


Pulling the Plug—Draining an Alpine Lake Failed to Eradicate Alien Minnows and Impacted Lower Trophic Levels

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
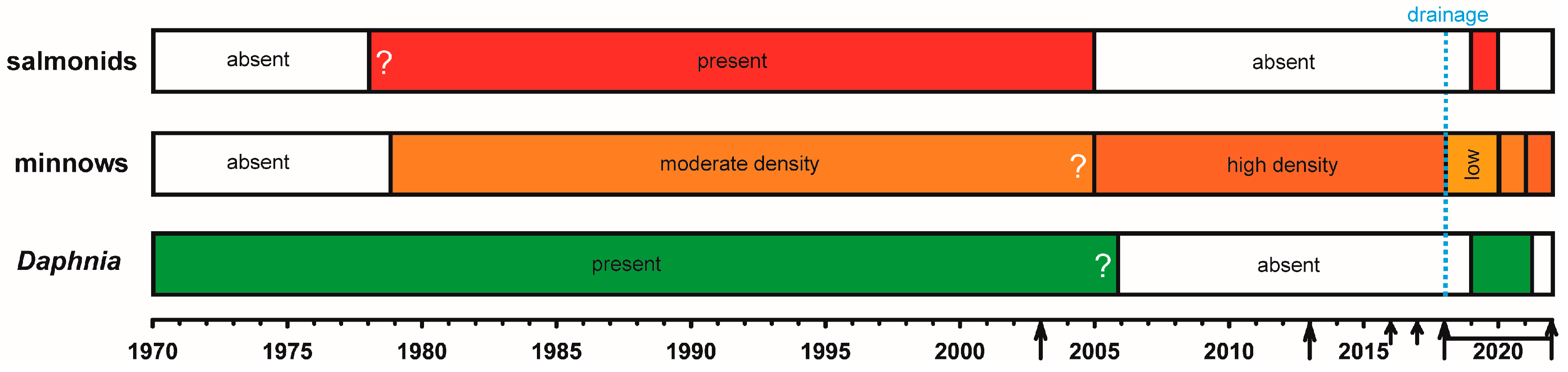
2.1. Study Area and Stocking History
2.2. Lake Drainage
2.3. Liming

2.4. Sample Collections
2.5. Fishing

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
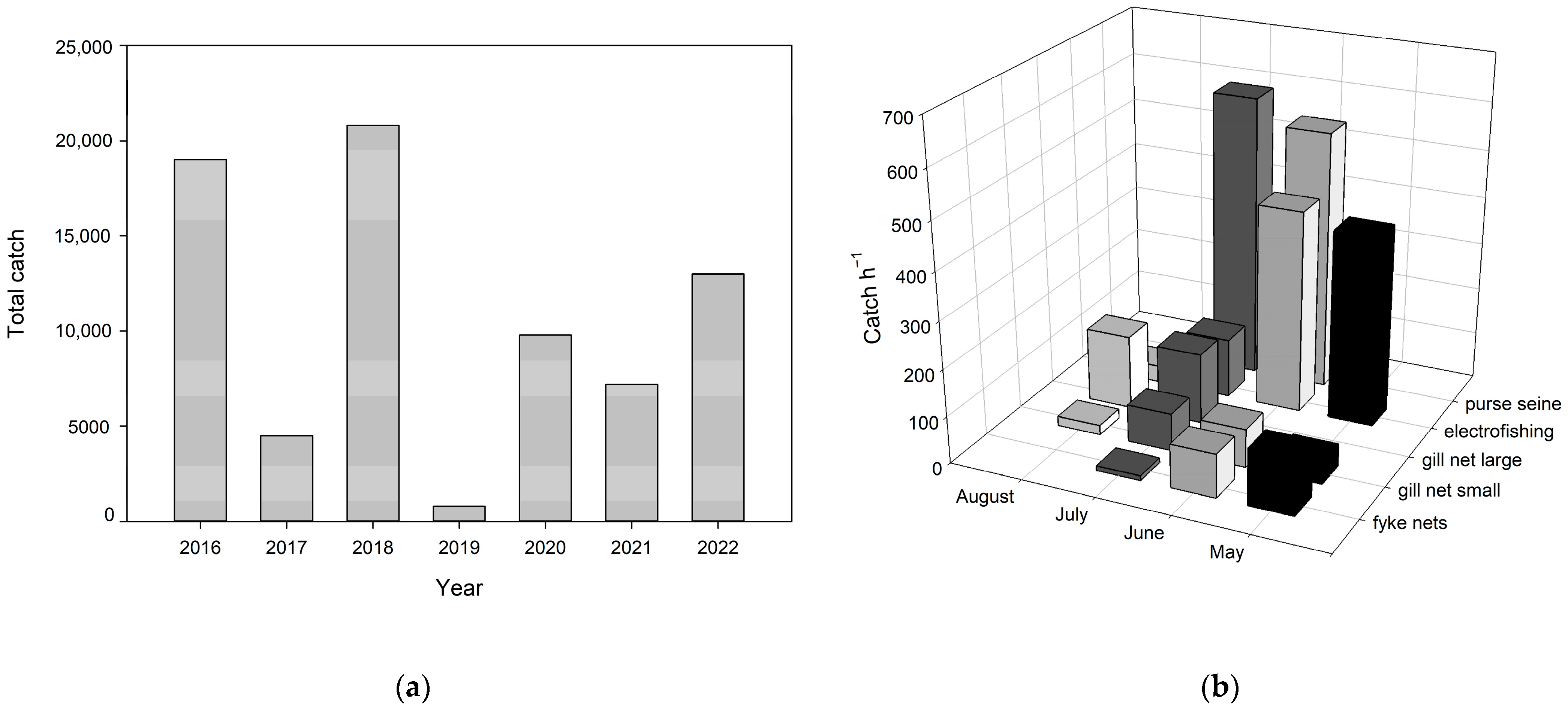
| Year | Gear | Dimensions | Total Time of Fishing Operation |
|---|---|---|---|
| 2003 | 1 multi-mesh gill net; deep lake | 50 m long × 1.5 m wide, 6.5–70 mm mesh size | 3 h |
| 2013 | 2 multi-mesh gill nets; deep lake | 30 m × 1.5 m, 5–55 mm | 26 h |
| Electrofishing | ELT-62 Grassl 2.2 kW | 3 h | |
| 2016 | 4 minnow traps | 1.5 long × 0.5 wide × 0.3 m high, 5 mm | 2 d |
| 2017 | bottle traps | 1.5 l PET bottles | Several weeks |
| 2018 | 4 rectangular minnow traps | 0.54 m × 0.25 m × 0.25 m, 3 mm | 13 d |
| 2019 | 4 umbrella minnow traps | 0.95 m × 0.95 m × 0.60 m, 5 mm | 16 d |
| 2 rectangular minnow traps | as above | ||
| 8 bottle traps | as above | ||
| 4 multi-mesh gillnets | 30 m × 1.5 m, 5–55 mm, effective mesh sizes 5, 6.25, 8, and 10 mm | 3 d | |
| 2020 | 14 umbrella traps | as above | 8 d |
| 2 small gill nets | 40 m × 0.3 m, (alternating 10 m of 5 mm and 6.25 mm mesh sizes) | 8 d | |
| 2 large gill nets | 50 m × 2 m, 6 mm | 3 d | |
| purse seine | 10 m (5 + 5 m wings) × 1.5 m, 7 mm | 3 d | |
| electrofishing | as above | 4 d | |
| 2021 | purse seine | 8 m × 0.5 m, 2 mm, cod-end 0.85 mm | 4 d |
| 2022 | purse seine 1 | 8 m × 0.8 m, 2 mm, 0.85 mm | 4 d |
| purse seine 2 | 16 m × 1 m, 2 mm, 0.85 mm | ||
| purse seine 3 | 12 m × 1 m, 2 mm, 0.85 mm |
2.6. Amphibians
3. Results
3.1. Lake Drainage
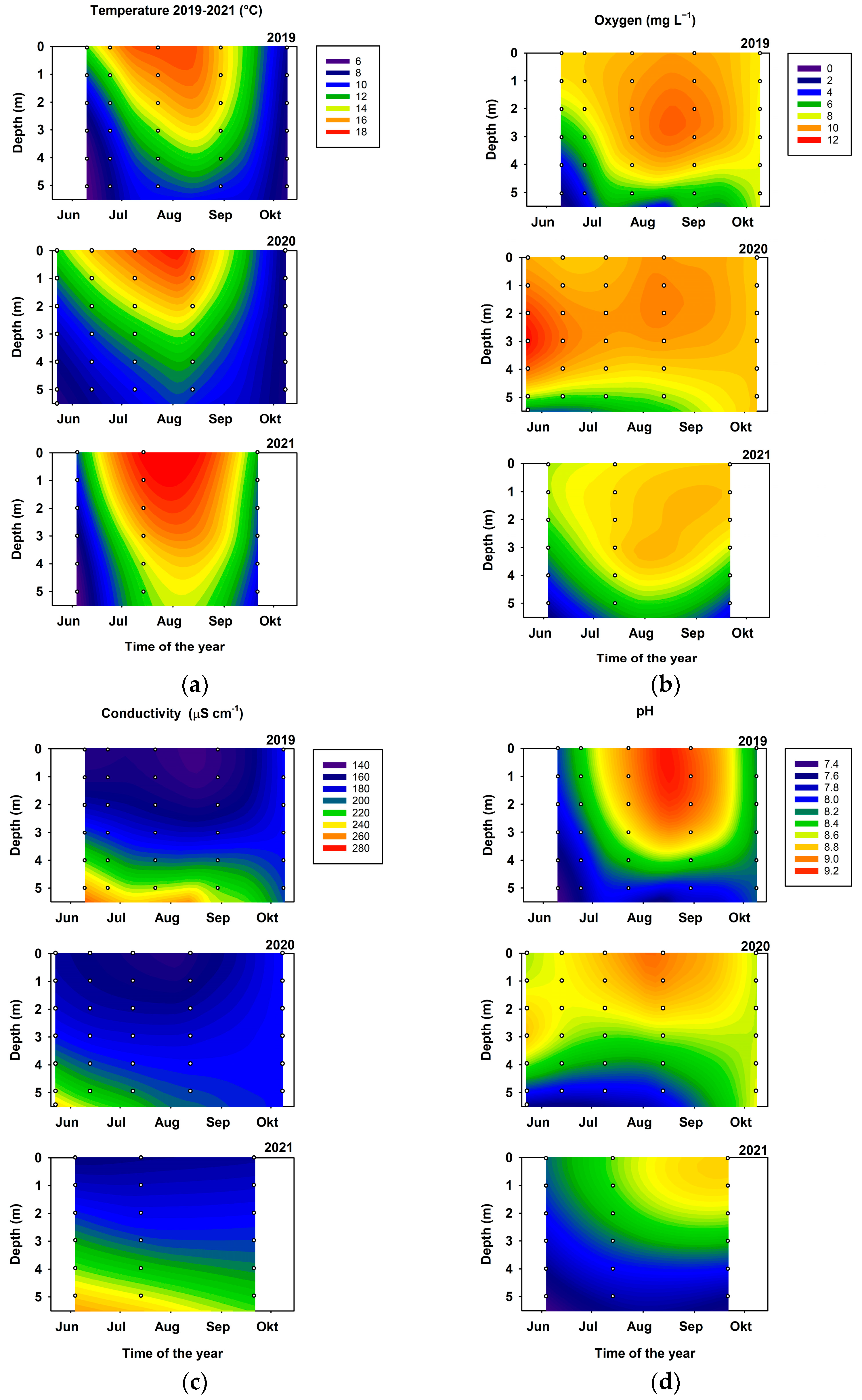
3.2. Stratification
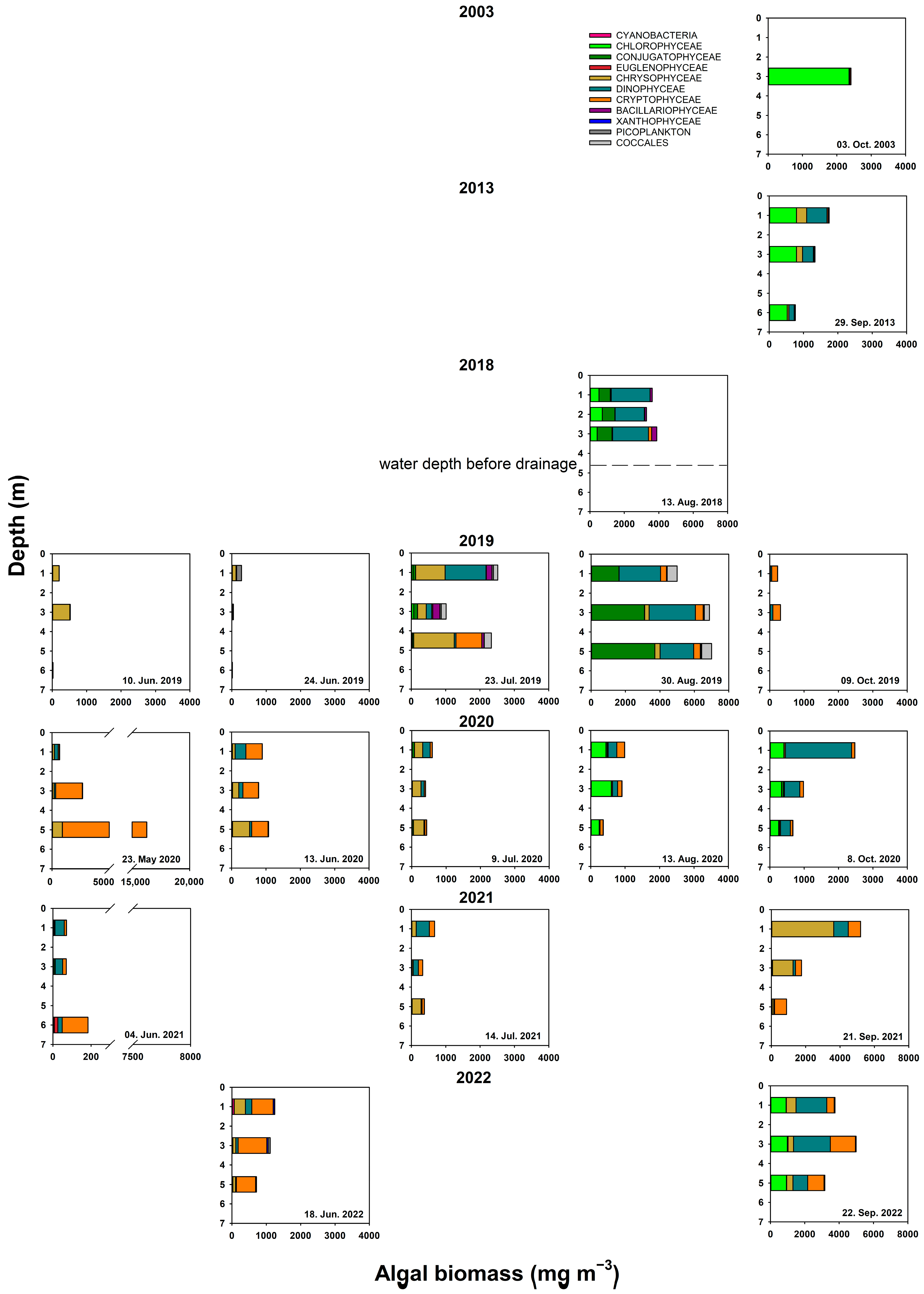
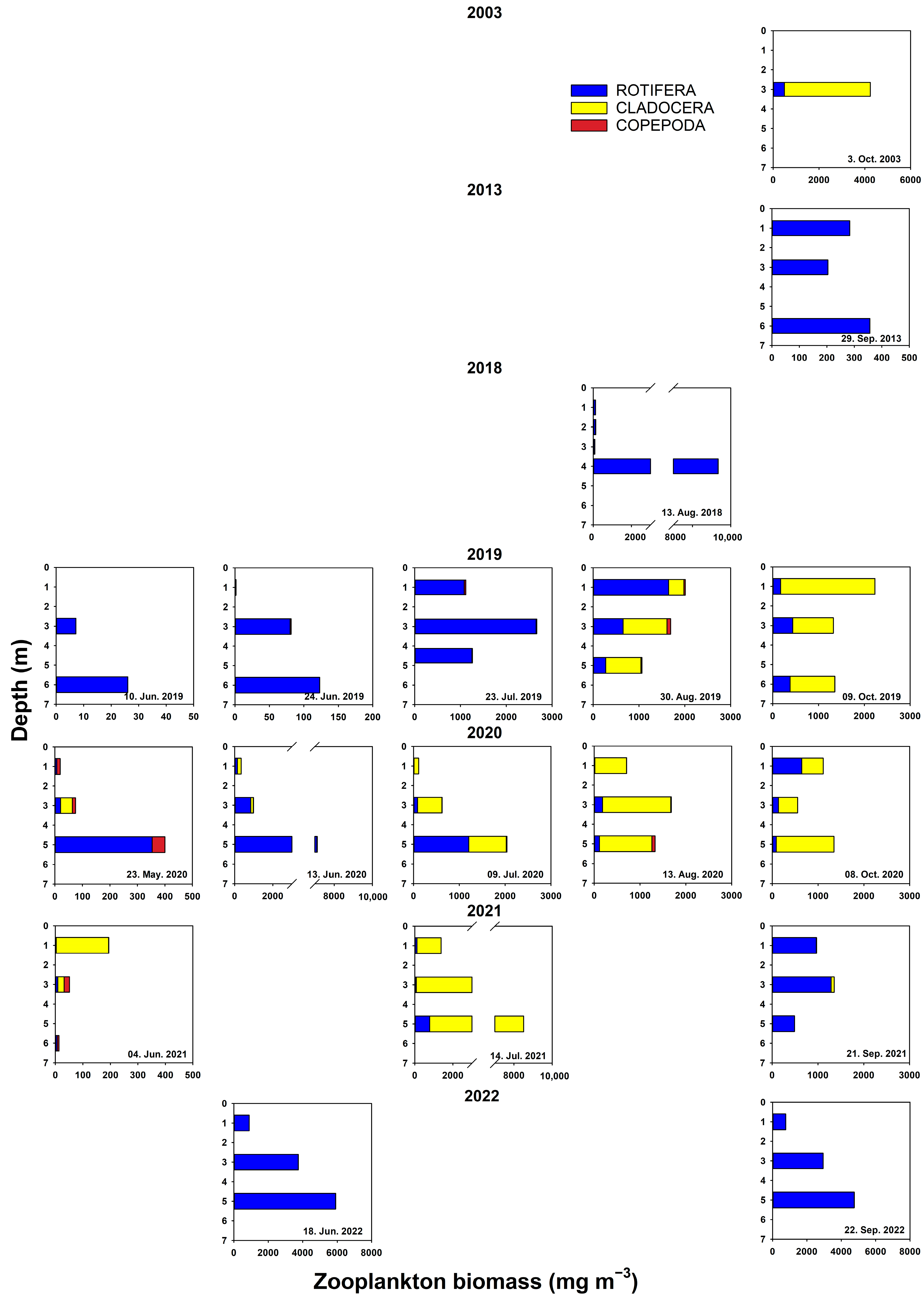
3.3. Plankton




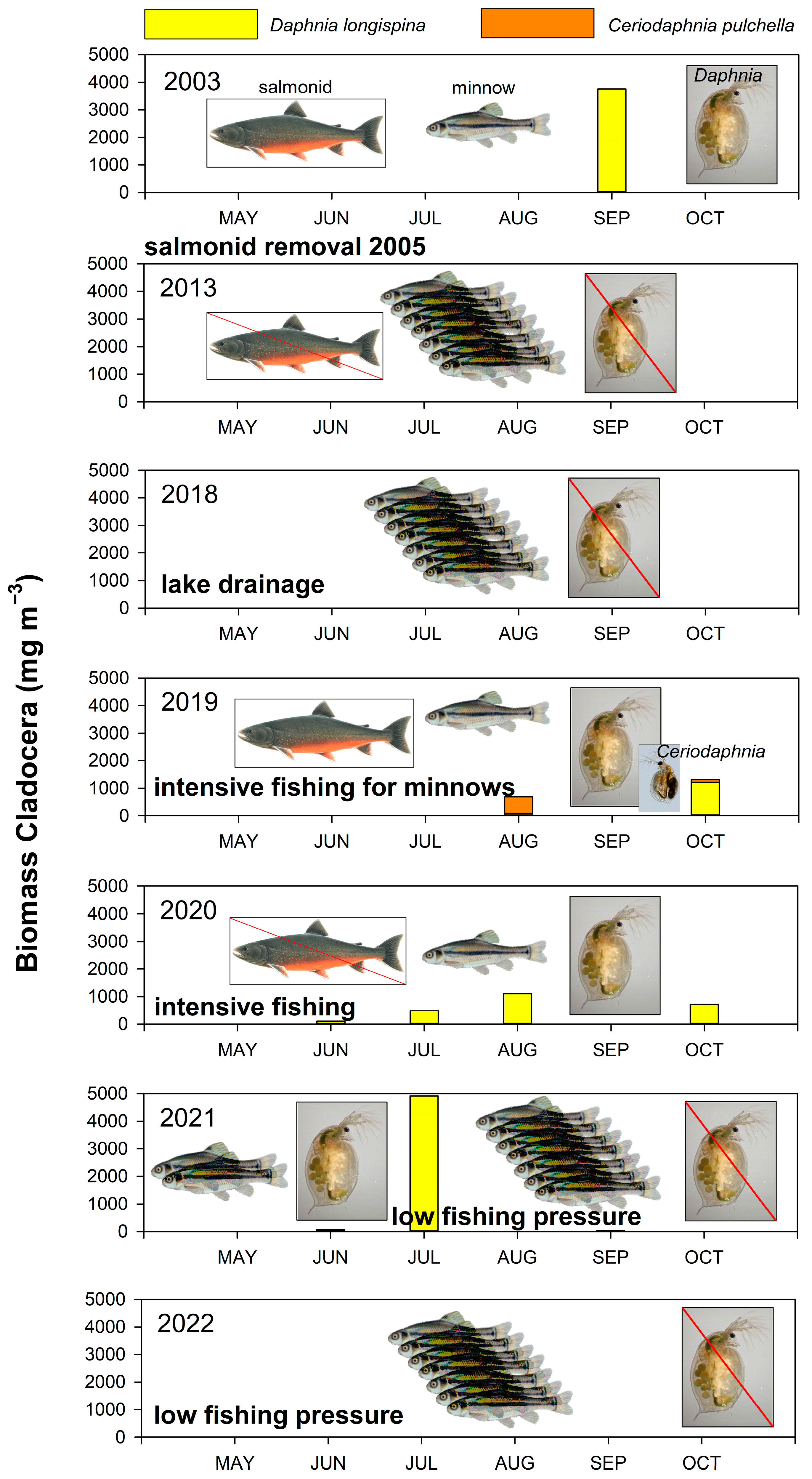
3.4. Fishing
3.5. Amphibians
4. Discussion
4.1. Fish Removal
4.2. Physical Environment and Impact of Fish on Lower Trophic Levels
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
| Depth (m) | Temp. (°C) | O2 (mg L−1) | O2 (%Sat) | pH | Cond. (µS cm−1) | |
| 3 October 2003 | 0.3 | 10.4 | 12.3 | 127 | 8.3 | 183 |
| 1 | 9.6 | 12.5 | 128 | 8.3 | 183 | |
| 2 | 9.3 | 12.6 | 127 | 8.4 | 183 | |
| 3 | 9.2 | 12.0 | 121 | 8.4 | 183 | |
| 4 | 9.1 | 12.0 | 121 | 8.4 | 184 | |
| 5 | 9.0 | 12.0 | 121 | 8.3 | 184 | |
| 6 | 9.0 | 11.9 | 120 | 8.3 | 184 | |
| 29 September 2013 | 0 | 10.0 | 10.4 | 115 | 7.9 | 189 |
| 1 | 9.4 | 10.5 | 112 | 8.0 | 190 | |
| 2 | 8.8 | 9.6 | 100 | 8.2 | 193 | |
| 3 | 8.3 | 9.0 | 93 | 8.2 | 196 | |
| 4 | 8.1 | 8.3 | 84 | 8.1 | 198 | |
| 5 | 7.9 | 8.0 | 81 | 8.1 | 198 | |
| 6 | 7.9 | 7.8 | 80 | 8.1 | 201 | |
| 7 | 7.8 | 7.4 | 75 | 8.1 | 204 | |
| 13 August 2018 | 0 | 20.0 | 8.8 | 122 | 9.0 | 142 |
| 1 | 18.7 | 9.2 | 122 | 9.0 | 145 | |
| 2 | 17.5 | 9.5 | 122 | 9.1 | 145 | |
| 3 | 14.0 | 12.6 | 156 | 9.0 | 180 | |
| 4 | 11.0 | 9.9 | 118 | 8.1 | 232 | |
| 18 June 2022 | 0 | 17.7 | 8.4 | 106 | 8.7 | 155 |
| 1 | 16.1 | 8.9 | 108 | 8.6 | 159 | |
| 2 | 12.9 | 10.1 | 114 | 8.6 | 171 | |
| 3 | 10.3 | 10.9 | 117 | 8.6 | 194 | |
| 4 | 8.4 | 10.1 | 102 | 8.2 | 211 | |
| 5 | 7.6 | 5.8 | 59 | 7.8 | 230 | |
| 22 September 2022 | 0 | 7.5 | 7.8 | 77 | 7.9 | 204 |
| 1 | 7.1 | 7.8 | 77 | 8.0 | 205 | |
| 2 | 7.0 | 7.8 | 76 | 8.0 | 206 | |
| 3 | 7.0 | 7.8 | 76 | 7.9 | 207 | |
| 4 | 6.9 | 7.7 | 75 | 7.9 | 208 | |
| 5 | 6.9 | 7.6 | 74 | 7.9 | 208 |
| Phytoplankton |
| Bacillariophyceae |
| Achnanthes sp. Bory, 1822 |
| Achnanthidium minutissimum (Kützing) Czarnecki, 1994 |
| Achnanthidium spp. Kützing, 1844 |
| Amphora sp. Ehrenberg ex Kützing, 1844 |
| Asterionella formosa Hassall, 1850 |
| Aulacoseira islandica (O. Müller) Simonsen, 1979 |
| Cyclotella spp. (Kützing) Brébisson, 1838, nom. et typ. cons. |
| Cymbella sp. C. Agardh, 1830, nom. et typ. cons. |
| Fragilaria spp. Lyngbye, 1819 |
| Lyngbya sp. C. Agardh ex Gomont, 1892, nom. et typ. cons. |
| Meridion circulare (Greville) C. Agardh, 1831 |
| Navicula sp. Bory, 1822 |
| Nitzschia acicularis (Kützing) W. Smith, 1853 |
| Nitzschia sp. Hassall, 1845, nom. cons. |
| Ulnaria acus (Kützing) Aboal, 2003 |
| Ulnaria delicatissima var. angustissima (Grunow) Aboal & P.C. Silva, 2004 |
| Chlorophyceae |
| Ankistrodesmus fusiformis Corda, 1838 |
| Ankistrodesmus spiralis (W.B. Turner) Lemmermann, 1908 |
| Botryococcus braunii Kützing, 1849 |
| Chlamydomonas spp. Ehrenberg, 1833, nom. cons. |
| Chlorococcales |
| Chlorococcales indet. |
| Coenochloris fottii (Hindák) Tsarenko, 1990 |
| Coenococcus planktonicus Korshikov, 1953 |
| Desmodesmus brasiliensis (Bohlin) E. Hegewald, 2000 |
| Dictyosphaerium subsolitarium Van Goor, 1924 |
| Elakatothrix cf. genevensis (Reverdin) Hindák, 1962 |
| Elakatothrix gelatinosa Wille, 1898 |
| Eutetramorus globosus Walton, 1918 |
| Koliella cf. longiseta (Vischer) Hindák, 1963 |
| Lagerheimia ciliata (Lagerheim) Chodat, 1895 |
| Monoraphidium griffithii (Berkeley) Komárková-Legnerová, 1969 |
| Monoraphidium tortile (West & G.S. West) Komárková-Legnerová, 1969 |
| Nephrocytium agardhianum Nägeli, 1849 |
| Oedogonium sp. Link ex Hirn, 1900 |
| Oocystis spp. Nägeli ex A. Braun, 1855 |
| Pandorina morum (O.F. Müller) Bory, 1826 |
| Pediastrum duplex Meyen, 1829 |
| Planktosphaeria gelatinosa G.M. Smith, 1918 |
| Pseudopediastrum boryanum (Turpin) E. Hegewald, 2005 |
| Pseudopediastrum integrum (Nägeli) M. Jena & C. Bock, 2014 |
| Pseudosphaerocystis lacustris (Lemmermann) Nováková, 1965 |
| Pteromonas sp. Seligo, 1887 |
| Raphidocelis danubiana (Hindák) Marvan, Komárek & Comas, 1984 |
| Scenedesmus ecornis (Ehrenberg) Chodat, 1926 |
| Scenedesmus obtusus Meyen, 1829 |
| Scenedesmus spp. Meyen, 1829 |
| Scenedesmus subspicatus Chodat, 1926 |
| Scourfieldia sp. G.S. West, 1912 |
| Sphaerocystis schroeteri Chodat, 1897 |
| Stauridium tetras (Ehrenberg) E. Hegewald, 2005 |
| Tetrabaena socialis (Dujardin) H. Nozaki & M. Itoh, 1994 |
| Tetrachlorella alternans (G.M. Smith) Korshikov, 1939 |
| Tetradesmus obliquus (Turpin) M.J. Wynne, 2016 |
| Tetraedron sp. Kützing, 1845 |
| Vitreochlamys fluviatilis (F. Stein) Batko, 1970 |
| Chrysophyceae |
| Bitrichia chodatii (Reverdin) Chodat, 1926 |
| Chromulina sp. Cienkowsky, 1870 |
| Chrysochromulina parva Lackey, 1939 |
| Chrysococcus sp. Klebs, 1892 |
| Chrysolykos planctonicus B. Mack, 1951 |
| Chrysophyceae spp. Pascher, 1914 |
| Dinobryon cylindricum O.E. Imhof, 1887 |
| Dinobryon cylindricum var. alpinum (O.E. Imhof) H. Bachmann, 1911 |
| Dinobryon divergens O.E. Imhof, 1887 |
| Dinobryon sociale (Ehrenberg) Ehrenberg, 1834 |
| Mallomonas spp. Perty, 1852 |
| Ochromonas sp. Vysotskii, 1887 |
| Pseudokephyrion sp. Pascher, 1913 |
| Pseudopedinella erkensis Skuja, 1948 |
| Uroglena sp. Ehrenberg, 1834 |
| Coccales |
| Coccales indetermined |
| Conjugatophyceae |
| Cosmarium bioculatum Brébisson ex Ralfs, 1848 |
| Cosmarium neodepressum G.J.P. Ramos & C.W.N. Moura, 2020 |
| Cosmarium pseudopyramidatum P. Lundell 1871 |
| Cosmarium reniforme (Ralfs) W. Archer, 1874 |
| Cosmarium spp. Corda ex Ralfs, 1848 |
| Cosmarium tenue W. Archer, 1868 |
| Gonatozygon brebissonii De Bary, 1858 |
| Gonatozygon monotaenium De Bary, 1856 |
| Hyalotheca dissiliens Brébisson ex Ralfs, 1848 |
| Mougeotia sp. C. Agardh, 1824 |
| Pleurotaenium sp. Nägeli, 1849 |
| Spirogyra sp. Link, 1820 |
| Spondylosium pygmaeum Cooke, 1880 |
| Staurastrum acutum var. varians (Raciborski) Coesel & Meesters, 2013 |
| Staurastrum cf. paradoxum Meyen ex Ralfs, 1848 |
| Staurastrum cf. polymorphum Brébisson, 1848 |
| Staurastrum muticum Brébisson ex Ralfs, 1848 |
| Staurastrum sp. Meyen ex Ralfs, 1848 |
| Staurodesmus cuspidatus (Brébisson) Teiling, 1967 |
| Staurodesmus patens (Nordstedt) Croasdale, 1957 |
| Staurodesmus sp. Teiling, 1948 |
| Cryptophyceae |
| Cryptomonas curvata Ehrenberg, 1832 |
| Cryptomonas erosa Ehrenberg, 1832 |
| Cryptomonas marssonii Skuja, 1948 |
| Cryptomonas ovata Ehrenberg, 1832 |
| Cryptomonas pyrenoidifera Geitler, 1922 |
| Cryptomonas spp. Ehrenberg, 1831 |
| Plagioselmis nannoplanctica (Skuja) G. Novarino, I.A.N. Lucas & Morrall, 1994 |
| Rhodomonas pusilla (Bachmann) Javornický, 1967 |
| Cyanobacteria |
| Anabaena sp. Bory ex Bornet & Flahault, 1886 |
| Anathece minutissima (West) Komárek, Kastovsky & Jezberová, 2011 |
| Aphanocapsa delicatissima West & G.S. West, 1912 |
| Aphanocapsa elachista West & G.S. West, 1894 |
| Aphanothece sp. Nägeli, 1849 |
| Chroococcus minutus (Kützing) Nägeli, 1849 |
| Coelosphaerium kuetzingianum Nägeli, 1849 |
| Dactylococcopsis fascicularis Lemmermann, 1898 |
| Eucapsis aphanocapsoides (Skuja) Komárek & Hindák, 2016 |
| Limnococcus limneticus (Lemmermann) Komárková, Jezberová, O. Komárek & Zapomelová, 2010 |
| Merismopedia glauca (Ehrenberg) Kützing, 1845 |
| Merismopedia sp. Meyen, 1839 |
| Nostoc sp. Vaucher ex Bornet & Flahault, 1886 |
| Oscillatoria limosa C. Agardh ex Gomont, 1892 |
| Oscillatoria sp. Vaucher ex Gomont, 1892 |
| Oscillatoriaceae Gen. sp. |
| Planktolyngbya limnetica (Lemmermann) Komárková-Legnerová & Cronberg, 1992 |
| Pseudanabaena catenata Lauterborn, 1915 |
| Pseudanabaena limnetica (Lemmermann) Komárek, 1974 |
| Pseudanabaena sp. Lauterborn, 1915 |
| Snowella lacustris (Chodat) Komárek & Hindák, 1988 |
| Dinophyceae |
| Ceratium hirundinella (O.F. Müller) Dujardin, 1841 |
| Glenodinium sp. Ehrenberg, 1836 |
| Gymnodinium lantzschii Utermöhl, 1925 |
| Gymnodinium spp. F. Stein, 1878 |
| Gymnodinium uberrimum (G.J. Allman) Kofoid & Swezy, 1921 |
| Parvodinium cf. umbonatum (F. Stein) Carty, 2008 |
| Peridinium cinctum (O.F. Müller) Ehrenberg, 1832 |
| Peridinium sp. Ehrenberg, 1830 |
| Peridinium volzii Lemmermann, 1905 |
| Peridinium willei Huitfeldt-Kaas, 1900 |
| Euglenophyceae |
| Euglena spp. Ehrenberg, 1830 |
| Lepocinclis spirogyroides B. Marin & Melkonian, 2003 |
| Menoidium sp. Perty, 1852 |
| Trachelomonas cf. hispida (Perty) F. Stein, 1878 |
| Trachelomonas sp. Ehrenberg, 1834 |
| Trachelomonas volvocina (Ehrenberg) Ehrenberg, 1834 |
| Picoplankton |
| µ-Algen (Picoplankton excl. Bacteria) |
| Flagellata indet. |
| Xanthophyceae |
| Tetraëdriella jovetii (Bourrelly) Bourrelly, 1968 |
| Tribonema sp. Derbès & Solier, 1851 |
| Zooplankton |
| Rotifera |
| Anuraeopsis fissa (Gosse, 1851) |
| Ascomorpha ecaudis Perty, 1850 |
| Asplanchna priodonta Gosse, 1850 |
| Bdelloidea Gen. sp. |
| Cephalodella cf. ventripes (Dixon-Nuttall, 1901) |
| Cephalodella gibba (Ehrenberg, 1830) |
| Cephalodella sp. |
| Cephalodella sterea (Gosse, 1887) |
| Collotheca sp. |
| Colurella obtusa (Gosse, 1886) |
| Colurella sp. |
| Encentrum lutra Wulfert, 1936 |
| Epiphanes brachionus (Ehrenberg, 1837) |
| Filinia passa (Müller, 1786) |
| Filinia terminalis (Plate, 1886) |
| Keratella cochlearis (Gosse, 1851) |
| Keratella cochlearis f. micracantha (Lauterborn, 1900) |
| Keratella cochlearis var. hispida (Lauterborn, 1898) |
| Keratella tecta (Gosse, 1851) |
| Keratella testudo (Ehrenberg, 1832) |
| Lecane closterocerca (Schmarda, 1859) |
| Lecane luna (Müller, 1776) |
| Lecane lunaris (Ehrenberg, 1832) |
| Lecane sp. |
| Lepadella patella (Müller, 1773) |
| Lepadella patella persimilis De Ridder, 1961 |
| Lepadella quadricarinata (Stenroos, 1898) |
| Lepadella sp. |
| Lophocharis cf. gracilis Dvořáková, 1960 |
| Monommata sp. |
| Mytilina mucronata (Müller, 1773) |
| Pleurotrocha petromyzon Ehrenberg, 1830 |
| Polyarthra dolichoptera Idelson, 1925 |
| Polyarthra sp. |
| Proales fallaciosa Wulfert, 1937 |
| Synchaeta cf. kitina Rousselet, 1902 |
| Synchaeta cf. pectinata Ehrenberg, 1832 |
| Synchaeta cf. tremula/kitina |
| Synchaeta lakowitziana Lucks, 1930 |
| Synchaeta pectinata Ehrenberg, 1832 |
| Synchaeta spp. |
| Synchaeta tremula (Müller, 1786) |
| Trichocerca iernis (Gosse, 1887) |
| Trichocerca longiseta (Schrank, 1802) |
| Trichocerca porcellus (Gosse, 1851) |
| Trichotria pocillum (Müller, 1776) |
| Cladocera |
| Acroperus harpae (Baird, 1834) |
| Alona rectangula G.O. Sars, 1862 |
| Alona sp. |
| Alona quadrangularis (O.F. Müller, 1776) |
| Ceriodaphnia pulchella G.O. Sars, 1862 |
| Daphnia longispina (O.F. Müller, 1776) |
| Copepoda |
| Eucyclops serrulatus (Fischer, 1851) |
References
- Sodhi, N.S.; Brook, B.W.; Bradshaw, C.J.A. Causes and Consequences of Species Extinction. In The Princeton Guide to Ecology; Levin, S.A., Carpenter, S.R., Godfra, H.C.J., Kinzig, A.P., Loreau, M., Losos, J.B., Walker, B., Wilcove, D.S., Eds.; Princeton University Press: Princeton, NJ, USA, 2012; pp. 514–520. [Google Scholar] [CrossRef]
- Knapp, R.A.; Matthews, K.E.; Sarnelle, O. Resistence and resilience of Alpine Lake Fauna to Fish Introductions. Ecol. Monogr. 2001, 71, 401–421. [Google Scholar] [CrossRef]
- Ventura, M.; Tiberti, R.; Buchaca, T.; Buñay, D.; Sabás, I.; Miró, A. Why Should We Preserve Fishless High Mountain Lakes? In High Mountain Conservation in a Changing World; Catalan, J., Ninot, J.M., Aniz, M., Eds.; Advances in Global Change Research; Springer: Cham, Switzerland, 2017; Volume 62, pp. 181–205. [Google Scholar] [CrossRef]
- Pechlaner, R. Salmonideneinsätze in Hochgebirgsseen und-Tümpel der Ostalpen. Verh. Int. Ver. Limnol. 1966, 16, 1182–1191. [Google Scholar] [CrossRef]
- Pechlaner, R. Historical Evidence for the Introduction of Alpine Charr into High-Mountain Lakes of the Alps by Man. In Biology of the Alpine Charr: Proceedings of the International Symposium on Alpine Charr; Johnson, L., Burns, B.L., Eds.; University of Manitoba Press: Winnipeg, MB, Canada, 1984; pp. 549–557. [Google Scholar]
- Bahls, P. The Status of Fish Populations and Management of High Mountain Lakes in the Western United States. NW Sci. 1992, 66, 183–193. [Google Scholar]
- Porcel, S.; Saas, J.F.; Marinone, M.C.; Izaguirre, I.; Lancelotti, J. Influence of the Fish Introduction in Lakes of the Arid Southern Patagonia. In Freshwater and Wetlands of Patagonia; Mataloni, G., Quintana, R.D., Eds.; Springer Nature: Cham, Switzerland, 2022; pp. 409–433. [Google Scholar] [CrossRef]
- Schabetsberger, R.; Grill, S.; Hauser, G.; Wukits, P. Zooplankton Successions in lakes with Contrasting Impacts of Amphibian and Fish Predators. Internat. Rev. Hydrobiol. 2006, 91, 197–221. [Google Scholar] [CrossRef]
- Miró, A.; Sabás, I.; Ventura, M. Large Negative Effect of Non-Native Trout and Minnows on Pyrenean Lake Amphibians. Biol. Cons. 2018, 218, 144–153. [Google Scholar] [CrossRef]
- Schindler, D.W.; Parker, B.R. Biological Pollutants: Alien Fishes in Mountain Lakes. Water Air Soil Pollut. Focus 2002, 2, 379–397. [Google Scholar] [CrossRef]
- Brancelj, A. The Extinction of Arctodiaptomus alpinus (Copepoda) Following the Introduction of Charr into a Small Alpine Lake Dvojno Jezero (NW Slovenia). Aquat. Ecol. 1999, 33, 355–361. [Google Scholar] [CrossRef]
- Schabetsberger, R.; Luger, M.; Drozdowski, G.; Jagsch, A. Only the Small Survive: Monitoring Long-Term Changes in the Zooplankton Community of an Alpine Lake after Fish Introduction. Biol. Inv. 2009, 11, 1335–1345. [Google Scholar] [CrossRef]
- Sienkiewicz, E.; Gąsiorowski, M. The Effect of Fish Stocking on Mountain Lake Plankton Communities Identified Using Palaeobiological Analyses of Bottom Sediment Cores. J. Palaeolimnol. 2016, 55, 129–150. [Google Scholar] [CrossRef]
- Pope, K.L.; Hannelly, E.C. Response of Benthic Macroinvertebrates to Whole-Lake, Non-Native Fish Treatments in Mid-Elevation Lakes of the Trinity Alps, California. Hydrobiologia 2013, 714, 201–215. [Google Scholar] [CrossRef]
- Pilliod, D.S.; Peterson, C.R. Local and Landscape Effects of Introduced Trout on Amphibians in Historically Fishless Watersheds. Ecosystems 2001, 4, 322–333. [Google Scholar] [CrossRef]
- Denoël, M.; Džukić, G.; Kalezić, M.L. Effect of Widespread Fish Introductions on Paedomorphic Newts in Europe. Conserv. Biol. 2005, 19, 162–170. [Google Scholar] [CrossRef]
- Tiberti, R.; von Hardenberg, A. Impact of Introduced Fish on Common Frog (Rana temporaria) Close to its Altitudinal Limit in Alpine Lakes. Amphib.-Reptil. 2012, 33, 303–307. [Google Scholar] [CrossRef]
- Denoël, M.; Ficetola, G.F.; Sillero, N.; Džukić, G.; Kalezić, M.L.; Vukov, T.D.; Muhovic, I.; Ikovic, V.; Lejeune, B. Traditionally Managed Landscapes do not Prevent Amphibian Decline and the Extinction of Paedomorphosis. Ecol. Monogr. 2019, 89, e01347. [Google Scholar] [CrossRef]
- Orizaola, G.; Braña, F. Effect of Salmonid Introduction and Other Environmental Characteristics on Amphibian Distribution and Abundance in Mountain Lakes of Northern Spain. Anim. Conserv. 2006, 9, 171–178. [Google Scholar] [CrossRef]
- Gacia, E.; Buchaca, T.; Bernal-Mendoza, N.; Sabás, I.; Ballesteros, E.; Ventura, M. Non-Native Minnows Threaten Quillwort Populations in High Mountain Shallow Lakes. Front. Plant Sci. 2018, 9, 329. [Google Scholar] [CrossRef] [PubMed]
- Izaguirre, I.; Lancelotti, J.; Saad, J.F.; Porcel, S.; O’Farrell, I.; Marinone, M.C.; Roesler, I.; Dieguez, M.C. Influence of Fish Introduction and Water Level Decrease on Lakes of the Arid Patagonian Plateaus with Importance for Biodiversity Conservation. Glob. Ecol. Conserv. 2018, 14, e00391. [Google Scholar] [CrossRef]
- Clearwater, S.J.; Hickey, C.W.; Martin, M.I. Overview of Potential Piscicides and Molluscicides for Controlling Aquatic Pest Species in New Zealand; Science Conservation 283; Science and Technical Publishing, Department of Conservation: Wellington, New Zealand, 2008; pp. 1–75. [Google Scholar]
- Britton, J.R.; Gozlan, E.; Copp, G.H. Managing Non-Native Fish in the Environment. Fish Fish. 2011, 12, 256–274. [Google Scholar] [CrossRef]
- Rytwinski, T.; Taylor, J.J.; Donaldson, L.A.; Britton, J.R.; Browne, D.R.; Gresswell, R.E.; Lintermans, M.; Prior, K.A.; Pellatt, M.G.; Vis, C.; et al. The Effectiveness of Non-Native Fish Removal Techniques in Freshwater Ecosystems: A Systematic Review. Environ. Rev. 2019, 27, 71–94. [Google Scholar] [CrossRef]
- Simberloff, D. Maintenance Management and Eradication of Established Aquatic Invaders. Hydrobiologia 2021, 848, 2399–2420. [Google Scholar] [CrossRef]
- Ryan, M.E.; Palen, W.J.; Adams, M.J.; Rochefort, R.M. Amphibians in the Climate Vice: Loss and Restoration of Resilience of Montane Wetland Ecosystems in the Western US. Front. Ecol. Environ. 2014, 12, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Knapp, R.A.; Sarnelle, O. Recovery after Local Extinction: Factors Affecting Re-Establishment of Alpine Lake Zooplankton. Ecol. Appl. 2008, 18, 1850–1859. [Google Scholar] [CrossRef] [PubMed]
- Knapp, R.A.; Hawkins, C.P.; Ladau, J.; McClory, J.G. Fauna of Yosemite National Park has Resistance but High Resilience to Fish Introductions. Ecol. Appl. 2005, 15, 835–847. [Google Scholar] [CrossRef]
- Knapp, R.A.; Boiano, D.M.; Vredenburg, V.T. Removal of Non-Native Fish Results in Population Expansion of a Declining Amphibian (Mountain Yellow-Legged Frog, Rana muscosa). Biol. Conserv. 2007, 135, 11–20. [Google Scholar] [CrossRef]
- Kats, L.B.; Ferrer, R.P. Alien Predators and Amphibian Declines: Review of Two Decades of Science and the Transition to Conservation. Divers. Distrib. 2003, 9, 99–110. [Google Scholar] [CrossRef]
- Vredenburg, V.T. Reversing Introduced Species Effects: Experimental Removal of Introduced Fish Leads to Rapid Recovery of a Declining Frog. Proc. Nat. Acad. Sci. USA 2004, 101, 7646–7650. [Google Scholar] [CrossRef] [PubMed]
- Knapp, R.A.; Fellers, G.M.; Kleeman, P.M.; Miller, D.A.W.; Vredenburg, V.T.; Rosenblum, E.B.; Briggs, C.J. Large-Scale Recovery of an Endangered Amphibian Despite Ongoing Exposure to Multiple Stressors. Proc. Nat. Acad. Sci. USA 2016, 113, 11889–11894. [Google Scholar] [CrossRef]
- Denoël, M.; Winandy, L. The importance of Phenotype Diversity in Conservation: Resilience of Palmate Newt Morphotypes after Fish Removal in Larzac Ponds (France). Biol. Conserv. 2015, 192, 402–408. [Google Scholar] [CrossRef]
- Komposch, C.; Kreiner, D. Gesäuse—An Alpine National Park of Endemic Species and Biodiversity Research. Eco-Mont 2018, 10, 79–83. [Google Scholar] [CrossRef]
- Pavuza, R.; Stummer, G. Geologie/Hydrologie/Karst-und Höhlenkunde des Sulzkarsees und seiner Umgebung; Report to the National Park Gesäuse; National Park Gesäuse: Weng im Gesäuse, Austria, 2004; pp. 1–13. [Google Scholar]
- Schwoerbel, J. Methoden der Hydrobiologie; Gustav Fischer: Stuttgart, Germany, 1993. [Google Scholar]
- Utermöhl, H. Zur Vervollkommnung der Quantitativen Phytoplanktonmethodik. Mitt. Int. Ver. Theor. Angew. Limnol. 1958, 9, 1–38. [Google Scholar]
- Deisinger, G. Leitfaden zur Bestimmung der Planktischen Algen der Kärntner Seen und ihrer Biomasse; Kärntner Institut für Seenforschung: Klagenfurt am Wörthersee, Austria, 1984; pp. 1–64. [Google Scholar]
- Ruttner-Kolisko, A. Suggestions for Biomass Calculation of Plankton Rotifers. Arch. Hydrobiol. Beihefte. Ergeb. Limnol. 1977, 8, 71–76. [Google Scholar]
- Downing, J.A.; Rigler, F.H. A Manual on Methods for the Assessment of Secondary Production in Fresh Waters, 2nd ed.; IBP Handbook 17; Blackwell Scientific Publications: Oxford, UK, 1984. [Google Scholar]
- Begon, M. Estimating Animal Abundance: Capture-Recapture for Biologists; E. Arnold: London, UK, 1979. [Google Scholar]
- Hasitschka, J. Die Geschichte der Sulzkaralm; Report to the National Park Gesäuse; National Park Gesäuse: Weng im Gesäuse, Austria, 2004; pp. 1–24. [Google Scholar]
- Ling, S. Rotenone—A Review of Its Toxicity and Use for Fisheries Management; Science for Conservation; Science and Technical Publishing, Department of Conservation: Wellington, New Zealand, 2008; Volume 211, pp. 1–40. [Google Scholar]
- Beaune, D.; Castelnau, F.; Sellier, Y.; Cucherousset, J. Native Top-Predator Cannot Eradicate an Invasive Fish from Small Pond Ecosystems. J. Nat. Conserv. 2019, 50, 125713. [Google Scholar] [CrossRef]
- Jersabek, C.; Schabetsberger, R. Limnological Aspects of an Alpine Karst Lake with Extreme Changes in Water Level. Limnologica 1996, 26, 1–13. [Google Scholar]
- Miskimmin, B.M.; Donahue, W.F.; Watson, D. Invertebrate Community Response to Experimental Lime (Ca(Oh)2) Treatment of an Eutrophic Pond. Aquat. Sci. 1995, 57, 20–30. [Google Scholar] [CrossRef]
- Winandy, L.; Legrand, P.; Denoël, M. Habitat Selection and Reproduction of Newts in Networks of Fish and Fishless Aquatic Patches. Anim. Behav. 2017, 123, 107–115. [Google Scholar] [CrossRef]
- Ludwig, G.; Sinsch, U.; Pelster, B. Migratory Behaviour during Autumn and Hibernation Site Selection in Common Frogs (Rana temporaria) at High Altitude. Herp. J. 2013, 23, 121–124. [Google Scholar]



| 2003 | 2013 | 2019 | 2021 | |
|---|---|---|---|---|
| TP | 0.015 | 0.01 | 0.136 | 0.012 |
| TDP | 0.0039 | n.a. | 0.003 | 0.004 |
| PO43−-P | n.a. | n.a. | 0.0033 | <0.0033 |
| NH4+-N | <0.002 | 0.016 | 2.176 | n.a. |
| NO2−-N | <0.003 | <0.015 | 0.003 | n.a. |
| NO3−-N | 0.13 | 0.102 | <0.25 | <0.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schabetsberger, R.; Jersabek, C.D.; Maringer, A.; Kreiner, D.; Kaltenbrunner, M.; Blažková, P.; Pokorný, P.; Denoël, M.; Emmerstorfer, H.; Lipovnik, C.; et al. Pulling the Plug—Draining an Alpine Lake Failed to Eradicate Alien Minnows and Impacted Lower Trophic Levels. Water 2023, 15, 1332. https://doi.org/10.3390/w15071332
Schabetsberger R, Jersabek CD, Maringer A, Kreiner D, Kaltenbrunner M, Blažková P, Pokorný P, Denoël M, Emmerstorfer H, Lipovnik C, et al. Pulling the Plug—Draining an Alpine Lake Failed to Eradicate Alien Minnows and Impacted Lower Trophic Levels. Water. 2023; 15(7):1332. https://doi.org/10.3390/w15071332
Chicago/Turabian StyleSchabetsberger, Robert, Christian D. Jersabek, Alexander Maringer, Daniel Kreiner, Magdalena Kaltenbrunner, Pavlína Blažková, Petr Pokorný, Mathieu Denoël, Heimo Emmerstorfer, Cvetka Lipovnik, and et al. 2023. "Pulling the Plug—Draining an Alpine Lake Failed to Eradicate Alien Minnows and Impacted Lower Trophic Levels" Water 15, no. 7: 1332. https://doi.org/10.3390/w15071332
APA StyleSchabetsberger, R., Jersabek, C. D., Maringer, A., Kreiner, D., Kaltenbrunner, M., Blažková, P., Pokorný, P., Denoël, M., Emmerstorfer, H., Lipovnik, C., & Wölger, H. (2023). Pulling the Plug—Draining an Alpine Lake Failed to Eradicate Alien Minnows and Impacted Lower Trophic Levels. Water, 15(7), 1332. https://doi.org/10.3390/w15071332