid. Int. J. Phytoremediation 2022, 24, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Naseem, Z.; Naveed, M.; Asghar, H.N.; Hameed, M. Metal resistant Enterobacter cloacae ZA14 enhanced seedling vigor and metal tolerance through improved growth, physiology and antioxidants in tomato (Solanum lycopersicum) irrigated with textile effluents. Sustainability 2022, 14, 13619. [Google Scholar] [CrossRef]
- Filipovic, M.R.; Zivanovic, J.; Alvarez, B.; Banerjee, R. Chemical biology of H2S signaling through persulfidation. Chem. Rev. 2018, 118, 1253–1337. [Google Scholar] [CrossRef]
- Banerjee, A.; Ghoshal, A.K. Biodegradation of an actual petroleum wastewater in a packed bed reactor by an immobilized biomass of Bacillus cereus. J. Environ. Chem. Eng. 2017, 5, 1696–1702. [Google Scholar] [CrossRef]
- Naveed, M.; Mitter, B.; Yousaf, S.; Pastar, M.; Afzal, M.; Sessitsch, A. The endophyte Enterobacter sp. FD17: A maize growth enhancer selected based on rigorous testing of plant beneficial traits and colonization characteristics. Biol. Fertil. Soils 2014, 50, 249–262. [Google Scholar] [CrossRef]
- Andreolli, M.; Lampis, S.; Poli, M.; Gullner, G.; Biró, B.; Vallini, G. Endo-phytic Burkholderia fungorum DBT1 can improve phytoremediation efficiency of polycyclic aromatic hydrocarbons. Chemosphere 2013, 92, 688–694. [Google Scholar] [CrossRef]
- Myo, E.M.; Ge, B.; Ma, J.; Cui, H.; Liu, B.; Shi, L.; Jiang, M.; Zhang, K. Indole-3-acetic acid production by Streptomyces fradiae NKZ-259 and its formulation to enhance plant growth. BMC Microbiol. 2019, 19, 155. [Google Scholar] [CrossRef]
- Vaishnav, A.; Shukla, A.K.; Sharma, A.; Kumar, R.; Choudhary, D.K. Endophytic bacteria in plant salt stress tolerance: Current and Future Prospects. J. Plant Growth Regul. 2018, 38, 650–668. [Google Scholar] [CrossRef]
- Shafqat, M.; Khalid, A.; Mahmood, T.; Muhammad, T.; Siddique, M.T.; Jong-In, H.; Habteselassiee, M.Y. Evaluation of bacteria isolated from textile wastewater and rhizosphere to simultaneously degrade azo dyes and promote plant growth. J. Chem. Technol. Biotechnol. 2017, 92, 2760–2768. [Google Scholar] [CrossRef]
- Prasad, A.; Rao, K.B. Physico chemical characterization of textile effluent and screening for dye decolorizing bacteria. Glob. J. Biotechnol. Biochem. 2011, 5, 80–86. [Google Scholar]
- Saratale, G.D.; Kalme, S.D.; Govindwar, S.P. Decolorization of textile dyes by Aspergillus ochraceus. Ind. J. Biotechnol. 2006, 5, 407–410. [Google Scholar]
- Holt, J.G.; Krieg, N.R.; Sneath, P.H.A.; Stanley, J.T.; William, S.T. Bergey’s Manual of Determinative Bacteriology; Williams and Wilikins: Baltimore, MD, USA, 1994; pp. 786–788. [Google Scholar]
- Pillai, H.P.J.S.; Girish, K.; Agsar, D. Isolation, characterization and screening of actinomycetes from textile industry effluent for dye degradation. Int. J. Curr. Microbiol. App. Sci. 2014, 3, 105–115. [Google Scholar]
- MacFaddin, J.F. Biochemical Tests for Identification of Medical Bacteria, 2nd ed.; Williams and Wilkins: Baltimore, MD, USA, 1980. [Google Scholar]
- Ngamau, C.N.; Matiru, V.N.; Tani, A.; Muthuri, C.W. Isolation and identification of endophytic bacteria of bananas (Musa spp.) in Kenya and their potential as biofertilizers for sustainable banana production. Afric. J. Microbiol. Res. 2012, 6, 6414–6422. [Google Scholar]
- Mustafa, Z.M.; Malik, N. Isolation and Characterization of Heavy Metal Resistant Plant Growth Promoting Bacteria. IOSR J. Pharm. 2019, 9, 18–24. [Google Scholar]
- Nguyen, C.; Yan, W.; Tacon, F.L.; Lapeyrie, F. Genetic variability of phosphate solubilizing activity by monocaryotic and dicaryotic mycelia of the ectomycorrhizal fungus Laccaria bicolor (Maire) P.D. Orton. Plant Soil 1992, 143, 193–199. [Google Scholar] [CrossRef]
- Tadashi, A. Culture media for actinomycetes. The society for actinomycetes. Jpn. Natl. Agric. Lib. 1975, 1, 31. [Google Scholar]
- Shirling, E.B.; Gottlieb, D. Methods of characterization of Streptomyces species. Int. J. Syst. Bacteriol. 1996, 61, 313–340. [Google Scholar] [CrossRef]
- Lorck, H.; Veterinary, R. Production of hydrocyanic acid by bacteria. Physiol. Plant. 1948, 1, 142–146. [Google Scholar] [CrossRef]
- Gebreyohannes, G. Isolation and optimization of amylase producing bacteria and actinomycetes from soil samples of Maraki and Tewedros campus, University of Gondar, North West Ethiopia. Afr. J. Microbiol. Res. 2015, 9, 1877–1882. [Google Scholar]
- Sarwar, M.; Arshad, M.; Martens, D.A.; Frankenberger, W.T. Tryptophan dependent biosynthesis of auxins in soil. Plant Soil 1992, 147, 207–215. [Google Scholar] [CrossRef]
- Cappuccino, J.G.; Sherman, N. Biochemical Activities of Microorganisms. Microbiology, a Laboratory Manual; The Benjamin Cummings Publishing: San Francisco, CA, USA, 2013. [Google Scholar]
- Barathi, S.; Arulselvi, P.I. Isolation and characterization of textile dye degrading native bacterial strains from textile effluent contaminated sites. Int. J. Multidiscip. Res. Dev. 2015, 2, 155–160. [Google Scholar]
- Kuffner, M.; Puschenreiter, M.; Wieshammer, G.; Gorfer, M.; Sessitsch, A. Rhizosphere bacteria affect growth and metal uptake of heavy metal accumulating willows. Plant Soil 2008, 304, 35–44. [Google Scholar] [CrossRef]
- Weatherly, P.E. Studies in water relations in cotton plants. The field measurement of water deficit in leaves. New Phytol. 1950, 49, 81–87. [Google Scholar] [CrossRef]
- Yagi, K. Assay for serum lipid peroxide level and its clinical significance. In Lipid Peroxides in Biology and Medicine; Yagi, K., Ed.; Academic Press: New York, NY, USA, 1982; Volume 223, p. 242. [Google Scholar]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain treated bean plants. Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S.R. Rapid assay for determination of water soluble quaternary ammonium compounds. Plant Soil 1983, 70, 303–307. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Mancinelli, A.L. Photoregulation of Anthocyanin Synthesis. Plant Physiol. 1984, 75, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Chance, B.; Maehly, A.C. Assay of catalase and peroxidase. Meth. Enzymol. 1955, 2, 764–775. [Google Scholar]
- Giannopolities, C.N.; Ries, S.K. Superoxidae dismutase. I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Asada, K.; Takahashi, M. Production and scavenging of active oxygen in photosynthesis. In Photoinhibition; Kyle, D.J., Osmond, C.B., Arntzen, O., Eds.; Elsevier: Amsterdam, The Netherlands, 1987; pp. 227–289. [Google Scholar]
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio, PBC: Boston, MA, USA, 2021; Available online: http://www.rstudio.com/ (accessed on 19 July 2022).
- Nazir, A.; Wahid, A.; Nawaz, S.; Gulshan, A.B.; Leghari, S.K.; Hussain, F.; Nijabat, A.; Khan, M.A.; Awan, A.N.; Shafqat, U.; et al. Vicissitudes in Morphological and Photosynthetic Attributes in Maize (Zea mays) plant by elevating the Cobalt Concentration in soil. GU J. Phytosci. 2022, 2, 114–120. [Google Scholar]
- Chanwala, J.; Kaushik, G.; Ashraf, D.M.; Upadhyay, S.; Agrawal, A. Process optimization and enhanced decolorization of textile effluent by Planococcus sp. isolated from textile sludge. Environ. Technol. Innov. 2019, 13, 122–129. [Google Scholar] [CrossRef]
- Selim, M.T.; Salem, S.S.; Mohamed, A.A.; El-Gamal, M.S.; Awad, M.F.; Fouda, A. Biological treatment of real textile effluent using aspergillus flavus and fusarium oxysporium and their consortium along with the evaluation of their phytotoxicity. J. Fungi 2021, 7, 193. [Google Scholar] [CrossRef]
- Holey, B.A. Decolourization of Congo Red dye by bacteria and consortium isolated from dye contaminated soil. Int. Res. J. Sci. Eng. 2015, 3, 107–112. [Google Scholar]
- Roy, D.C.; Biswas, S.K.; Saha, A.K.; Sikdar, B.; Rahman, M.; Roy, A.K.; Prodhan, Z.H.; Tang, S. Biodegradation of Crystal Violet dye by bacteria isolated from textile industry effluents. PeerJ. 2018, 6, e5015. [Google Scholar] [CrossRef]
- Das, S.; Dash, H.R. Handbook of Metal-Microbe Interactions and Bioremediation; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Hakim, S.; Naqqash, T.; Nawaz, M.S.; Laraib, I.; Siddique, M.J.; Zia, R.; Mirza, M.S.; Imran, A. Rhizosphere Engineering with Plant Growth-Promoting Microorganisms for Agriculture and Ecological Sustainability. Front. Sustain. Food Syst. 2021, 5, 617157. [Google Scholar] [CrossRef]
- Naveed, M.; Mustafa, A.; Majeed, S.; Naseem, Z.; Saeed, Q.; Khan, A.; Nawaz, A.; Baig, K.S.; Jen-Tsung, C. Enhancing Cadmium Tolerance and Pea Plant Health through Enterobacter sp. MN17 Inoculation Together with Biochar and Gravel Sand. Plants 2020, 9, 530. [Google Scholar] [CrossRef] [PubMed]
- Weilharter, A.; Mitter, B.; Shin, M.V.; Chain, P.S.; Nowak, J.; Sessitsch, A. Complete genome sequence of the plant growth-promoting endophyte Burkholderia phytofirmans strain PsJN. J. Bacteriol. 2011, 193, 3383–3384. [Google Scholar] [CrossRef] [PubMed]
- Krause, A.; Ramakumar, A.; Bartels, D.; Battistoni, F.; Bekel, T.; Boch, J.; Böhm, M.; Friedrich, F.; Hurek, T.; Krause, L.; et al. Complete genome of the mutualistic, N2-fixing grass endophyte Azoarcus sp. strain BH72. Nat. Biotechnol. 2006, 24, 1385–1391. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski-Dyé, F.; Borziak, K.; Khalsa-Moyers, G.; Alexandre, G.; Sukharnikov, L.O.; Wuichet, K.; Hurst, G.B.; McDonald, W.H.; Robertson, J.S.; Barbe, V.; et al. Azospirillum genomes reveal transition of bacteria from aquatic to terrestrial environments. PLoS Genet. 2011, 7, e1002430. [Google Scholar] [CrossRef]
- Gulati, A.; Sharma, N.; Vyas, P.; Sood, S.; Rahi, P.; Pathania, V.; Prasad, R. Organic acid production and plant growth promotion as a function of phosphate solubilization by Acinetobacter rhizosphaerae strain BIHB 723 isolated from the cold deserts of the trans-Himalayas. Arch. Microbiol. 2010, 192, 975–983. [Google Scholar] [CrossRef]
- Szymańska, S.; Borruso, L.; Brusetti, L.; Hulisz, P.; Furtado, B.; Hrynkiewicz, K. Bacterial microbiome of root-associated endophytes of Salicornia europaea in correspondence to diferent levels of salinity. Environ. Sci. Pollut. Res. 2018, 25, 25420–25431. [Google Scholar] [CrossRef]
- Yaish, M.W.; Al-Lawati, A.; Jana, G.A.; Vishwas, H.P.; Glick, B.R. Impact of soil salinity on the structure of the bacterial endophytic community identifed from the roots of Caliph Medic (Medicago truncatula). PLoS ONE 2015, 11, e0159007. [Google Scholar]
- Joe, M.M.; Devaraj, S.; Benson, A.; Sa, T. Isolation of phosphate solubilizing endophytic bacteria from Phyllanthus amarus S-chum & Thonn: Evaluation of plant growth promotion and antioxidant activity under salt stress. J. Appl. Res. Med. Aromat. Plants 2016, 3, 71–77. [Google Scholar]
- Rajkumar, M.; Sandhya, S.; Prasad, M.N.V.; Freitas, H. Perspectives of plant-associated microbes in heavy metal phytoremediation. Biotechnol. Adv. 2012, 30, 1562–1574. [Google Scholar] [CrossRef] [PubMed]
- Pramanik, K.; Mitra, S.; Sarkar, A.; Soren, T.; Maiti, T.K. Characterization of a Cd2þ-resistant plant growth promoting rhizobacterium (Enterobacter sp.) and its effects on rice seedling growth promotion under Cd2þ-stress in vitro. Agric. Nat. Res. 2018, 52, 215–221. [Google Scholar]
- Karthik, C.; Elangovan, N.; Kumara, T.S.; Govindharaju, S.; Barathi, S.; Oves, M.; Arulselvia, P.I. Characterization of multifarious plant growth promoting traits of rhizobacterial strain AR6 under Chromium (VI) stress. Microbiol. Res. 2017, 204, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Wins, J.A.; Murugan, M. Effect of textile mill effluent on growth and germination of black gram-Vigna mungo (L.). Int. J. Pharma Bio Sci. Hepper. 2010, 1, 1–7. [Google Scholar]
- Aydinalp, C.; Marinova, S. The Effects of heavy metals on seed germination and plant growth on alfalfa plant (Medicago sativa). Bulg. J. Agric. Sci. 2009, 15, 347–350. [Google Scholar]
- Woldeamanuale, T.B. Investigation the Impact of Textile Industrial waste waters on the Soil and Plants. J. Environ. Sci. Toxicol. Food Technol. 2021, 15, 9–12. [Google Scholar]
- Jamee, R.; Siddique, R. Biodegradation of synthetic dyes of textile effluent by microorganisms: An environmentally and economically sustainable approach. Eur. J. Microbiol. Immunol. 2019, 9, 114–118. [Google Scholar] [CrossRef]
- Parvin, K.; Hasanuzzaman, M.; Borhannuddin, B.M.H.M.; Nahar, K.; Mohsin, S.M.; Fujita, M. Comparative Physiological and Biochemical Changes in Tomato (Solanum lycopersicum L.) under Salt Stress and Recovery: Role of Antioxidant Defense and Glyoxalase Systems. Antioxidants 2019, 8, 350. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Gill, S.S.; Alharby, H.F.; Razafindrabe, B.H.; Fujita, M. Hydrogen Peroxide pretreatment mitigates cadmium-induced oxidative stress in Brassica napus L. An intrinsic study on antioxidant defense and glyoxalase systems. Front. Plant Sci. 2017, 8, 115. [Google Scholar] [CrossRef]
- Seyedi, Z.S.; Zahraei, Z.; Kashi, F.J. Decolorization of reactive black 5 and reactive red 152 azo dyes by new haloalkaliphilic bacteria isolated from the textile Wastewater. Curr. Microbiol. 2020, 77, 2084–2092. [Google Scholar] [CrossRef]
- El-Sayed, M.H. Multiple Heavy Metal and Antibiotic Resistance of Acinetobacter baumanni Strain HAFC 13 Isolated from Industrial Effluents. J. Microbiol. Res. 2016, 4, 26–36. [Google Scholar]
- Samanta, A.; Bera, P.; Khatun, M.; Sinha, C.; Pal, P.; Lalee, A.; Mandal, A. An investigation on heavy metal tolerance and antibiotic resistance properties of bacterial strain Bacillus sp. isolated from municipal waste. J. Microbiol. Biotech. Res. 2012, 2, 178–189. [Google Scholar]
- Rizvi, A.; Ahmed, B.; Zaidi, A.; Khan, M.A. Heavy metal mediated phytotoxic impact on winter wheat: Oxidative stress and microbial management of toxicity by Bacillus subtilis BM2. RSC Adv. 2019, 9, 6125–6142. [Google Scholar] [CrossRef]
- Singh, R.; Singh, S.; Parihar, P.; Mishra, R.K.; Tripathi, D.K.; Singh, V.P.; Chauhan, D.K.; Prasad, S.M. Reactive Oxygen Species (ROS): Beneficial Companions of Plants’ Developmental Processes. Front. Plant Sci. 2016, 7, 1299. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Orenga, S.; Leandro, M.E.D.A.; Tortajada, L.; Marius, N.; Grigore, J.A.; Llorens, P.; Ferrer-Gallego, P.; Laguna, E.; Boscaiu, M.; Vicente, O. Comparative studies on the stress responses of two Bupleurum (Apiaceae) species in support of conservation programmes. Environ. Exp. Bot. 2021, 191, 104616. [Google Scholar] [CrossRef]
- Bose, J.; Rodrigo-Moreno, A.; Shabala, S. ROS homeostasis in halophytes in the context of salinity stress tolerance. J. Exp. Bot. 2014, 65, 1241–1257. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, U.K.; Islam, M.N.; Siddiqui, M.N.; Cao, X.; Khan, M.A.R. Proline, a multifaceted signalling molecule in plant responses to abiotic stress: Understanding the physiological mechanisms. Plant Biol. J. 2022, 24, 227–239. [Google Scholar] [CrossRef]
- Santos-Sánchez, N.F.; Salas-Coronado, R.; Hernández-Carlos, B.; Villanueva-Cañongo, C. Shikimic Acid Pathway in Biosynthesis of Phenolic Compounds. In Plant Physiological Aspects of Phenolic Compounds; IntechOpen: London, UK, 2019; p. 73. [Google Scholar]
- Khanna, K.; Kohli, S.K.; Ohri, P.; Bhardwaj, R.; Al-Huqail, A.A.; Siddiqui, M.H.; Alosaimi, G.S.; Ahmad, P. Microbial Fortification Improved Photosynthetic Efficiency and Secondary Metabolism in Lycopersicon esculentum Plants Under Cd Stress. Biomolecules 2019, 9, 581. [Google Scholar] [CrossRef]
- Siddique, S.; Naveed, M.; Yaseen, M.; Shahbaz, M. Exploring potential of seed endophytic bacteria for enhancing drought stress resilience in maize (Zea mays L.). Sustainability 2022, 14, 673. [Google Scholar] [CrossRef]
- Khan, S.; Malik, A. Toxicity evaluation of textile effluents and role of native soil bacterium in biodegradation of a textile dye. Environ. Sci. Pollut. Res. Int. 2018, 25, 4446–4458. [Google Scholar] [CrossRef]
- Rezvani, M.; Zaefarian, F. Bioaccumulation and translocation factors of cadmium and lead in Aeluropus littoralis. Aust. J. Agri. Eng. 2011, 2, 114–119. [Google Scholar]
- Saeed, Z.; Naveed, M.; Muhammad, I.; Muhammad, A.B.; Annum, S.; Adnan, M.; Azhar, H.; Minggang, X. Combined use of Enterobacter sp. MN17 and zeolite reverts the adverse effects of cadmium on growth, physiology and antioxidant activity of Brassica napus. PLoS ONE 2019, 14, e0213016. [Google Scholar] [CrossRef]
- Asgher, M.; Khan, M.I.R.; Anjum, N.A.; Khan, N.A. Minimising toxicity of cadmium in plants—Role of plant growth regulators. Protoplasma. 2015, 252, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Ismail, M.A.; Amin, M.A.; Eid, A.M.; Hassan, S.E.D.; Mahgoub, H.A.M.; Lashin, I.; Abdelwahab, A.T.; Azab, E.; Gobouri, A.A.; ElKelish, A. Comparative Study between Exogenously Applied Plant Growth Hormones versus Metabolites of Microbial Endophytes as Plant Growth-Promoting for Phaseolus vulgaris L. Cells 2021, 10, 1059. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.; Farooq, M.; Naveed, M.; Nawaz, A.; Shahzad, B. Seed priming of Zn with endophytic bacteria improves the productivity and grain biofortification of bread wheat. Eur. J. Agron. 2018, 94, 98–107. [Google Scholar] [CrossRef]
- Rehman, A.; Farooq, M.; Naveed, M.; Ozturk, L.; Nawaz, A. Pseudomonas-aided zinc application improves the productivity and biofortification of bread wheat. Crop Pasture Sci. 2018, 69, 659–672. [Google Scholar] [CrossRef]






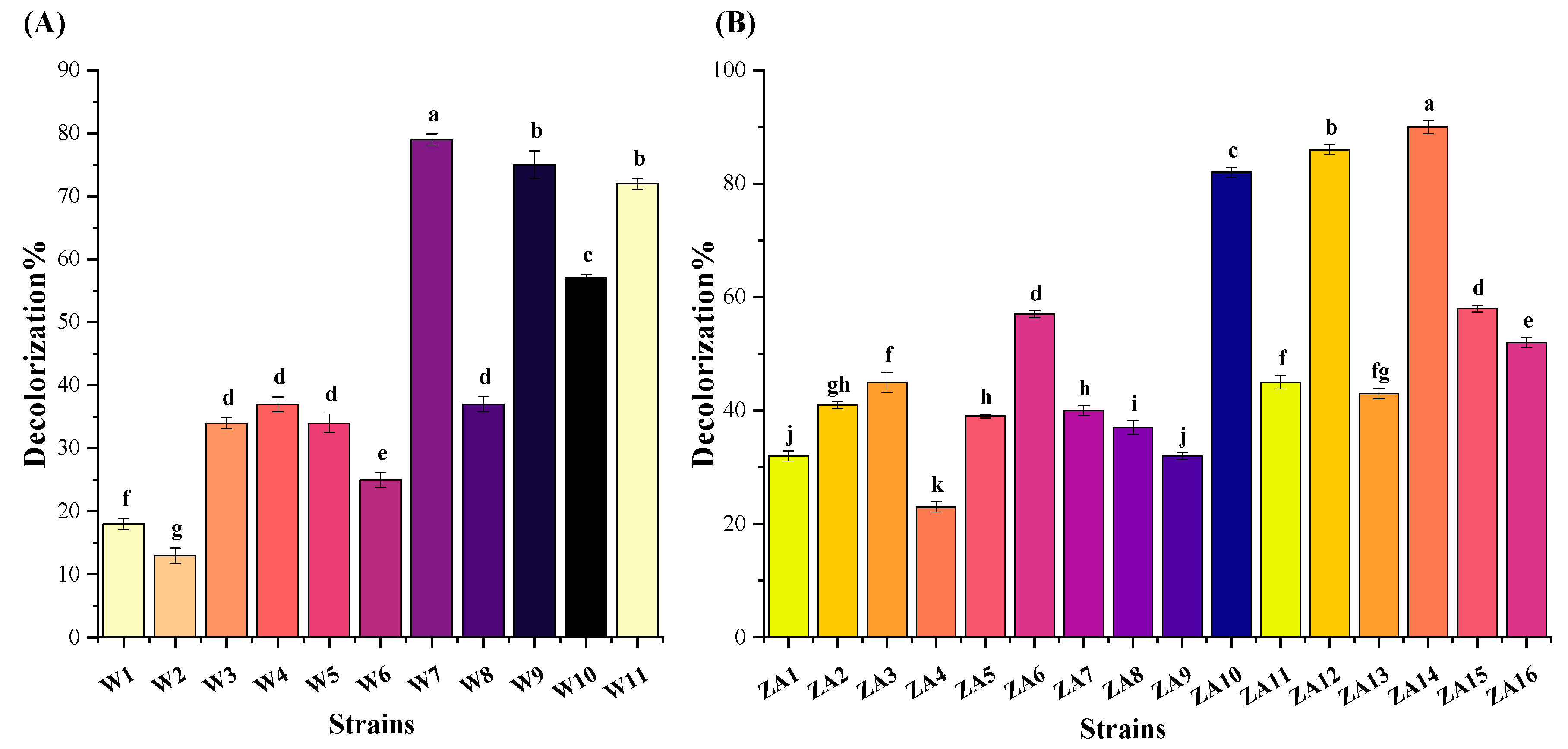{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No. | Characteristics | W7 | W9 | W11 | ZA10 | ZA12 | ZA14 |
|---|---|---|---|---|---|---|---|
| 1. | Gram Reaction | − | − | − | − | − | − |
| 2. | Cellulase | + | + | + | + | + | + |
| 3. | Catalase | + | + | + | + | + | + |
| 4. | N-Fixation | + | + | + | + | + | + |
| 5. | NaCl Tolerance | + | + | + | + | + | + |
| 6. | P-Solubilization | + | + | + | + | + | + |
| 7. | Zn-Solubilization | − | − | − | − | − | − |
| 8. | H2S Production | − | − | − | − | − | − |
| 9. | HCN Production | − | − | − | − | + | − |
| 10. | Amylase | − | + | − | + | + | − |
| 11. | Protease | − | − | − | − | − | − |
| 12. | Ammonia Production | − | + | + | − | − | + |
| 13. | Cellulose | − | − | − | − | − | − |
| 14. | IAA Production | + | + | + | + | + | + |
| Strains | Colony Diameter (cm) | Halozone Diameter (cm) | Solubilization Index | Solubilization Efficiency |
|---|---|---|---|---|
| W7 | 0.8 | 1 | 2.25 | 125 |
| W9 | 0.7 | 1 | 2.43 | 143 |
| W11 | 0.9 | 1.6 | 2.78 | 178 |
| ZA10 | 0.9 | 1.5 | 2.67 | 167 |
| ZA12 | 1 | 1.4 | 2.4 | 140 |
| ZA14 | 0.6 | 2.1 | 4.5 | 350 |
| Strains | SL1 | SL2 | SL3 | SL4 |
|---|---|---|---|---|
| W7 | +++ | ++ | − | − |
| W9 | +++ | ++ | + | − |
| W11 | +++ | ++ | − | − |
| ZA10 | +++ | ++ | + | − |
| ZA12 | +++ | ++ | + | − |
| ZA14 | +++ | +++ | ++ | + |
| Strains | Cd | Cr | Pb | ||||||
|---|---|---|---|---|---|---|---|---|---|
| ML1 | ML2 | ML3 | ML1 | ML2 | ML3 | ML1 | ML2 | ML3 | |
| W7 | +++ | + | − | + | − | − | +++ | ++ | + |
| W9 | ++ | + | − | + | − | − | + | − | − |
| W11 | +++ | ++ | + | + | − | − | ++ | − | − |
| ZA10 | ++ | + | − | + | − | − | + | − | − |
| ZA12 | +++ | ++ | + | ++ | + | − | +++ | ++ | + |
| ZA14 | +++ | ++ | + | +++ | ++ | + | +++ | ++ | + |
| Treatments | Root Length (cm) | Shoot Length (cm) | Fresh Weight (g) | Dry Weight (g) |
|---|---|---|---|---|
| T1 | 15.43 ± 0.15 c | 20.23 ± 0.18 c | 4.17 ± 0.006 c | 1.78 ± 0.02 c |
| T2 | 18.77 ± 0.35 a | 24.03 ± 0.12 a | 4.6 ± 0.01 a | 1.98 ± 0.01 a |
| T3 | 13.27 ± 0.12 d | 17.07 ± 0.09 e | 3.93 ± 0.02 e | 1.69 ± 0.01 d |
| T4 | 16.87 ± 0.20 b | 21.33 ± 0.23 b | 4.38 ± 0.02 b | 1.90 ± 0.01 b |
| T5 | 12.27 ± 0.09 e | 15.30 ± 0.12 f | 3.53 ± 0.03 g | 1.48 ± 0.02 f |
| T6 | 15.73 ± 0.09 c | 19.47 ± 0.09 d | 4.01 ± 0.04 d | 1.70 ± 0.01 d |
| T7 | 10.13 ± 0.18 f | 13.23 ± 0.09 g | 3.29 ± 0.01 h | 1.4 ± 0.01 g |
| T8 | 13.80 ± 0.17 d | 17.36 ± 0.15 e | 3.71 ± 0.01 f | 1.52 ± 0.01 e |
| T9 | 8.80 ± 0.06 g | 11.73 ± 0.09 h | 2.93 ± 0.02 i | 1.24 ± 0.01 i |
| T10 | 12.67 ± 0.26 e | 15.20 ± 0.06 f | 3.32 ± 0.01 h | 1.34 ± 0.01 h |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naseem, Z.; Naveed, M.; Imran, M.; Saqlain, M.; Asif, M.; Bashir, M.; Alamri, S.; Siddiqui, M.H.; Brtnicky, M.; Mustafa, A. Elucidating the Potential of Dye-Degrading Enterobacter cloacae ZA14 for Cultivation of Solanum lycopersicum Plants with Textile Effluents. Water 2023, 15, 3163. https://doi.org/10.3390/w15173163
Naseem Z, Naveed M, Imran M, Saqlain M, Asif M, Bashir M, Alamri S, Siddiqui MH, Brtnicky M, Mustafa A. Elucidating the Potential of Dye-Degrading Enterobacter cloacae ZA14 for Cultivation of Solanum lycopersicum Plants with Textile Effluents. Water. 2023; 15(17):3163. https://doi.org/10.3390/w15173163
Chicago/Turabian StyleNaseem, Zainab, Muhammad Naveed, Muhammad Imran, Muhammad Saqlain, Muhammad Asif, Mohsin Bashir, Saud Alamri, Manzer H. Siddiqui, Martin Brtnicky, and Adnan Mustafa. 2023. "Elucidating the Potential of Dye-Degrading Enterobacter cloacae ZA14 for Cultivation of Solanum lycopersicum Plants with Textile Effluents" Water 15, no. 17: 3163. https://doi.org/10.3390/w15173163
APA StyleNaseem, Z., Naveed, M., Imran, M., Saqlain, M., Asif, M., Bashir, M., Alamri, S., Siddiqui, M. H., Brtnicky, M., & Mustafa, A. (2023). Elucidating the Potential of Dye-Degrading Enterobacter cloacae ZA14 for Cultivation of Solanum lycopersicum Plants with Textile Effluents. Water, 15(17), 3163. https://doi.org/10.3390/w15173163