


Spatial Genetic Structure and Diversity of Large Yellow Croaker (Larimichthys crocea) from the Southern Yellow Sea and North-Central East China Sea: Implications for Conservation and Stock Enhancement

Abstract
:1. Introduction
2. Materials and Methods
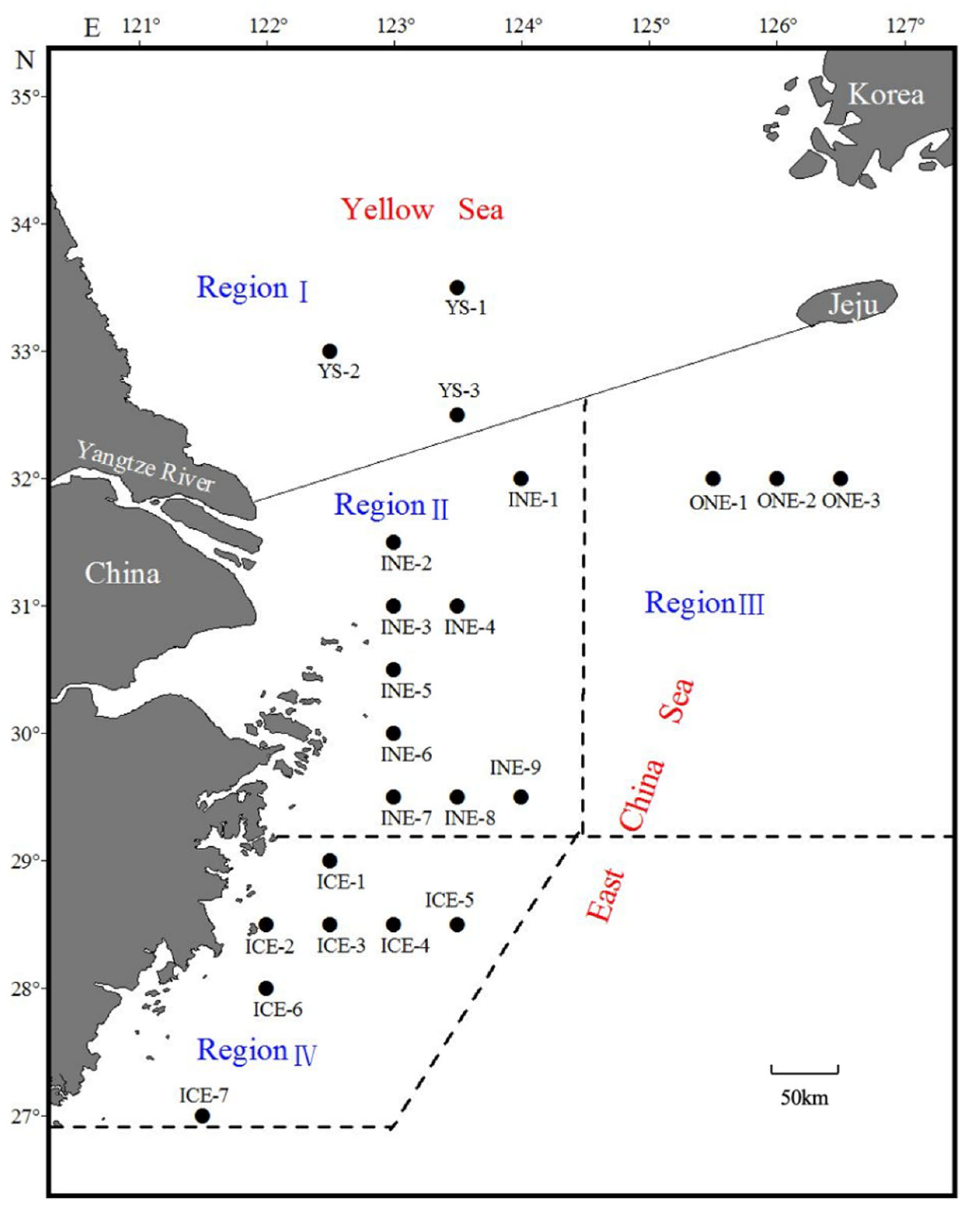
2.1. Sample Collection
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Sequence Alignment and Data Analysis
3. Results
3.1. Characteristics of COI Sequence and Genetic Diversity of the Large Yellow Croaker
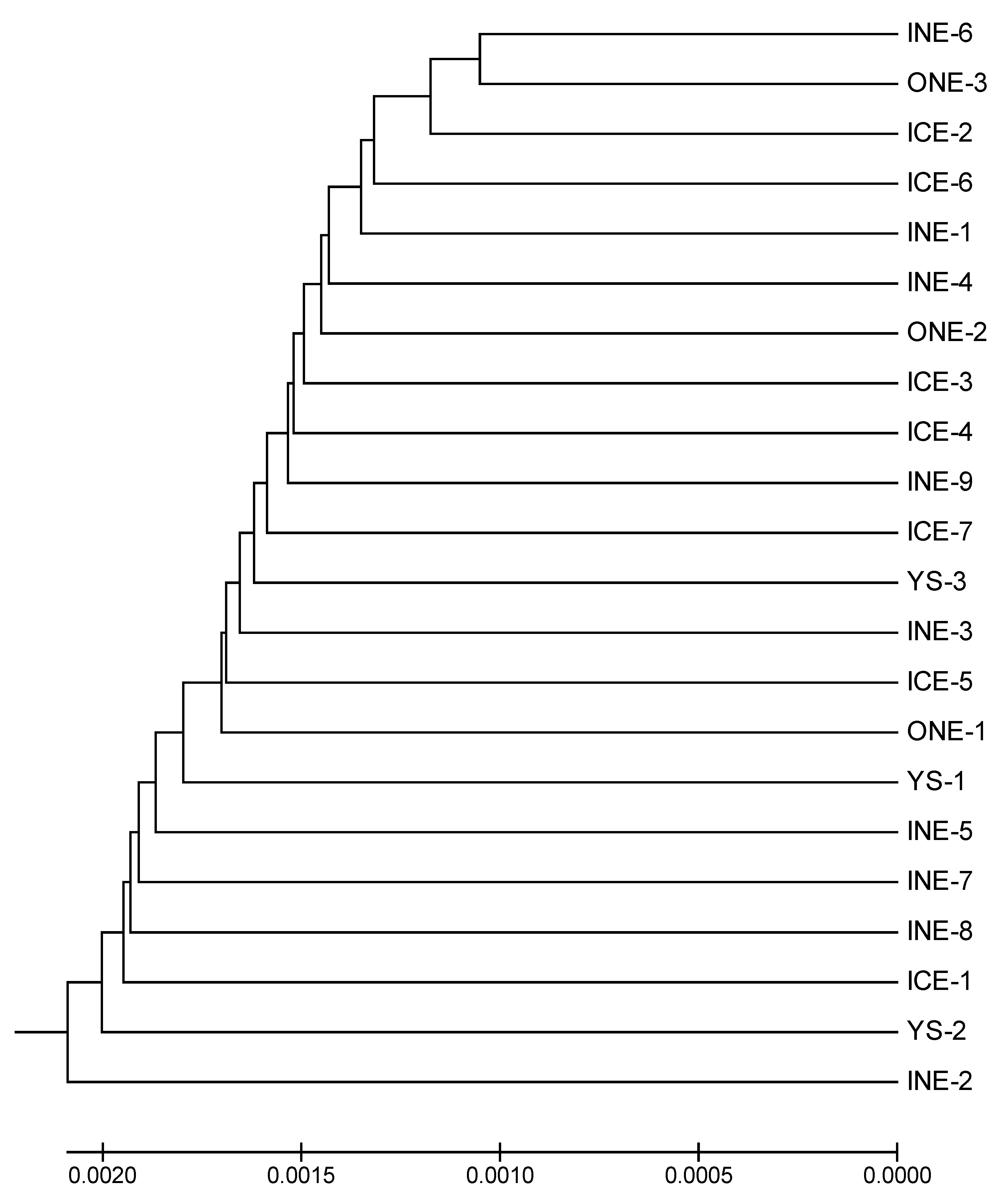
3.2. Genetic Differentiation and Phylogeny of the Large Yellow Croaker
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Su, Y.Q.; Zhang, C.L.; Wang, J. The Culture of Large Yellow Croaker; China Ocean Press: Beijing, China, 2004; pp. 32–36. (In Chinese) [Google Scholar]
- Huang, B.; Walters, C.J. Cohort analysis and population dynamics of large yellow croaker in the China Sea. N. Am. J. Fish. Manag. 1983, 3, 295–305. [Google Scholar] [CrossRef]
- Liu, M.; de Mitcheson, Y.S. Profile of a fishery collapse: Why mariculture failed to save the large yellow croaker. Fish Fish. 2008, 9, 219–242. [Google Scholar] [CrossRef]
- Xu, P.; Ke, Q.; Su, Y.; Liu, J.; Zheng, W. Protection and utilization status and prospect of large yellow croaker (Larimichthys crocea) germplasm resources. J. Fish. China 2022, 46, 674–682. (In Chinese) [Google Scholar]
- Araki, H.; Schmid, C. Is hatchery stocking a help or harm? Evidence, limitations and future directions in ecological and genetic surveys. Aquaculture 2010, 308 (Suppl. S1), S2–S11. [Google Scholar] [CrossRef]
- Lorenzen, K.; Beveridge, M.C.M.; Mangel, M. Cultured fish: Integrative biology and management of domestication and interactions with wild fish. Biol. Rev. 2012, 87, 639–660. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Shi, X.; Su, Y.; Meng, Z.; Lin, H. Loss of genetic diversity in the cultured stocks of the large yellow croaker, Larimichthys crocea, revealed by microsatellites. Int. J. Mol. Sci. 2012, 13, 5584–5597. [Google Scholar] [CrossRef] [Green Version]
- Goodman, D. Selection equilibrium for hatchery and wild spawning fitness in integrated breeding programs. Can. J. Fish. Aquat. Sci. 2005, 62, 374–389. [Google Scholar] [CrossRef]
- Araki, H.; Cooper, B.; Blouin, M.S. Genetic effects of captive breeding cause a rapid, cumulative fitness decline in the wild. Science 2007, 318, 100–103. [Google Scholar] [CrossRef] [Green Version]
- Kitada, S. Economic, ecological and genetic impacts of marine stock enhancement and sea ranching: A systematic review. Fish Fish. 2018, 19, 511–532. [Google Scholar] [CrossRef] [Green Version]
- Cross, T.F. Genetic implications of translocation and stocking of fish species, with particular reference to Western Australia. Aquac. Res. 2000, 31, 83–94. [Google Scholar] [CrossRef]
- Ward, R.D. The importance of identifying spatial population structure in restocking and stock enhancement programmes. Fish. Res. 2006, 80, 9–18. [Google Scholar] [CrossRef]
- Lü, H.; Xu, J.; Haegen, G.V. Supplementing marine capture fisheries in the East China Sea: Sea ranching of Prawn Penaeus orientalis, restocking of large yellow croaker Pseudosciaena crocea, and cage culture. Rev. Fish. Sci. 2008, 16, 366–376. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Wang, Y.L.; Lin, L.M.; Qiu, S.Z.; Ben, X.M. Genetic polymorphisms in wild and cultured large yellow croaker Pseudosciaena crocea using AFLP fingerprinting. J. Fish. Sci. China 2002, 9, 198–202. (In Chinese) [Google Scholar]
- Chen, J.; Li, Z.B.; Fang, X.; Lei, G.G.; Zhang, G.L.; Zhao, B.L.; Wang, Z.L. The genetic structure of wild and cultivated populations of Pseudosciaena crocea. Mar. Sci. 2010, 34, 45–48. [Google Scholar]
- Chen, W.; Zhang, F.; Wang, J.; Wei, H.; Jiang, Y.; Zhang, H.; Ling, J.; Cheng, J.; Ma, L. Genetic diversity of wild and cultured populations of Larimichthys crocea in the East China Sea and Yellow Sea based on COI sequence. J. Fish. Sci. China 2016, 23, 1255–1267. (In Chinese) [Google Scholar]
- Liu, B.Q.; Dong, W.Q.; Wang, Y.J.; Zhu, S.H.; Wu, W.X. Identification of germ plasm in Pseudosciaena crocea Tai-chu race by AFLP. Acta Hydrob. Sin. 2005, 29, 413–416. [Google Scholar]
- Chen, S.Y.; Xu, S.X.; Zhang, Z.Y.; Guo, Z.R.; Sun, Z.X.; Zhu, L.J. Study of genetic diversity of wild and culture populations of Pseudosciaena crocea using two molecular markers. Mar. Sci. 2011, 35, 82–87. (In Chinese) [Google Scholar]
- Yuan, J.; Lin, H.; Wu, L.; Zhuang, X.; Ma, J.; Kang, B.; Ding, S. Resource status and effect of long-term stock enhancement of large yellow croaker in China. Front. Mar. Sci. 2021, 8, 743836. [Google Scholar] [CrossRef]
- Jiang, L.; Chen, Y.; Zhang, J.; Zhu, A.; Wu, C.; Liu, L.; Lin, Z.; Dong, Y. Population structure of large yellow croaker (Larimichthys crocea) revealed by single nucleotide polymorphisms. Biochem. Syst. Ecol. 2015, 63, 136–142. [Google Scholar] [CrossRef]
- Petit-Marty, N.; Vázquez-Luis, M.; Hendriks, I.E. Use of the nucleotide diversity in COI mitochondrial gene as an early diagnostic of conservation status of animal species. Conserv. Lett. 2021, 14, e12756. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987; 512p. [Google Scholar]
- Rozas, J.; Sanchez-Delbarrio, J.C.; Messeguer, X.; Rozas, R. DnaSP, DNA polymorphism analyses by coalescent and other methods. Bioinformatics 2003, 19, 2496–2497. [Google Scholar] [CrossRef] [Green Version]
- Polzin, T.; Daneshmand, S.V. On Steiner trees and minimum spanning trees in hypergraphs. Oper. Res. Lett. 2003, 31, 12–20. [Google Scholar] [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. Online 2005, 1, 47–50. [Google Scholar] [CrossRef] [Green Version]
- Rice, W.R. Analyzing tables of statistical tests. Evolution 1989, 43, 223–225. [Google Scholar] [CrossRef]
- Lin, X.Z. Biological characteristics and resources status of three main commercial fishes in offshore waters of China. J. Fish. China 1987, 11, 187–194. (In Chinese) [Google Scholar]
- Zhao, S.L.; Wang, R.X.; Liu, X.S. Reasons of exhaustion of resources of Pseudosciaena crocea in Zhoushan fishing ground and the measures of protection and proliferation. J. Zhejiang Ocean. Univ. (Nat. Sci.) 2002, 21, 160–165. (In Chinese) [Google Scholar]
- Xu, H.Y.; Zhou, Y.D. A preliminary study on release and enhancement of large yellow croaker Pseudosciaena crocea in the north coast of Zhejiang. Mar. Fish. 2003, 25, 69–72. (In Chinese) [Google Scholar]
- Wang, L.; Shi, X.; Su, Y.; Meng, Z.; Lin, H. Genetic divergence and historical demography in the endangered large yellow croaker revealed by mtDNA. Biochem. Syst. Ecol. 2013, 46, 137–144. [Google Scholar] [CrossRef]
- Kon, T.; Pei, L.; Ichikawa, R.; Chen, C.; Wang, P.; Takemura, I.; Ye, Y.; Yan, X.; Guo, B.; Li, W.; et al. Whole-genome resequencing of large yellow croaker (Larimichthys crocea) reveals the population structure and signatures of environmental adaptation. Sci. Rep. 2021, 11, 11235. [Google Scholar] [CrossRef]
- Li, M.; Miao, L.; Chen, J.; Shi, Y.; Lu, X. Division of populations for Pseudosciaena crocea based on population ecology concept: Discussion. J. Ningbo Univ. (NSEE) 2013, 26, 1–5. (In Chinese) [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Sea Area | Sites | No. of Samples | No. of Polymorphic Sites | No. of Haplotypes | Haplotype Diversity, h | Nucleotide Diversity, π |
|---|---|---|---|---|---|---|
| Region I: southern Yellow Sea (YS) | YS-1 | 43 | 19 | 17 | 0.907 | 0.00443 |
| YS-2 | 35 | 19 | 15 | 0.892 | 0.00443 | |
| YS-3 | 19 | 13 | 10 | 0.895 | 0.00358 | |
| Region II: inshore region of northern East China Sea (INE) | INE-1 | 26 | 13 | 10 | 0.738 | 0.00298 |
| INE-2 | 33 | 16 | 15 | 0.917 | 0.00473 | |
| INE-3 | 26 | 13 | 10 | 0.895 | 0.00358 | |
| INE-4 | 31 | 14 | 13 | 0.892 | 0.00309 | |
| INE-5 | 91 | 28 | 27 | 0.858 | 0.00407 | |
| INE-6 | 20 | 9 | 9 | 0.795 | 0.00225 | |
| INE-7 | 32 | 22 | 18 | 0.993 | 0.00437 | |
| INE-8 | 36 | 23 | 18 | 0.916 | 0.00440 | |
| INE-9 | 35 | 16 | 14 | 0.832 | 0.00327 | |
| Region III: offshore region of northern East China Sea (ONE) | ONE-1 | 28 | 14 | 11 | 0.839 | 0.00367 |
| ONE-2 | 5 | 4 | 5 | 1.000 | 0.00308 | |
| ONE-3 | 7 | 4 | 5 | 0.857 | 0.00220 | |
| Region IV: inshore region of central East China Sea (ICE) | ICE-1 | 35 | 25 | 21 | 0.948 | 0.00444 |
| ICE-2 | 9 | 4 | 5 | 0.861 | 0.00256 | |
| ICE-3 | 69 | 28 | 25 | 0.852 | 0.00325 | |
| ICE-4 | 36 | 20 | 17 | 0.875 | 0.00335 | |
| ICE-5 | 22 | 11 | 10 | 0.866 | 0.00367 | |
| ICE-6 | 10 | 5 | 4 | 0.644 | 0.00282 | |
| ICE-7 | 14 | 9 | 6 | 0.813 | 0.00338 | |
| Total | 662 | 0.872 | 0.00373 |
| Locality | YS-1 | YS-2 | YS-3 | INE-1 | INE-2 | INE-3 | INE-4 | INE-5 | INE-6 | INE-7 | INE-8 | INE-9 | ICE-1 | ICE-2 | ICE-3 | ICE-4 | ICE-5 | ICE-6 | ICE-7 | ONE-1 | ONE-2 | ONE-3 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| YS-1 | 0.0041 | −0.0016 | −0.0110 | −0.0088 | 0.0051 | −0.0071 | 0.0101 | 0.0089 | −0.0111 | −0.0095 | −0.0082 | −0.0061 | −0.0141 | 0.0052 | 0.0023 | −0.0061 | −0.0128 | −0.0325 | 0.0062 | −0.0003 | −0.0051 | −0.0323 |
| YS-2 | 0.0043 | 0.0045 | 0.0301 | 0.0110 | 0.0031 | 0.0085 | 0.0417 | 0.0017 | 0.0245 | −0.0018 | 0.0022 | 0.0004 | 0.0033 | 0.0469 | 0.0264 | 0.0206 | −0.0075 | −0.0462 | 0.0386 | 0.0053 | 0.0035 | −0.0139 |
| YS-3 | 0.0038 | 0.0042 | 0.0036 | −0.0166 | 0.0125 | 0.0015 | 0.0024 | 0.0265 | −0.0124 | −0.0001 | −0.0069 | 0.0147 | −0.0148 | −0.0421 | −0.0061 | −0.0231 | −0.0112 | −0.0056 | −0.0286 | −0.0013 | 0.0269 | −0.0525 |
| INE-1 | 0.0035 | 0.0038 | 0.0032 | 0.0030 | −0.0025 | −0.0108 | 0.0036 | 0.0057 | −0.0075 | −0.0103 | −0.0165 | −0.0121 | −0.0200 | 0.0114 | −0.0141 | −0.0158 | −0.0102 | −0.0220 | −0.0007 | −0.0145 | 0.0240 | −0.0462 |
| INE-2 | 0.0044 | 0.0046 | 0.0042 | 0.0039 | 0.0048 | 0.0191 | 0.0538 | −0.0099 | 0.0378 | 0.0116 | 0.0071 | 0.0123 | 0.0032 | 0.0342 | 0.0242 | 0.0195 | −0.0180 | −0.0511 | 0.0079 | −0.0125 | 0.0310 | 0.0000 |
| INE-3 | 0.0038 | 0.0041 | 0.0036 | 0.0033 | 0.0043 | 0.0036 | 0.0082 | 0.0128 | −0.0129 | −0.0149 | −0.0136 | −0.0180 | −0.0185 | 0.0319 | 0.0081 | 0.0008 | 0.0069 | −0.0158 | 0.0373 | 0.0002 | −0.0101 | −0.0291 |
| INE-4 | 0.0036 | 0.0039 | 0.0033 | 0.0031 | 0.0042 | 0.0034 | 0.0031 | 0.0486 | −0.0160 | 0.0062 | 0.0027 | 0.0109 | 0.0007 | 0.0049 | −0.0002 | −0.0087 | 0.0299 | 0.0328 | 0.0187 | 0.0266 | −0.0349 | −0.0499 |
| INE-5 | 0.0041 | 0.0043 | 0.0040 | 0.0036 | 0.0044 | 0.0039 | 0.0038 | 0.0041 | 0.0300 | 0.0131 | 0.0133 | 0.0047 | 0.0073 | 0.0553 | 0.0246 | 0.0222 | −0.0074 | −0.0495 | 0.0297 | −0.0101 | 0.0189 | 0.0069 |
| INE-6 | 0.0031 | 0.0035 | 0.0029 | 0.0026 | 0.0037 | 0.0029 | 0.0026 | 0.0033 | 0.0023 | −0.0043 | −0.0076 | −0.0079 | −0.0152 | 0.0170 | −0.0114 | −0.0204 | 0.0147 | 0.0233 | 0.0201 | 0.0086 | −0.0085 | −0.0380 |
| INE-7 | 0.0042 | 0.0044 | 0.0040 | 0.0037 | 0.0046 | 0.0039 | 0.0038 | 0.0043 | 0.0033 | 0.0044 | −0.0088 | −0.0142 | −0.0136 | 0.0034 | 0.0039 | −0.0007 | −0.0060 | −0.0324 | 0.0093 | 0.0072 | −0.0142 | −0.0417 |
| INE-8 | 0.0042 | 0.0045 | 0.0040 | 0.0037 | 0.0046 | 0.0040 | 0.0038 | 0.0043 | 0.0033 | 0.0044 | 0.0044 | −0.0096 | −0.0125 | 0.0069 | 0.0025 | −0.0029 | −0.0082 | −0.0258 | 0.0107 | −0.0001 | −0.0156 | −0.0524 |
| INE-9 | 0.0037 | 0.0039 | 0.0035 | 0.0031 | 0.0041 | 0.0034 | 0.0032 | 0.0037 | 0.0028 | 0.0038 | 0.0038 | 0.0033 | −0.0179 | 0.0503 | 0.0018 | 0.0030 | 0.0033 | −0.0236 | 0.0401 | −0.0036 | −0.0185 | −0.0242 |
| ICE-1 | 0.0042 | 0.0045 | 0.0040 | 0.0037 | 0.0046 | 0.0040 | 0.0038 | 0.0043 | 0.0033 | 0.0044 | 0.0044 | 0.0038 | 0.0045 | 0.0064 | −0.0070 | −0.0113 | −0.0071 | −0.0269 | 0.0052 | −0.0090 | −0.0289 | −0.0390 |
| ICE-2 | 0.0034 | 0.0038 | 0.0030 | 0.0028 | 0.0039 | 0.0032 | 0.0029 | 0.0036 | 0.0024 | 0.0036 | 0.0036 | 0.0031 | 0.0036 | 0.0026 | 0.0008 | −0.0203 | 0.0004 | 0.0448 | −0.0517 | 0.0283 | 0.0802 | −0.0424 |
| ICE-3 | 0.0037 | 0.0039 | 0.0034 | 0.0031 | 0.0041 | 0.0034 | 0.0032 | 0.0038 | 0.0027 | 0.0038 | 0.0038 | 0.0033 | 0.0038 | 0.0030 | 0.0033 | −0.0120 | −0.0002 | −0.0099 | −0.0006 | 0.0016 | −0.0056 | −0.0420 |
| ICE-4 | 0.0037 | 0.0040 | 0.0034 | 0.0031 | 0.0041 | 0.0035 | 0.0032 | 0.0038 | 0.0028 | 0.0039 | 0.0039 | 0.0033 | 0.0039 | 0.0029 | 0.0033 | 0.0034 | −0.0052 | −0.0118 | −0.0158 | −0.0056 | −0.0064 | −0.0433 |
| ICE-5 | 0.0038 | 0.0040 | 0.0036 | 0.0033 | 0.0042 | 0.0037 | 0.0035 | 0.0039 | 0.0030 | 0.0040 | 0.0040 | 0.0035 | 0.0041 | 0.0032 | 0.0035 | 0.0035 | 0.0037 | −0.0599 | −0.0167 | −0.0177 | 0.0343 | −0.0306 |
| ICE-6 | 0.0034 | 0.0035 | 0.0032 | 0.0029 | 0.0037 | 0.0032 | 0.0031 | 0.0033 | 0.0026 | 0.0035 | 0.0036 | 0.0030 | 0.0036 | 0.0028 | 0.0030 | 0.0031 | 0.0031 | 0.0028 | −0.0015 | −0.0424 | 0.0467 | 0.0016 |
| ICE-7 | 0.0038 | 0.0041 | 0.0034 | 0.0032 | 0.0041 | 0.0036 | 0.0033 | 0.0039 | 0.0029 | 0.0039 | 0.0040 | 0.0035 | 0.0040 | 0.0029 | 0.0033 | 0.0033 | 0.0035 | 0.0031 | 0.0034 | 0.0034 | 0.0452 | −0.0297 |
| ONE-1 | 0.0039 | 0.0041 | 0.0036 | 0.0033 | 0.0042 | 0.0036 | 0.0035 | 0.0039 | 0.0030 | 0.0041 | 0.0041 | 0.0035 | 0.0040 | 0.0033 | 0.0035 | 0.0035 | 0.0036 | 0.0031 | 0.0036 | 0.0037 | 0.0070 | −0.0182 |
| ONE-2 | 0.0036 | 0.0039 | 0.0035 | 0.0031 | 0.0042 | 0.0033 | 0.0030 | 0.0038 | 0.0026 | 0.0038 | 0.0038 | 0.0031 | 0.0038 | 0.0030 | 0.0032 | 0.0032 | 0.0036 | 0.0031 | 0.0034 | 0.0035 | 0.0031 | 0.0107 |
| ONE-3 | 0.0031 | 0.0034 | 0.0028 | 0.0025 | 0.0036 | 0.0029 | 0.0026 | 0.0033 | 0.0021 | 0.0033 | 0.0032 | 0.0027 | 0.0033 | 0.0023 | 0.0027 | 0.0027 | 0.0029 | 0.0025 | 0.0028 | 0.0030 | 0.0026 | 0.0022 |
| Source of Variation | Sum of Squares | Variance Components | Percentage of Vatiation (%) | p Value |
|---|---|---|---|---|
| Among groups | 3.704 | 0.00150 | 0.15419 | 0.39003 |
| Among populations within groups | 18.250 | 0.00160 | 0.16444 | 0.24829 |
| Within populations | 619.574 | 0.96808 | 99.68137 | 0.16031 |
| Total | 641.527 | 0.97118 | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, F.; Jiang, Y.; Ma, C.; Chen, W.; Cheng, J.; Ma, L. Spatial Genetic Structure and Diversity of Large Yellow Croaker (Larimichthys crocea) from the Southern Yellow Sea and North-Central East China Sea: Implications for Conservation and Stock Enhancement. Water 2023, 15, 338. https://doi.org/10.3390/w15020338
Zhang F, Jiang Y, Ma C, Chen W, Cheng J, Ma L. Spatial Genetic Structure and Diversity of Large Yellow Croaker (Larimichthys crocea) from the Southern Yellow Sea and North-Central East China Sea: Implications for Conservation and Stock Enhancement. Water. 2023; 15(2):338. https://doi.org/10.3390/w15020338
Chicago/Turabian StyleZhang, Fengying, Yazhou Jiang, Chunyan Ma, Wei Chen, Jiahua Cheng, and Lingbo Ma. 2023. "Spatial Genetic Structure and Diversity of Large Yellow Croaker (Larimichthys crocea) from the Southern Yellow Sea and North-Central East China Sea: Implications for Conservation and Stock Enhancement" Water 15, no. 2: 338. https://doi.org/10.3390/w15020338
APA StyleZhang, F., Jiang, Y., Ma, C., Chen, W., Cheng, J., & Ma, L. (2023). Spatial Genetic Structure and Diversity of Large Yellow Croaker (Larimichthys crocea) from the Southern Yellow Sea and North-Central East China Sea: Implications for Conservation and Stock Enhancement. Water, 15(2), 338. https://doi.org/10.3390/w15020338