Managing Local Coastal Stressors to Reduce the Ecological Effects of Ocean Acidification and Warming

Abstract
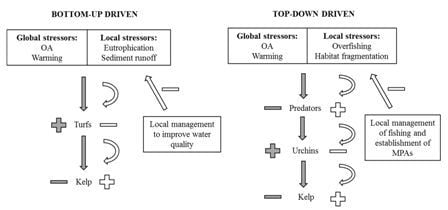
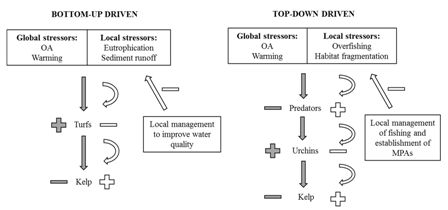
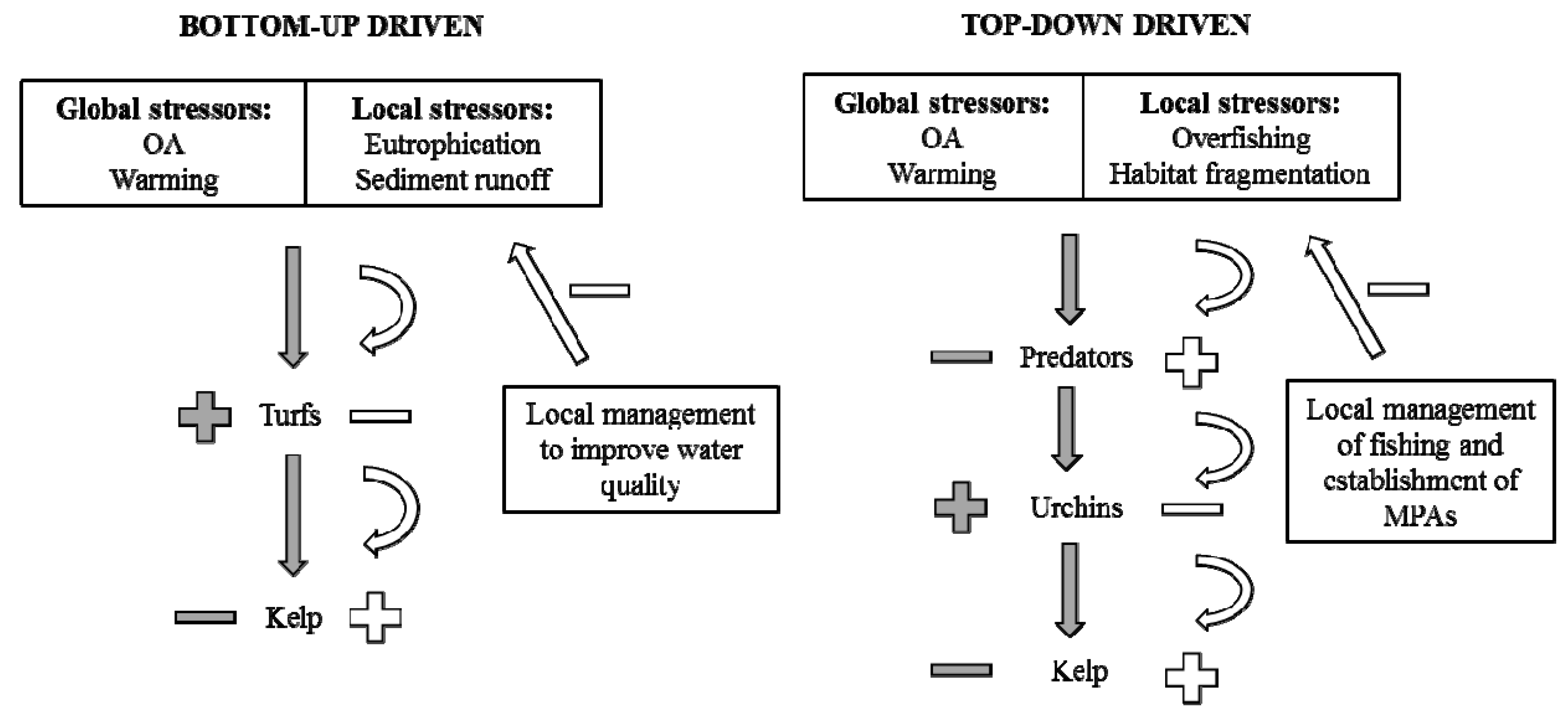
:
{kind=link}
{kind=link}
1. Introduction—Synergistic Effects between Long–Slow Global Stressors and Short–Rapid Local Stressors
2. An Example from Kelp Forests

3. Discussion—Acting on Rapidly Changing Local Stressors to Reduce the Effect of Global Stressors
Acknowledgments
Conflicts of Interest
References
- IPCC (Intergovernmental Panel on Climate Change), The Physical Science Basis; Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007.
- IPCC, Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation; A Special Report of Working Groups I and II of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2012.
- Harley, C.G.; Connell, S. Shifts in abiotic variables and consequences for diversity. In Marine Hard Bottom Communities; Wahl, M., Ed.; Springer: Berlin, Germany, 2009; pp. 257–268. [Google Scholar]
- Godbold, J.A.; Solan, M. Long-term effects of warming and ocean acidification are modified by seasonal variation in species responses and environmental conditions. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368. [Google Scholar] [CrossRef]
- Hönisch, B.; Ridgwell, A.; Schmidt, D.N.; Thomas, E.; Gibbs, S.J.; Sluijs, A.; Zeebe, R.; Kump, L.; Martindale, R.C.; Greene, S.E.; et al. The Geological record of ocean acidification. Science 2012, 335, 1058–1063. [Google Scholar] [CrossRef] [Green Version]
- Joshi, M.; Hawkins, E.; Sutton, R.; Lowe, J.; Frame, D. Projections of when temperature change will exceed 2 °C above pre-industrial levels. Nat. Clim. Change 2011, 1, 407–412. [Google Scholar] [CrossRef]
- Caldeira, K.; Wickett, M.E. Anthropogenic carbon and ocean pH. Nature 2003, 425, 365. [Google Scholar] [CrossRef]
- Raven, J.R.; Caldeira, K.; Elderfield, H.; Hoegh-Guldberg, O.; Liss, P.; Riebesell, U.; Shepherd, J.; Turley, C.; Watson, A.; Heap, R.; et al. Ocean Acidification Due to Increasing Atmospheric Carbon Dioxide; The Royal Society: London, UK, 2005. [Google Scholar]
- Doney, S.C.; Schimel, D.S. Carbon and climate system coupling on timescales from the precambrian to the anthropocene. Annu. Rev. Environ. Resour. 2007, 32, 31–66. [Google Scholar] [CrossRef]
- Piazzi, L.; Balata, D.; Ceccherelli, G.; Cinelli, F. Interactive effect of sedimentation and Caulerpa racemosa var. cylindracea invasion on macroalgal assemblages in the Mediterranean Sea. Estuar. Coast. Shelf Sci. 2005, 64, 467–474. [Google Scholar] [CrossRef]
- Worm, B.; Lotze, H.K. Effects of eutrophication, grazing, and algal blooms on rocky shores. Limnol. Oceanogr. 2006, 51, 569–579. [Google Scholar] [CrossRef] [Green Version]
- Jackson, J.B.C.; Kirby, M.X.; Berger, W.H.; Bjorndal, K.A.; Botsford, L.W.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.; Erlandson, J.; Estes, J.A.; et al. Historical overfishing and the recent collapse of coastal ecosystems. Science 2001, 293, 629–637. [Google Scholar] [CrossRef]
- Bulleri, F.; Chapman, M.G. The introduction of coastal infrastructure as a driver of change in marine environments. J. Appl. Ecol. 2010, 47, 26–35. [Google Scholar] [CrossRef]
- Brown, C.J.; Saunders, M.I.; Possingham, H.P.; Richardson, A.J. Managing for interactions between local and global stressors of ecosystems. PLoS ONE 2013, 8, e65765. [Google Scholar]
- Wilson, S.K.; Graham, N.A.J.; Pratchett, M.S.; Jones, G.P.; Polunin, N.C.V. Multiple disturbances and the global degradation of coral reefs: Are reef fishes at risk or resilient? Glob. Change Biol. 2006, 12, 2220–2234. [Google Scholar] [CrossRef]
- Hall, J.; Gruber, N.; Díaz, R.; Wilhelmsson, D. The impacts of multiple stressors: A complex web of challenges. In Valuing the Ocean; Noone, K., Sumaila, R., Díaz, R.J., Eds.; Stockholm Environment Institute: Stockholm, Sweden, 2013. [Google Scholar]
- Crain, C.M.; Kroeker, K.; Halpern, B.S. Interactive and cumulative effects of multiple human stressors in marine systems. Ecol. Lett. 2008, 11, 1304–1315. [Google Scholar] [CrossRef]
- Anthony, K.R.N.; Maynard, J.A.; Diaz-Pulido, G.; Mumby, P.J.; Marshall, P.A.; Cao, L.; Hoegh-Guldberg, O. Ocean acidification and warming will lower coral reef resilience. Glob. Change Biol. 2011, 17, 1798–1808. [Google Scholar] [CrossRef]
- Connell, S.D.; Russell, B.D. The direct effects of increasing CO2 and temperature on non-calcifying organisms: Increasing the potential for phase shifts in kelp forests. Proc. R. Soc. Biol. Sci. 2010, 277, 1409–1415. [Google Scholar] [CrossRef]
- Pettorelli, N. Climate change as a main driver of ecological research. J. Appl. Ecol. 2012, 49, 542–545. [Google Scholar] [CrossRef]
- Wooldridge, S.A. Water quality and coral bleaching thresholds: Formalising the linkage for the inshore reefs of the Great Barrier Reef, Australia. Mar. Pollut. Bull. 2009, 58, 745–751. [Google Scholar] [CrossRef]
- Connell, S.D.; Kroeker, K.J.; Fabricius, K.E.; Kline, D.I.; Russell, B.D. The other ocean acidification problem: CO2 as a resource among competitors for ecosystem dominance. Philos. Trans. R. Soc. B Biol. Sci. 2013, 20120442. [Google Scholar]
- Bulling, M.T.; Hicks, N.; Murray, L.; Paterson, D.M.; Raffaelli, D.; White, P.C.L.; Solan, M. Marine biodiversity-ecosystem functions under uncertain environmental futures. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2107–2116. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Zou, D.; Gao, K. Effects of elevated CO2 and phosphorus supply on growth, photosynthesis and nutrient uptake in the marine macroalga Gracilaria lemaneiformis (Rhodophyta). Bot. Mar. 2010, 53, 123–129. [Google Scholar]
- Hughes, T.P.; Rodrigues, M.J.; Bellwood, D.R.; Ceccarelli, D.; Hoegh-Guldberg, O.; McCook, L.; Moltschaniwskyj, N.; Pratchett, M.S.; Steneck, R.S.; Willis, B. Phase shifts, herbivory, and the resilience of coral reefs to climate change. Curr. Biol. 2007, 17, 360–365. [Google Scholar] [CrossRef]
- Gorman, D.; Connell, S.D. Recovering subtidal forests in human-dominated landscapes. J. Appl. Ecol. 2009, 46, 1258–1265. [Google Scholar] [CrossRef]
- Waycott, M.; Duarte, C.M.; Carruthers, T.J.B.; Orth, R.J.; Dennison, W.C.; Olyarnik, S.; Calladine, A.; Fourqurean, J.W.; Heck, K.L., Jr.; Hughes, A.R.; et al. Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sci. USA 2009, 106, 12377–12381. [Google Scholar] [CrossRef] [Green Version]
- Dayton, P.K. Ecology of kelp communities. Annu. Rev. Ecol. Syst. 1985, 16, 215–245. [Google Scholar]
- Goodsell, P.J.; Fowler-Walker, M.J.; Gillanders, B.M.; Connell, S.D. Variations in the configuration of algae in subtidal forests: Implications for invertebrate assemblages. Austral Ecol. 2004, 29, 350–357. [Google Scholar] [CrossRef]
- Smale, D.A.; Wernberg, T. Extreme climatic event drives range contraction of a habitat-forming species. Proc. R. Soc. B Biol. Sci. 2013, 280. [Google Scholar] [CrossRef]
- Dayton, P.K.; Tegner, M.J. Catastrophic storms, El-Nino, and patch stability in a southern California kelp community. Science 1984, 224, 283–285. [Google Scholar]
- Graham, M.H. Effects of local deforestation on the diversity and structure of Southern California giant kelp forest food webs. Ecosystems 2004, 7, 341–357. [Google Scholar] [CrossRef]
- Steneck, R.S.; Graham, M.H.; Bourque, B.J.; Corbett, D.; Erlandson, J.M.; Estes, J.A.; Tegner, M.J. Kelp forest ecosystems: Biodiversity, stability, resilience and future. Environ. Conserv. 2002, 29, 436–459. [Google Scholar]
- Kennelly, S.J. Inhibition of kelp-recruitment by turfing-algae and consequences for an Australasian kelp-community. J. Exp. Mar. Biol. Ecol. 1987, 112, 49–60. [Google Scholar] [CrossRef]
- Connell, S.D. The monopolization of understorey habitat by subtidal encrusting coralline algae: A test of the combined effects of canopy-mediated light and sedimentation. Mar. Biol. 2003, 142, 1065–1071. [Google Scholar]
- Irving, A.D.; Connell, S.D. Physical disturbance by kelp abrades erect algae from the understorey. Mar Ecol. Prog. Ser. 2006, 324, 127–137. [Google Scholar] [CrossRef]
- Airoldi, L. The effects of sedimentation on rocky coast assemblages. Oceanogr. Mar. Biol. 2003, 41, 161–236. [Google Scholar]
- Connell, S.D.; Russell, B.D.; Turner, D.J.; Shepherd, S.A.; Kildea, T.; Miller, D.; Airoldi, L.; Cheshire, A. Recovering a lost baseline: Missing kelp forests from a metropolitan coast. Mar. Ecol. Prog. Ser. 2008, 360, 63–72. [Google Scholar] [CrossRef]
- Falkenberg, L.J.; Russell, B.D.; Connell, S.D. Contrasting resource limitations of marine primary producers: Implications for competitive interactions under enriched CO2 and nutrient regimes. Oecologia 2013, 172, 575–583. [Google Scholar] [CrossRef]
- Hepburn, C.D.; Pritchard, D.W.; Cornwall, C.E.; McLeod, R.J.; Beardall, J.; Raven, J.A.; Hurd, C.L. Diversity of carbon use strategies in a kelp forest community: Implications for a high CO2 ocean. Glob. Change Biol. 2011, 17, 2488–2497. [Google Scholar] [CrossRef]
- Russell, B.D.; Thompson, J.-A.I.; Falkenberg, L.J.; Connell, S.D. Synergistic effects of climate change and local stressors: CO2 and nutrient-driven change in subtidal rocky habitats. Glob. Change Biol. 2009, 15, 2153–2162. [Google Scholar] [CrossRef]
- Falkenberg, L.J.; Connell, S.D.; Russell, B.D. Disrupting the effects of synergies between stressors: Improved water quality dampens the effects of future CO2 on a marine habitat. J. Appl. Ecol. 2013, 50, 51–58. [Google Scholar] [CrossRef]
- Ling, S.D.; Johnson, C.R.; Frusher, S.D.; Ridgway, K.R. Overfishing reduces resilience of kelp beds to climate-driven catastrophic phase shift. Proc. Natl. Acad. Sci. USA 2009, 106, 22341–22345. [Google Scholar] [CrossRef]
- Ling, S.D.; Johnson, C.R. Marine reserves reduce risk of climate-driven phase shift by reinstating size- and habitat-specific trophic interactions. Ecol. Appl. 2012, 22, 1232–1245. [Google Scholar] [CrossRef]
- Foster, M.S.; Schiel, D.R. Loss of predators and the collapse of southern California kelp forests (?): Alternatives, explanations and generalizations. J. Exp. Mar. Biol. Ecol. 2010, 393, 59–70. [Google Scholar] [CrossRef]
- Russell, B.D.; Connell, S.D. Origins and consequences of global and local stressors: Incorporating climatic and non-climatic phenomena that buffer or accelerate ecological change. Mar. Biol. 2012, 159, 2633–2639. [Google Scholar] [CrossRef]
- Boyd, P.W.; Hutchins, D.A. Understanding the responses of ocean biota to a complex matrix of cumulative anthropogenic change. Mar. Ecol. Prog. Ser. 2012, 470, 125–135. [Google Scholar] [CrossRef]
- Gonzalez, A.; Loreau, M. The causes and consequences of compensatory dynamics in ecological communities. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 393–414. [Google Scholar] [CrossRef]
- Ruckelshaus, M.; Doney, S.C.; Galindo, H.M.; Barry, J.P.; Chan, F.; Duffy, J.E.; English, C.A.; Gaines, S.D.; Grebmeier, J.M.; Hollowed, A.B.; et al. Securing ocean benefits for society in the face of climate change. Mar. Policy 2013, 40, 154–159. [Google Scholar] [CrossRef]
- Hughes, T.P.; Linares, C.; Dakos, V.; van de Leemput, I.A.; van Nes, E.H. Living dangerously on borrowed time during slow, unrecognized regime shifts. Trends Ecol. Evol. 2013, 28, 149–155. [Google Scholar] [CrossRef] [Green Version]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ghedini, G.; Russell, B.D.; Connell, S.D. Managing Local Coastal Stressors to Reduce the Ecological Effects of Ocean Acidification and Warming. Water 2013, 5, 1653-1661. https://doi.org/10.3390/w5041653
Ghedini G, Russell BD, Connell SD. Managing Local Coastal Stressors to Reduce the Ecological Effects of Ocean Acidification and Warming. Water. 2013; 5(4):1653-1661. https://doi.org/10.3390/w5041653
Chicago/Turabian StyleGhedini, Giulia, Bayden D. Russell, and Sean D. Connell. 2013. "Managing Local Coastal Stressors to Reduce the Ecological Effects of Ocean Acidification and Warming" Water 5, no. 4: 1653-1661. https://doi.org/10.3390/w5041653
APA StyleGhedini, G., Russell, B. D., & Connell, S. D. (2013). Managing Local Coastal Stressors to Reduce the Ecological Effects of Ocean Acidification and Warming. Water, 5(4), 1653-1661. https://doi.org/10.3390/w5041653