
Interannual Variation of Ichthyoplankton Community Structure in the Yellow River Estuary, China

Abstract
:1. Introduction
2. Materials and Methods
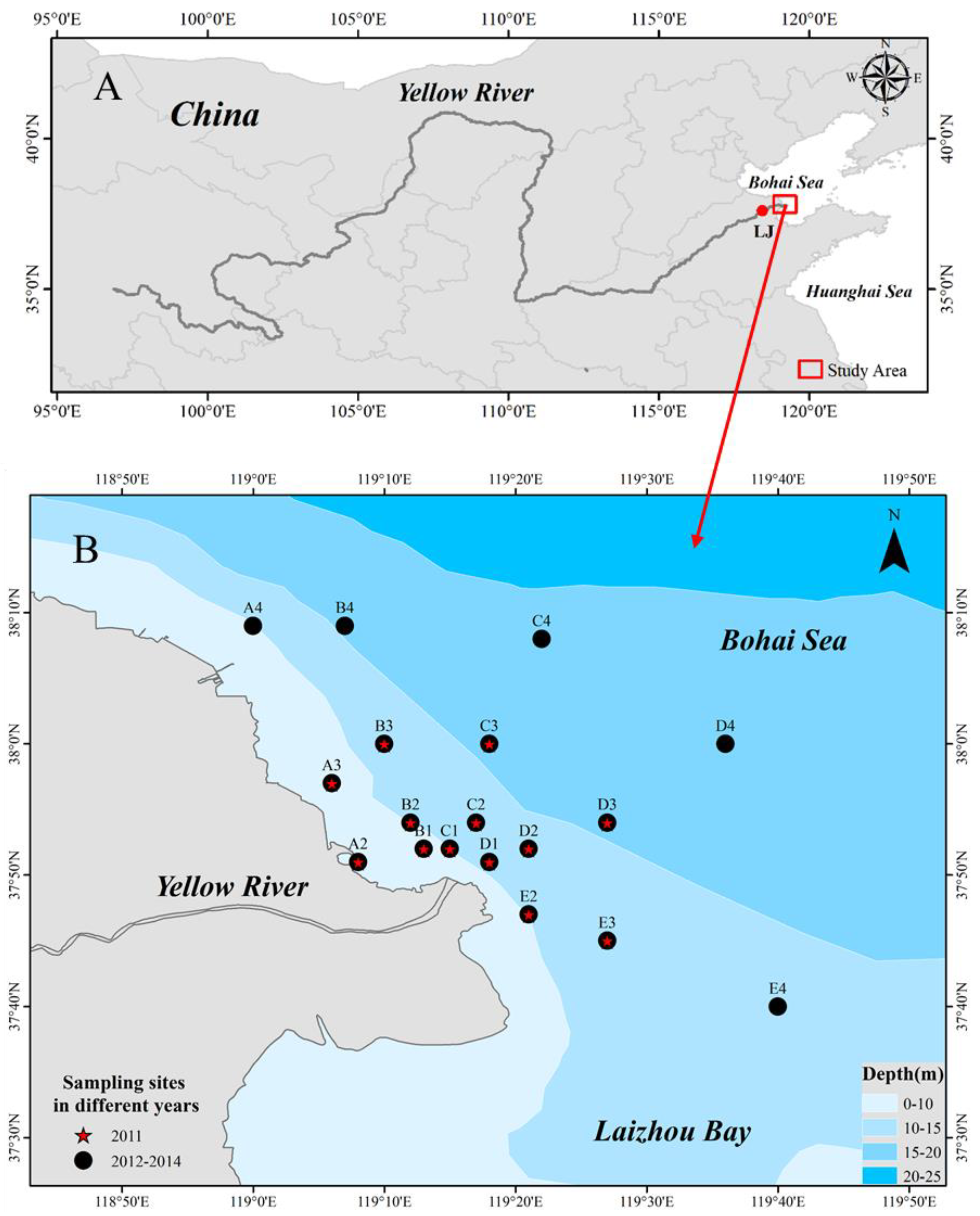
2.1. Survey Area and Stations
2.2. Sample Collection, Processing, and Analysis
2.3. Statistical Analyses
3. Results
3.1. Annual Change in Environmental Factors
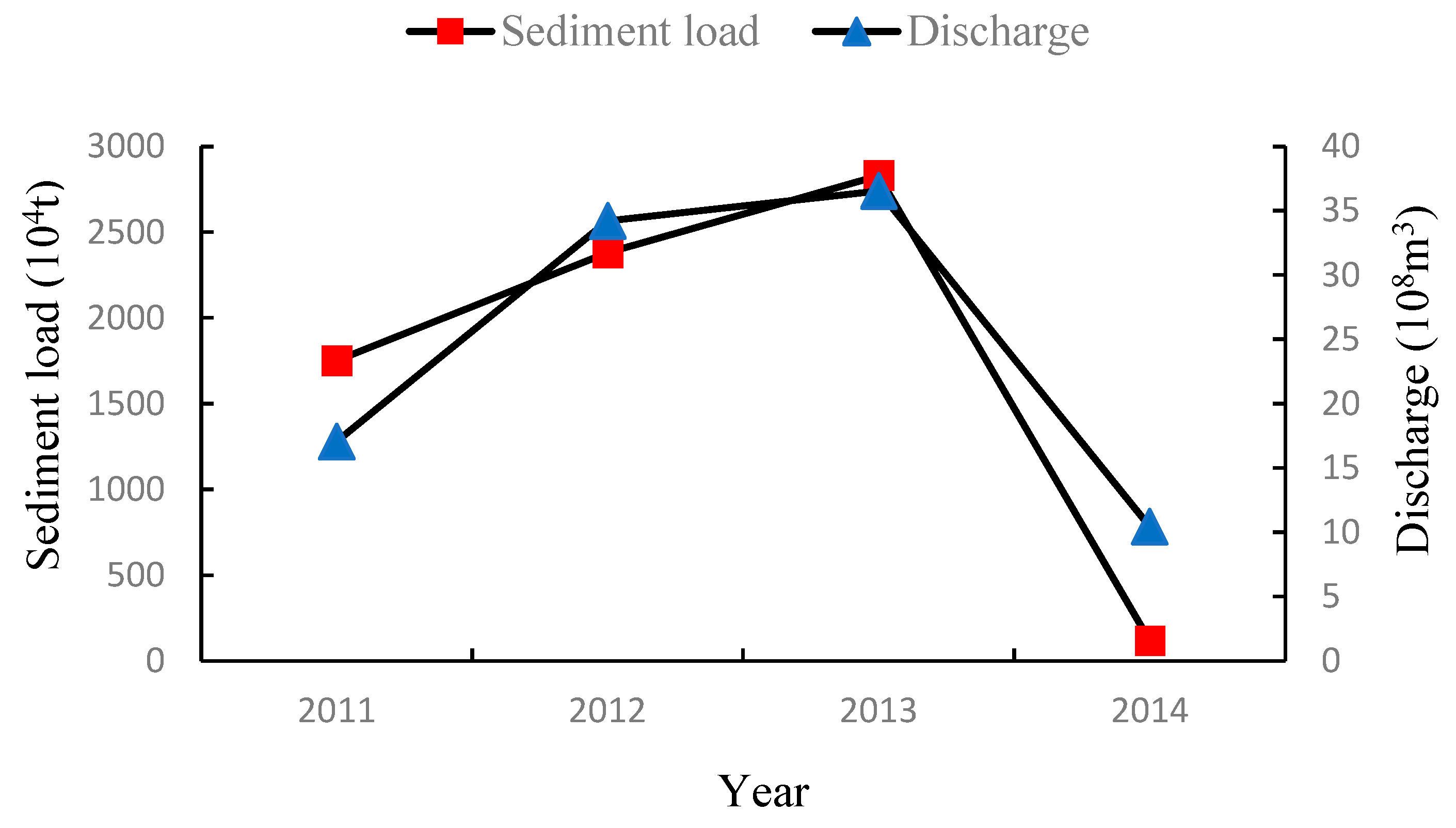
3.1.1. Interannual Variation of Water and Sediment Flux into the Sea of the Yellow River
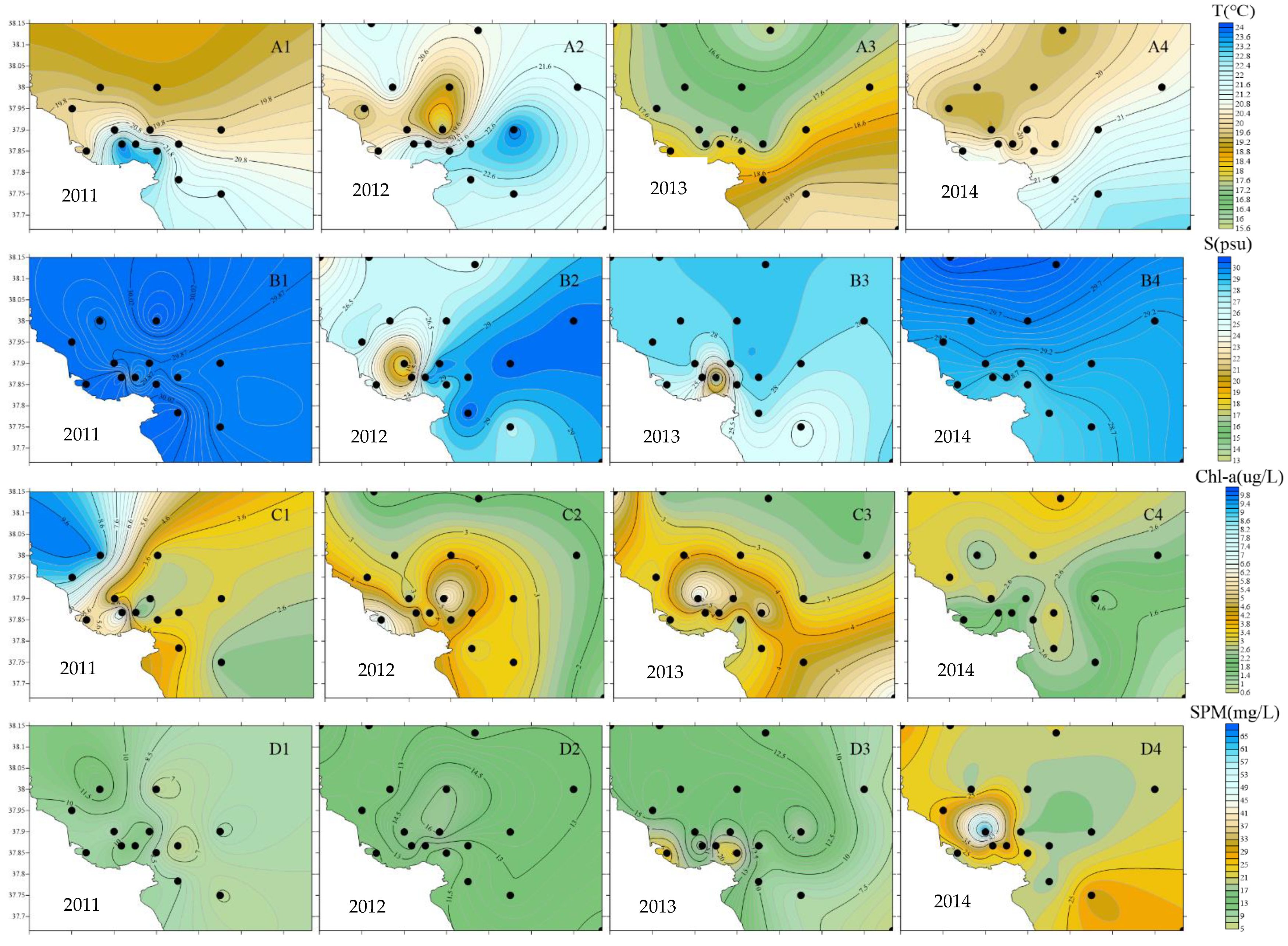
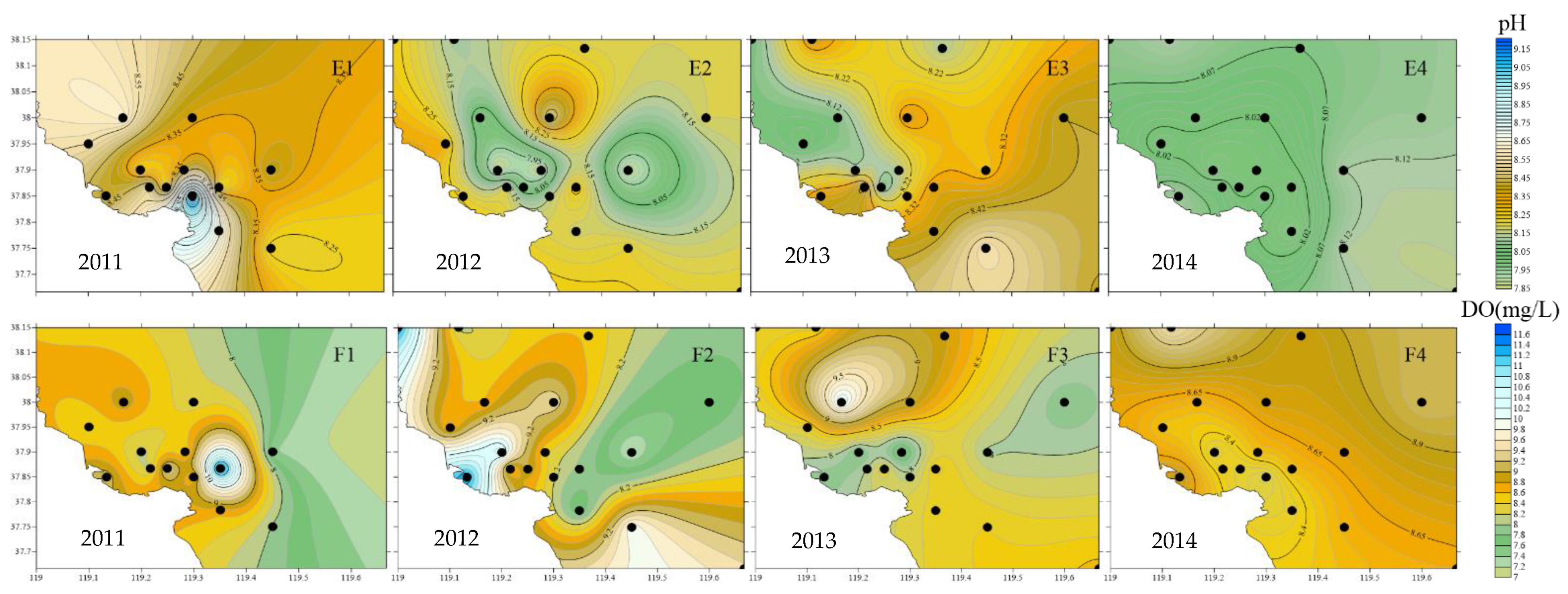
3.1.2. Analysis of Environmental Parameter
3.2. Species Composition and Interannual Variation of Eggs and Larvae
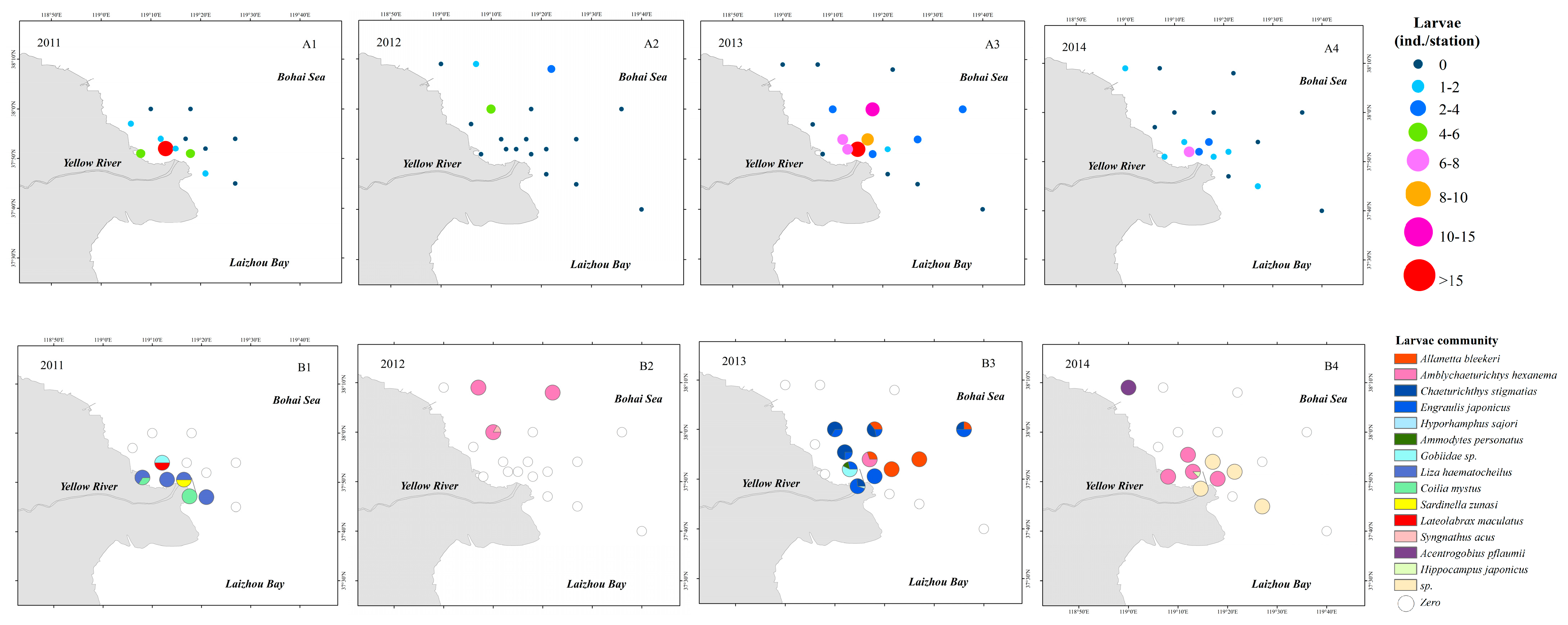
3.3. Spatial Distribution of Eggs and Larvae
3.4. Community Structure Analysis
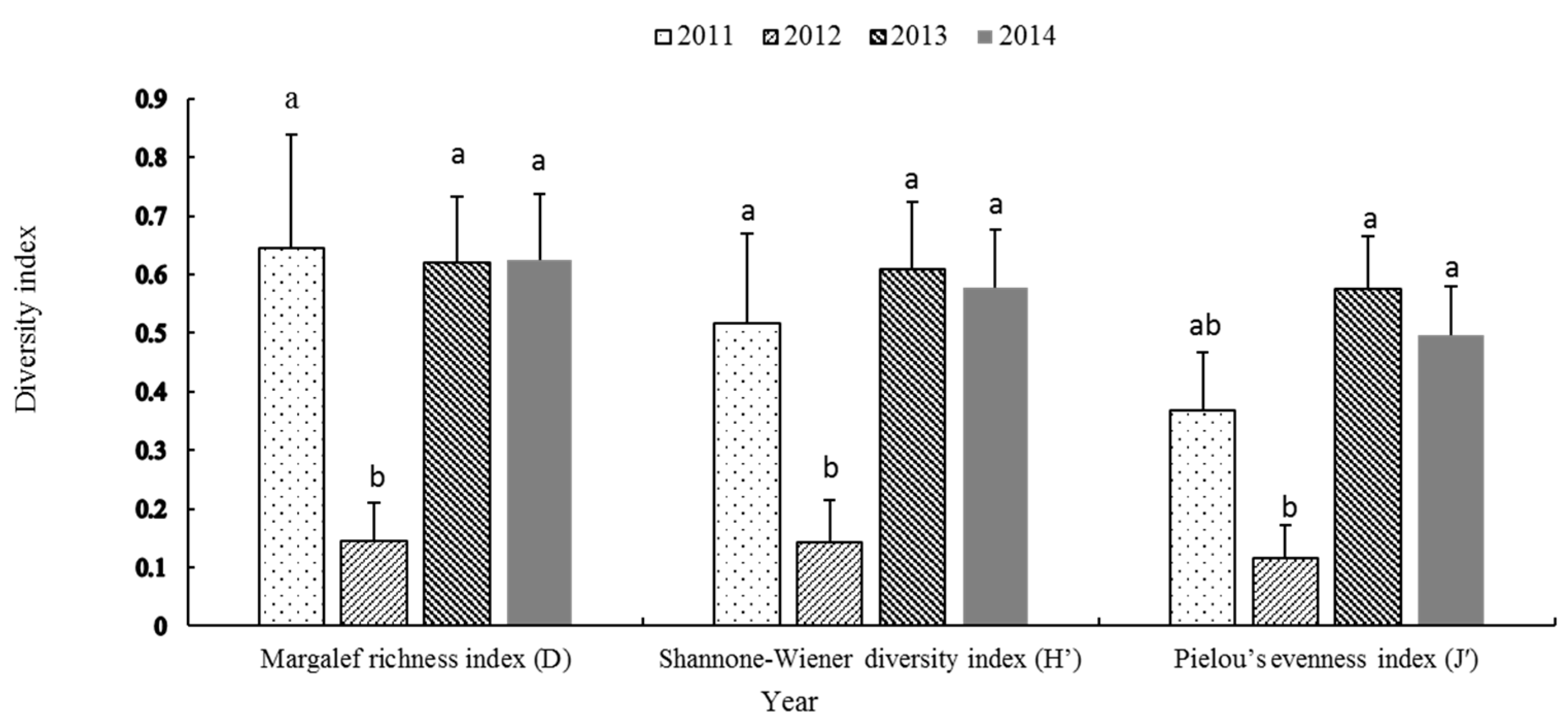
3.5. Biodiversity Levels
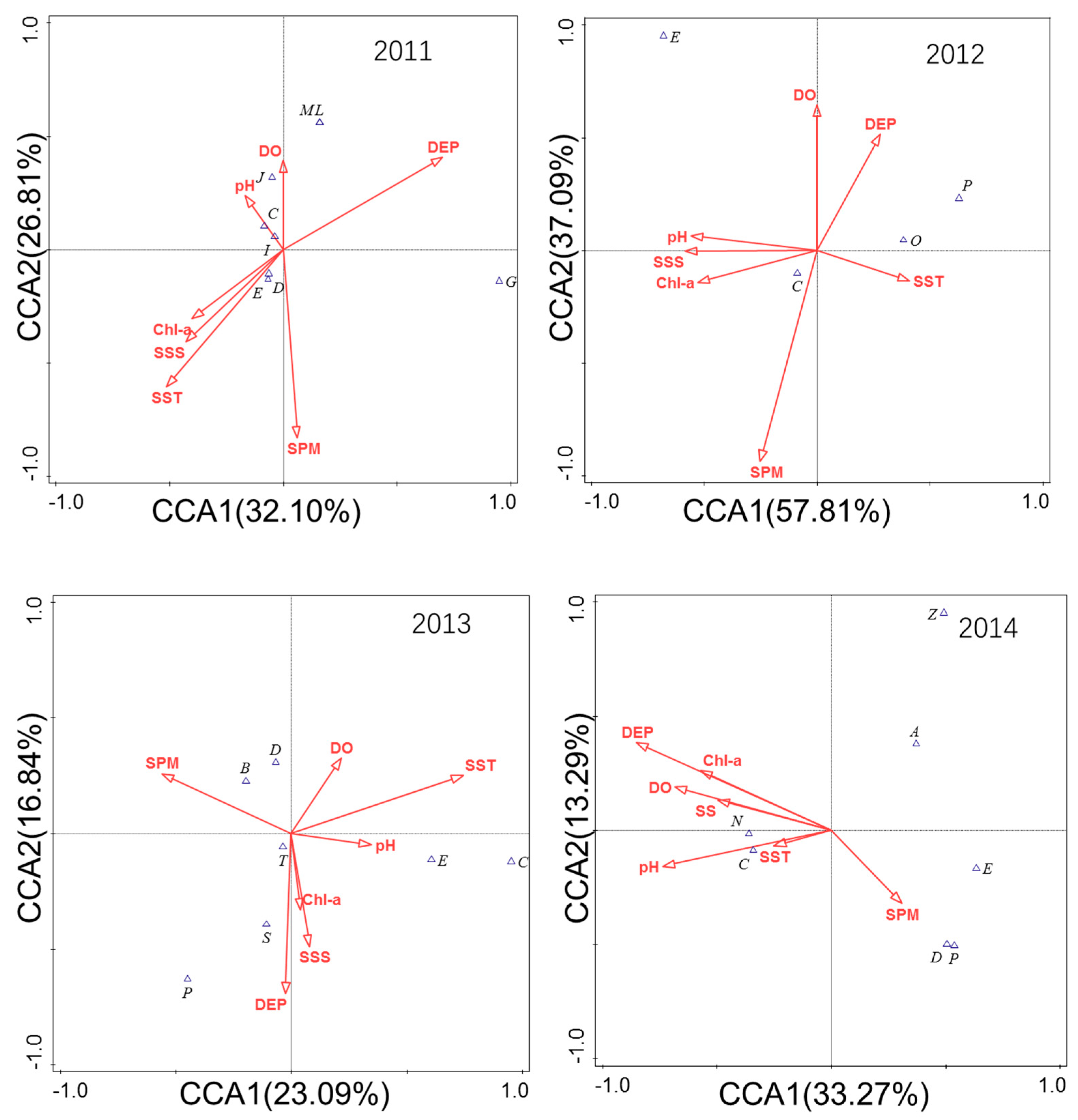
3.6. Analysis of the Relationship between Community Structure and Environmental Factors
4. Discussion
4.1. Species Composition and Population Distributions
4.2. Interannual Variation of Community Diversity
4.3. Community Structure Changes
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhuang, P.; Wang, Y.H.; Li, S.F.; Deng, S.M.; Li, C.S.; Ni, Y. Fishes of the Yangtze Estuary; Shanghai Scientific and Technical Publishers: Shanghai, China, 2006. [Google Scholar]
- Wingard, G.L.; Lorenz, J.J. Integrated conceptual ecological model and habitat indices for the southwest Florida coastal wetlands. Ecol. Ind. 2014, 44, 92–107. [Google Scholar] [CrossRef]
- Luo, B.Z. Estuarine Ecology and Environment before Impoundment of the Three Gorges Project; Ocean Press: Beijing, China, 2015. [Google Scholar]
- Strokal, M.; Kroeze, C.; Li, L.L.; Luan, S.J.; Wang, H.Z.; Yang, S.S.; Zhang, Y.S. Increasing dissolved nitrogen and phosphorus export by the Pearl River (Zhujiang): A modeling approach at the sub-basin scale to assess effective nutrient management. Biogeochemistry 2015, 125, 221–242. [Google Scholar] [CrossRef] [Green Version]
- Roubeix, V.; Rousseau, V.; Lancelot, C. Diatom succession and silicon removal from freshwater in estuarine mixing zones: From experiment to modelling. Estuar. Coast Shelf Sci. 2008, 78, 14–26. [Google Scholar] [CrossRef]
- Shan, X.J.; Sun, P.F.; Jin, X.S.; Li, X.S.; Dai, F.Q. Long-term changes in fish assemblage structure in the Yellow River Estuary ecosystem. Mar. Coast. Fish. 2013, 5, 65–78. [Google Scholar] [CrossRef]
- Berasategui, A.D.; Acha, E.M.; Araoz, F.N.C. Spatial patterns of ichthyoplankton assemblages in the Río de la Plata Estuary (Argentina–Uruguay). Estuar. Coast. Shelf Sci. 2004, 60, 599–610. [Google Scholar] [CrossRef]
- Hou, G.; Wang, J.R.; Liu, L.M.; Chen, Y.Y.; Pan, C.H.; Li, J.B.; Zhang, H. Assemblage Structure of the ichthyoplankton and its Relationship with environmental factors in spring and autumn off the Pearl River estuary. Front. Mar. Sci. 2021, 8, 732970. [Google Scholar] [CrossRef]
- Wan, R.J.; Zhou, F.; Shan, X.J.; Sun, S. Impacts of variability of habitat factors on species composition of ichthyoplankton and distribution of fish spawning ground in the Changjiang River estuary and its adjacent waters. Acta Ecol. Sin. 2010, 30, 155–165. [Google Scholar]
- Hernandez, F.J., Jr.; Powers, S.P.; Graham, W.M. Seasonal variability in ichthyoplankton abundance and assemblage composition in the northern Gulf of Mexico off Alabama. Fish. Bull. 2010, 108, 192–206. [Google Scholar]
- Bertone, E.; Sahin, O.; Richards, R.; Roiko, A. Extreme events, water quality and health a participatory Bayesian risk assessment tool for managers of reservoirs. J. Clean. Prod. 2016, 135, 657–667. [Google Scholar] [CrossRef] [Green Version]
- Chambers, R.C. Environmental influences on egg and propagule sizes in marine fishes. In Early Life History and Recruitment in Fish Populations; Chambers, R.C., Trippel, E.A., Eds.; Chapman & Hall: London, UK, 1997. [Google Scholar]
- Bian, X.D.; Zhang, X.M.; Gao, T.X.; Wan, R.J.; Zhang, P.D. Category composition and distributional patterns of ichthyoplankton in the Yellow River estuary during spring and summer 2007. J. Fish Sci. Chin. 2010, 17, 815–827. [Google Scholar]
- Deng, J.Y.; Jin, X.S. Study on fishery biodiversity and its conservation in Laizhou Bay and Yellow River Estuary. Zool. Res. 2000, 21, 76–82. [Google Scholar]
- Jin, X.S.; Deng, J.Y. Variations in community structure of fishery resources and biodiversity in the Laizhou Bay, Shandong. Biodivers. Sci. 2000, 8, 65–72. [Google Scholar]
- Shabanzadeh-Khoshrody, M.; Azadi, H.; Khajooeipour, A.; Nabavi-Pelesaraei, A. Analytical investigation of the effects of dam construction on the productivity and efficiency of farmers. J. Clean. Prod. 2016, 135, 549–557. [Google Scholar] [CrossRef]
- Wu, H.P.; Chen, J.; Xu, J.J.; Zeng, G.M.; Sang, L.H.; Liu, Q.; Yin, Z.J.; Dai, J.; Yin, D.; Liang, J. Effects of dam construction on biodiversity: A review. J. Clean. Prod. 2019, 221, 480–489. [Google Scholar] [CrossRef]
- Yang, J.M.; Li, J. Progress of Chinese marine ichthyology in the last 15 years. Stud. Mar. Sin. 1995, 36, 297–310. [Google Scholar]
- Zhu, X.H.; Miao, F.; Liu, D.; Xian, W.W. Spatiotemporal pattern and dominant component of fish community in the Yellow River estuary and its adjacent waters. Stud. Mar. Sin. 2001, 43, 141–151. [Google Scholar]
- Jiao, Y.M.; Zhang, X.H.; Li, H.X. Influence on fish diversity in the sea area off the Huanghe River estuary by the cutoff of water supply. Trans. Oceanol. Limnol. 1998, 4, 48–53. [Google Scholar]
- Muneo, O. An Atlas of the Early Stage Fishes in Japan; Tokai University Press: Tokyo, Japan, 1988. [Google Scholar]
- Zhao, C.Y.; Zhang, R.Z. Fish Eggs and Larvae in the Coastal Waters of China; Shanghai Scientific & Technical Publishers: Shanghai, China, 1985. [Google Scholar]
- Pianka, E.R. Ecology of the agamid lizard Amphibolurus isolepis in Western Australia. Copeia 1971, 3, 527–536. [Google Scholar] [CrossRef]
- Margalef, R. Information theory in ecology. Gen. System. 1958, 3, 36–71. [Google Scholar]
- Wilhm, J.L. Use of biomass units in Shannon’s formula. Ecology 1968, 49, 153–156. [Google Scholar] [CrossRef]
- Pielou, E.C. Ecological Diversity; Wiley: New York, NY, USA, 1975. [Google Scholar]
- Lepš, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Tian, M.C.; Sun, B.L.; Yang, J.M. Analysis of the fish fauna of the Bohai Sea. Stud. Mar. Sin. 1993, 34, 157–167. [Google Scholar]
- Liu, J.; Ning, P. Species composition and faunal characteristics of fishes in the Yellow Sea. Biodivers. Sci. 2011, 19, 764–769. [Google Scholar]
- Bauchot, M.L.; Pras, A. Guia de Los Peces de Mar de Espafiay de Europa; Ediciones Omega: Bracelona, Spain, 1987. [Google Scholar]
- Zhang, C.N.; Wang, J.; Zhou, A.H.; Ye, Q.; Feng, Y.Y.; Wang, Z.L.; Wang, S.D.; Xu, G.H.; Zou, J.X. Species-specific effect of microplastics on fish embryos and observation of toxicity kinetics in larvae. J. Hazard. Mater. 2021, 403, 123948. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, D.Y.; Hensley, D.A.; Appeldoorn, R.S. Pelagic spawning and egg transport in coral-reef fishes: A skeptical overview. Environ. Biol. Fish. 1988, 22, 3–14. [Google Scholar] [CrossRef]
- Downie, A.T.; Llling, B.; Faria, A.M.; Rummer, J.L. Swimming performance of marine fish larvae: Review of a universal trait under ecological and environmental pressure. Rev. Fish Biol. Fish. 2020, 30, 93–108. [Google Scholar] [CrossRef]
- Formicki, K.; Korzelecka-Orkisz, A.; Tański, A. The effect of an anthropogenic magnetic field on the early developmental stages of fishes—A review. Int. J. Mol. Sci. 2021, 22, 1210. [Google Scholar] [CrossRef] [PubMed]
- Amini Chermahini, M.; Shabani, A.; Naddafi, R.; Ghorbani, R.; Rabbaniha, M.; Noorinejad, M. Diversity, distribution, and abundance patterns of ichthyoplankton assemblages in some inlets of the northern Persian Gulf. J. Sea Res. 2021, 167, 101981. [Google Scholar] [CrossRef]
- Castillo-Rivera, M.; Zavala-Hurtado, J.A. Exploration of spatial and temporal patterns of fish diversity and composition in a tropical estuarine system of Mexico. Rev. Fish Boil. Fish. 2002, 12, 167–177. [Google Scholar] [CrossRef]
- Huang, D.J.; Su, J.L. The effects of the Huanghe River Delta on the circulation and transportation of larvae. Haiyang Xuebao 2002, 24, 104–111. [Google Scholar]
- Jjiang, M.; Shen, X.Q.; Wang, Y.L.; Yuan, Q.; Chen, L.F. Species of fish eggs and larvae and distribution in Changjiang Estuary and vicinity waters. Haiyang Xuebao 2006, 28, 171–174. [Google Scholar]
- Azeiteiro, U.M.; Bacelar-Nicolau, L.; Resende, P.; Goncalves, F.; Pereira, M.J. Larval fish distribution in shallow coastal waters off North Western Iberia (NE Atlantic). Estuar. Coast Shelf Sci. 2006, 69, 554–566. [Google Scholar] [CrossRef] [Green Version]
- Shan, X.J.; Jin, X.S. Diversity of fish community structure in the spring-time offshore waters at the Yangtze river estuary. Oceanol. Et Limngia Sin. 2011, 42, 32–40. [Google Scholar]
- Zhao, M.M.; Xu, Z.L. Species composition, temporal and spatial distributions and diversity of hoplocarida and decapods shrimps in summer and winter in Sanmen Bay. Chin. J. Zool. 2012, 46, 11–18. [Google Scholar]
- Yang, Y.Y.; Zhu, M.M.; Xu, B.Q.; Li, F.; Gao, Y.J.; Sun, C.X.; Wei, X.; Wang, B. Seasonal variation of community structure and diversity of ichthyoplankton in the Laizhou Bay. Haiyang Xuebao 2021, 43, 59–70. [Google Scholar]
- Connell, J.H. Diversity in tropical rainforests and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [Green Version]
- Sanders, H.L. Marine benthic diversity: A comparative study. Am. Nat. 1968, 102, 243–282. [Google Scholar] [CrossRef]
- Zhai, L.; Xu, B.D.; Ji, Y.P.; Ren, Y.P. Spatial pattern of fish assemblage and the relationship with environmental factors in Yellow River Estuary and its adjacent waters in summer. Chin. J. Appl. Ecol. 2015, 26, 2852–2858. [Google Scholar]
- Liu, F.; Chen, S.L.; Peng, J.; Chen, G.Q. Multi-scale variability of flow discharge and sediment load of Yellow River to sea and its impacts on the Estuary during the past 60 years. Acta Geogr. Sin. 2011, 3, 313–323. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Range and Mean Values (±sd) | 2011 | 2012 | 2013 | 2014 |
|---|---|---|---|---|---|
| SST (°C) | Range | 19.20–24.00 | 19.00–23.80 | 15.74–20.08 | 19.16–22.97 |
| mean values (±sd) | 21.05 ± 1.52 | 21.14 ± 1.54 | 17.80 ± 1.05 | 20.49 ± 1.08 | |
| SSS‰ | Range | 29.70–30.17 | 16.28–30.36 | 12.65–28.56 | 28.21–30.44 |
| mean values (±sd) | 29.93 ± 0.16 | 26.82 ± 3.88 | 26.97 ± 3.69 | 29.12 ± 0.61 | |
| Chl-a µg/L | Range | 1.93–9.49 | 1.14–6.92 | 2.15–6.83 | 0.67–3.83 |
| mean values (±sd) | 4.42 ± 2.58 | 3.49 ± 1.50 | 3.95 ± 1.54 | 2.48 ± 0.83 | |
| SPM (mg/L) | Range | 5.60–13.10 | 10.30–15.80 | 5.4–23.40 | 17.0–68.80 |
| mean values (±sd) | 8.96 ± 2.06 | 13.16 ± 1.81 | 13.65 ± 5.44 | 25.18 ± 11.99 | |
| pH | Range | 8.22–9.26 | 7.85–8.50 | 8.02–8.56 | 7.97–8.14 |
| mean values (±sd) | 8.48 ± 0.28 | 8.15 ± 0.17 | 8.27 ± 0.16 | 8.05 ± 0.06 | |
| DO (mg/L) | Range | 7.44–11.73 | 7.19–11.42 | 7.73–10.08 | 8.18–9.05 |
| mean values (±sd) | 8.73 ± 1.05 | 8.93 ± 1.23 | 8.32 ± 0.57 | 8.61 ± 0.30 |
| Family | Species | Developmental Stage | Ecological Type | Letter Number | |||
|---|---|---|---|---|---|---|---|
| Eggs | Larvae | Temperature Suitability | Type of Migration | Habitat Type | |||
| Clupeidae | Konosiruspunctatus | + | + | WT | MM | CPN | E |
| Sardinella zunasi | + | WT | MM | CPN | K | ||
| Engraulidae | Engraulis japonicus | + | + | WT | MM | CPN | D |
| Coilia mystus | + | + | WT | AM | CPN | J | |
| Thryssa kammalensis | + | WW | MM | CPN | H | ||
| Thrissa mystax | + | + | WW | MM | CPN | G | |
| Synodidae | Saurida elongata | + | WT | MS | CD | B | |
| Mugilidae | Liza haematocheilus | + | + | WT | ES | CPN | I |
| Serranidae | Lateolabraxjaponicus | + | WT | ES | CRA | M | |
| Sciaenidae | Johnius belengerii | + | WW | MM | CBD | W | |
| Atherinidae | Allanetta bleekeri | + | WW | MM | CPN | S | |
| Hemiramphidae | Hyporhamphus sajori | + | WT | ES | CPN | U | |
| Sillaginidae | Sillago sihama | + | WW | MM | CRA | C | |
| Trichiuridae | Eupleurogrammus muticus | + | + | WT | ES | CBD | A |
| Cynoglossidae | Cynoglossus joyneri | + | CT | ES | CD | N | |
| Pleuronectidae | Tanakius kitaharae | + | CT | MM | CD | Q | |
| Paralichthyidae | Paralichthys olivaceus | + | WT | MM | CD | R | |
| Syngnathidae | Syngnathus acus | + | WT | MM | CD | O | |
| Hippocampus coronatus | + | WT | MM | CD | Y | ||
| Salangidae | Protosalanx hyalocranius | + | WT | AM | CPN | v | |
| Gobiidae | Acentrogobius pflaumii | + | WT | ES | CD | X | |
| Amblychaeturichthys hexanema | + | WT | ES | CD | P | ||
| Chaeturichthys stigmatias | + | WT | ES | CD | T | ||
| Gobiidae sp. | + | L | |||||
| Unidentified species | + | F | |||||
| Year | Number of Fish Eggs (inds) | Species of Fish Eggs (kinds) | Number of Larvae (inds) | Species of Larvae (kinds) |
|---|---|---|---|---|
| 2011 | 270 (9.31 ± 27.25) | 7 | 37 (1.28 ± 3.15) | 5 |
| 2012 | 608 (25.33 ± 52.51) | 3 | 11 (0.46 ± 1.22) | 2 |
| 2013 | 1082 (28.47 ± 145.38) | 5 | 73 (1.85 ± 2.79) | 4 |
| 2014 | 580 (13.81 ± 27.92) | 6 | 22 (0.52 ± 1.29) | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, F.; Zhu, M.; Chen, W.; Su, B.; Yang, Y.; Wang, B. Interannual Variation of Ichthyoplankton Community Structure in the Yellow River Estuary, China. Water 2023, 15, 1040. https://doi.org/10.3390/w15061040
Li F, Zhu M, Chen W, Su B, Yang Y, Wang B. Interannual Variation of Ichthyoplankton Community Structure in the Yellow River Estuary, China. Water. 2023; 15(6):1040. https://doi.org/10.3390/w15061040
Chicago/Turabian StyleLi, Fan, Mingming Zhu, Wei Chen, Bo Su, Yanyan Yang, and Bin Wang. 2023. "Interannual Variation of Ichthyoplankton Community Structure in the Yellow River Estuary, China" Water 15, no. 6: 1040. https://doi.org/10.3390/w15061040
APA StyleLi, F., Zhu, M., Chen, W., Su, B., Yang, Y., & Wang, B. (2023). Interannual Variation of Ichthyoplankton Community Structure in the Yellow River Estuary, China. Water, 15(6), 1040. https://doi.org/10.3390/w15061040