
Effects of Seasonal Thermal Stratification on the Functional Diversity and Composition of the Microbial Community in a Drinking Water Reservoir

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Sampling

2.2. Water Physical and Chemical Analysis
2.3. Total Cell Counts
2.4. Microbial Community Functional Diversity Determination
2.5. Roche 454 Pyrosequencing and Sequence Analysis
2.6. Statistical Analysis
3. Results
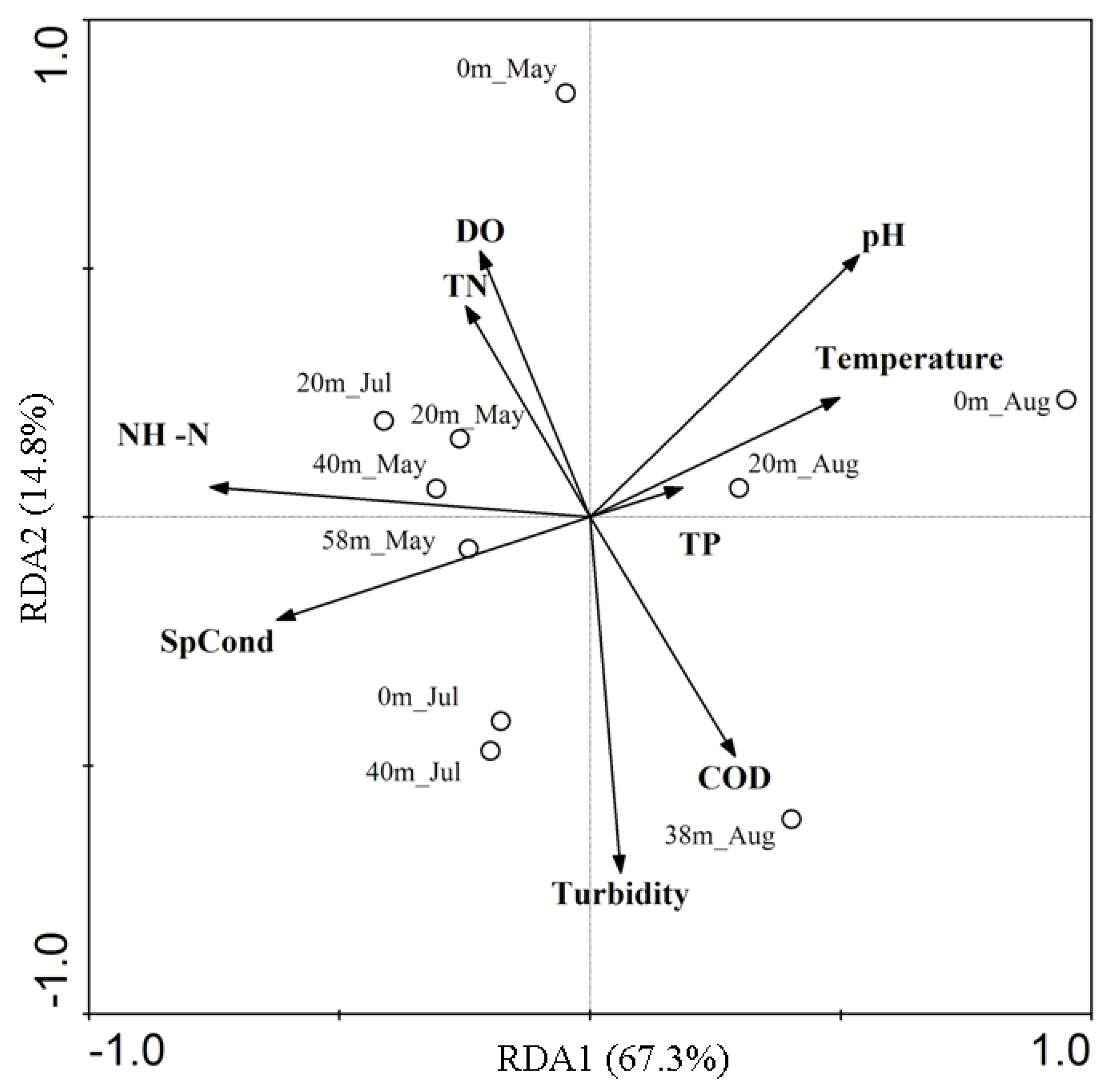
3.1. Physicochemical Properties of the Jinpen Reservoir

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water Layer | Mixing Layer (n = 12) | Epilimnion (n = 8) | Metalimnion (n = 6) | Hypolimnion (n = 8) |
|---|---|---|---|---|
| Temperature (°C) | 12.26 ± 3.82 BC | 21.88 ± 5.55 A | 14.27 ± 3.49 B | 8.50 ± 0.67 C |
| pH | 7.65 ± 0.42 A | 7.82 ± 0.74 A | 7.04 ± 0.40 AB | 6.78 ± 0.61 B |
| DO (mg·L−1) | 6.67 ± 2.58 AB | 8.31 ± 0.20 A | 6.0 ± 1.04 B | 0 ± 0 C |
| TN (mg·L−1) | 2.11 ± 0.13 A | 1.84 ± 0.03 B | 1.89 ± 0.11 BC | 1.99 ± 0.01 C |
| TP (mg·L−1) | 0.027 ± 0.006 A | 0.037 ± 0.001 A | 0.031 ± 0.010 A | 0.045 ± 0.013 B |
| CODMn (mg·L−1) | 3.40 ± 0.19 A | 4.03 ± 0.10 C | 3.77 ± 0.13 B | 3.96 ± 0.14 BC |
| NH4-N (mg·L−1) | 0.183 ± 0.001 AB | 0.126 ± 0.036 A | 0.116 ± 0.040 A | 0.218 ± 0.090 B |
| SpCond (ms·cm−1) | 172.0 ± 24.8 A | 144.7 ± 29.2 A | 150.5 ± 8.2 A | 172.7 ± 28.4 A |
| Turbidity (NTU) | 2.43 ± 0.84 A | 4.07 ± 1.75 A | 5.30 ± 0.33 A | 18.15 ± 9.69 B |
| Total cell count (1 × 105 cells·mL−1) | 55.4 ± 11.5 A | 69.6 ± 28.3 A | 56.8 ± 31.7 A | 63.9 ± 19.7 A |
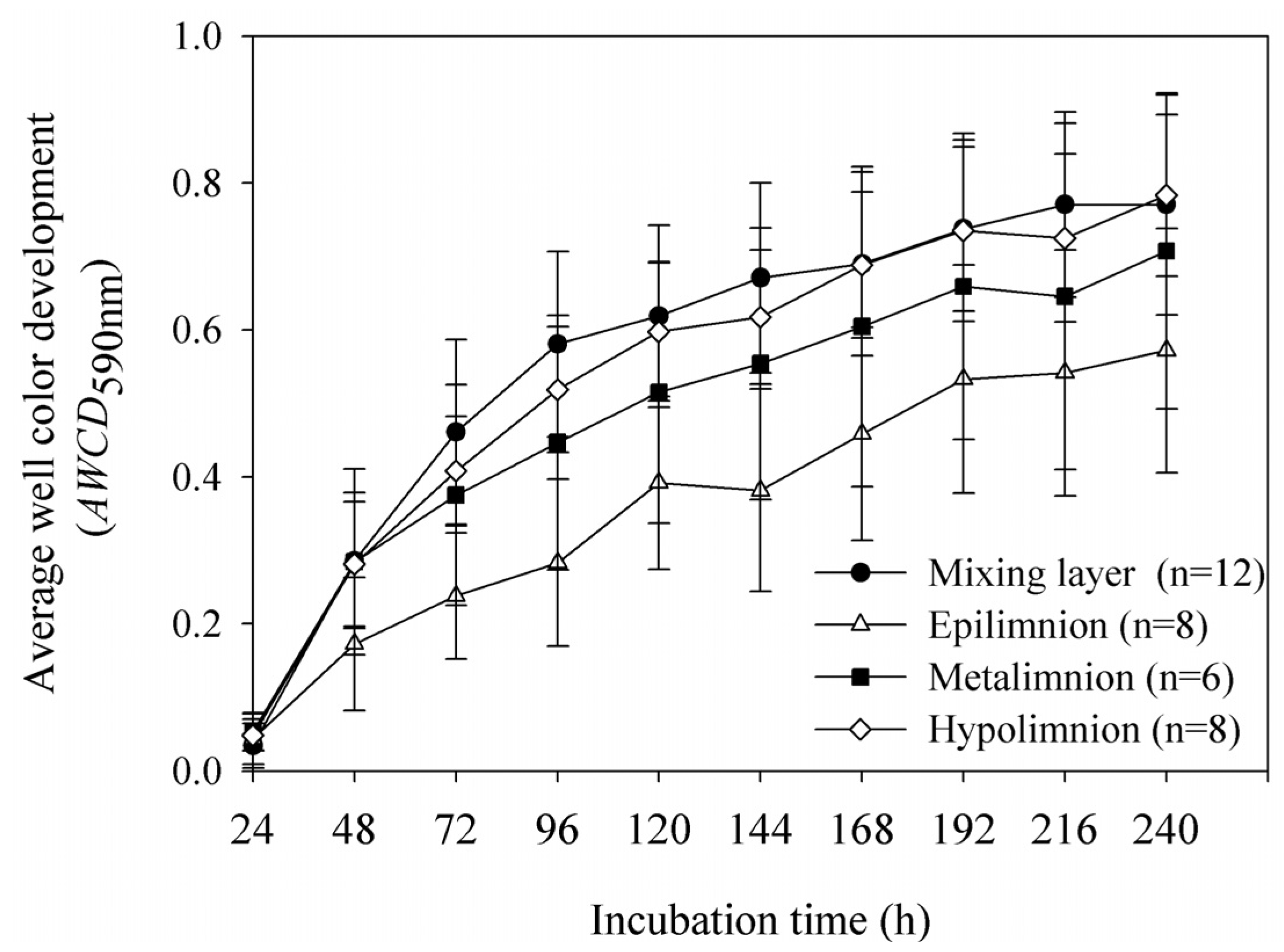
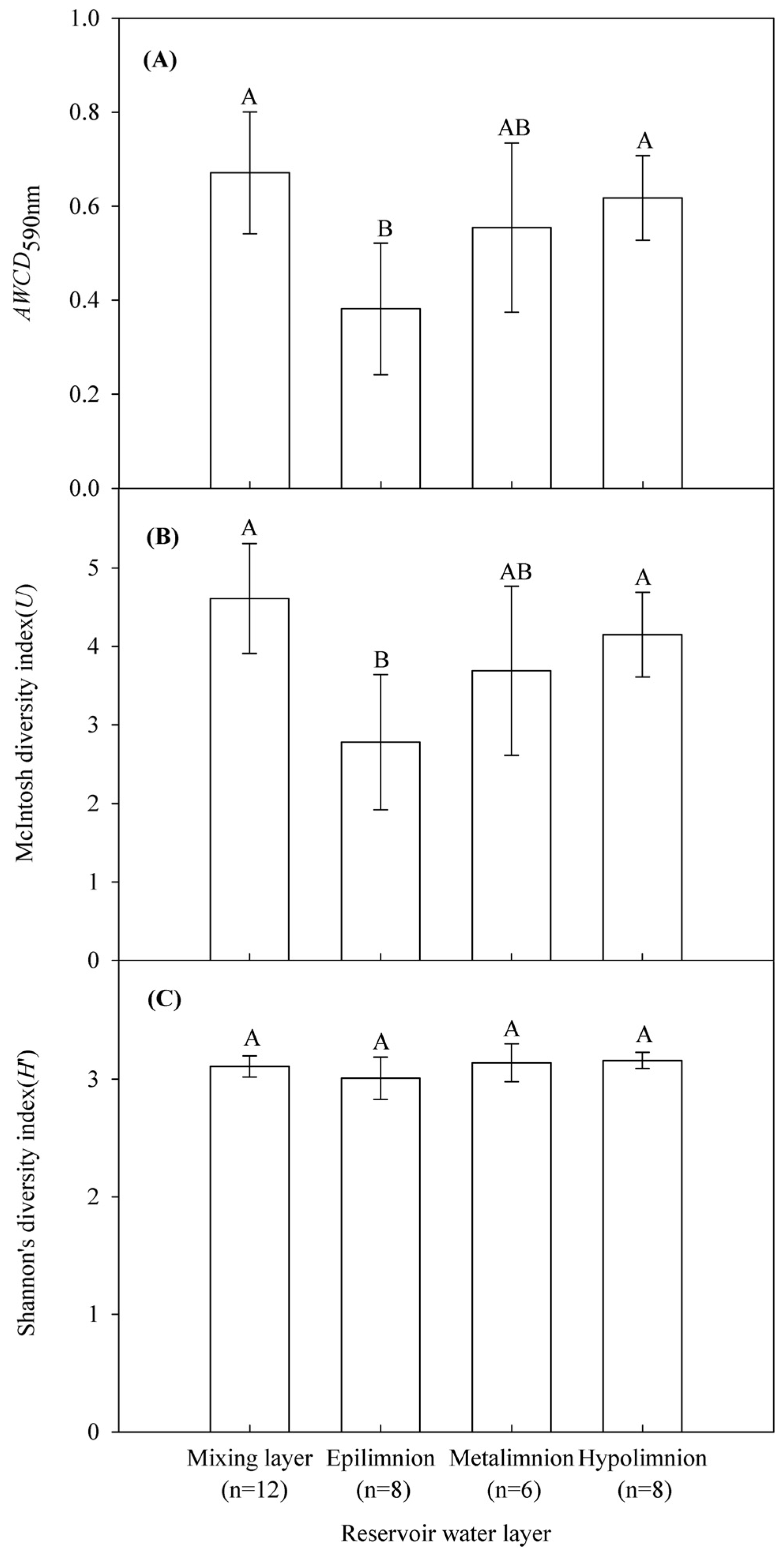
3.2. Microbial Community Functional Diversity


| Water Layer | Carbohydrates | Amino Acids | Esters | Alcohols | Amines | Carboxylic Acids |
|---|---|---|---|---|---|---|
| Mixing Layer (n = 12) | 0.69 ± 0.21 AB | 0.67 ± 0.11 A | 0.94 ± 0.16 A | 0.66 ± 0.10 A | 0.70 ± 0.21 A | 0.49 ± 0.15 A |
| Epilimnion (n = 8) | 0.46 ± 0.18 A | 0.31 ± 0.15 B | 0.56 ± 0.26 B | 0.29 ± 0.12 B | 0.43 ± 0.17 B | 0.28 ± 0.16 B |
| Metalimnion (n = 6) | 0.61 ± 0.27 AB | 0.54 ± 0.21 A | 0.84 ± 0.26 A | 0.50 ± 0.19 AC | 0.38 ± 0.19 B | 0.43 ± 0.17 AB |
| Hypolimnion (n = 8) | 0.81 ± 0.19 B | 0.55 ± 0.13 A | 0.95 ± 0.12 A | 0.47 ± 0.12 C | 0.59 ± 0.14 AB | 0.41 ± 0.11 AB |
| One Way ANOVA | 3.865 * | 9.899 ** | 6.873 ** | 13.097 ** | 5.480 ** | 3.017 * |

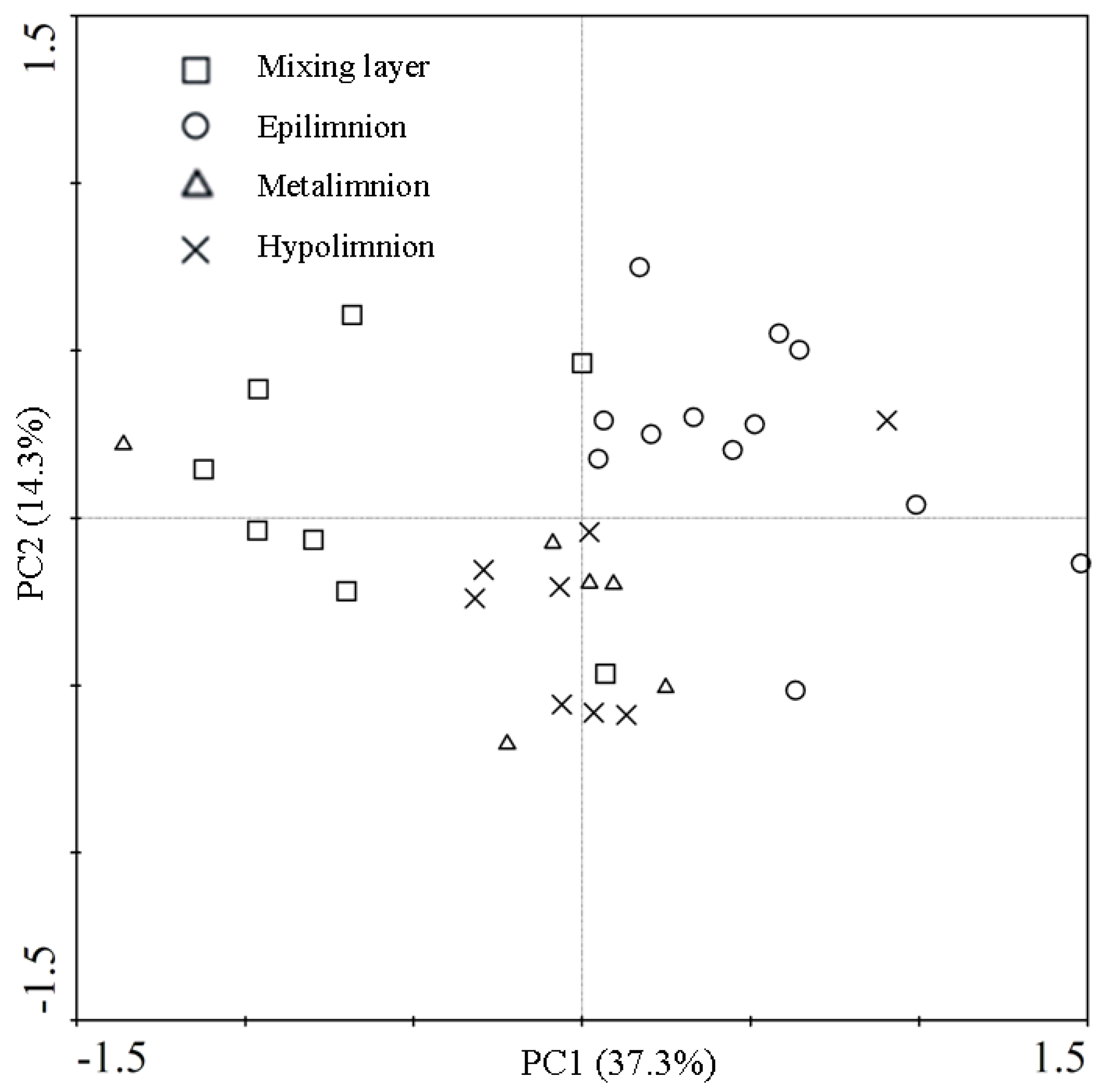
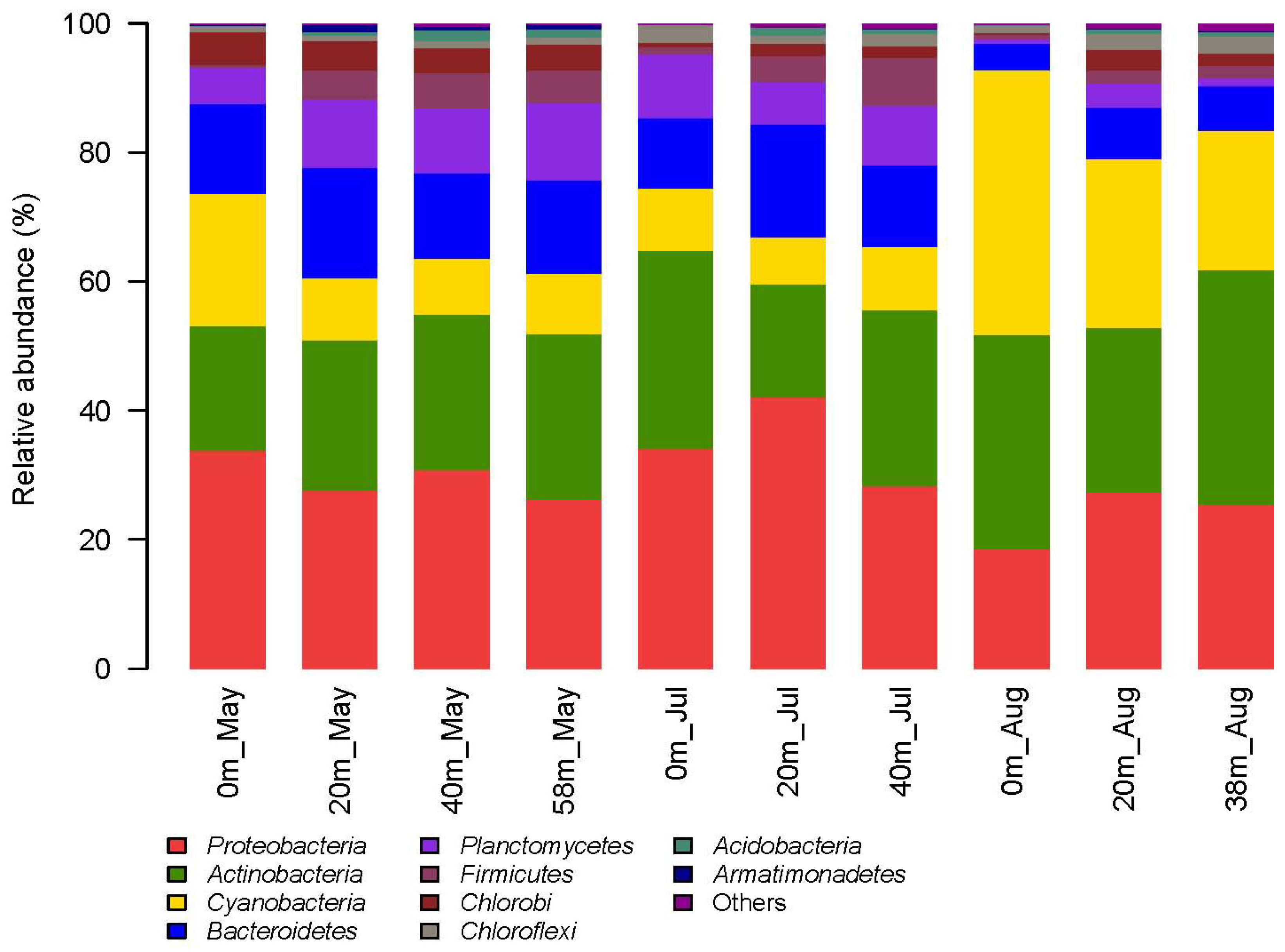
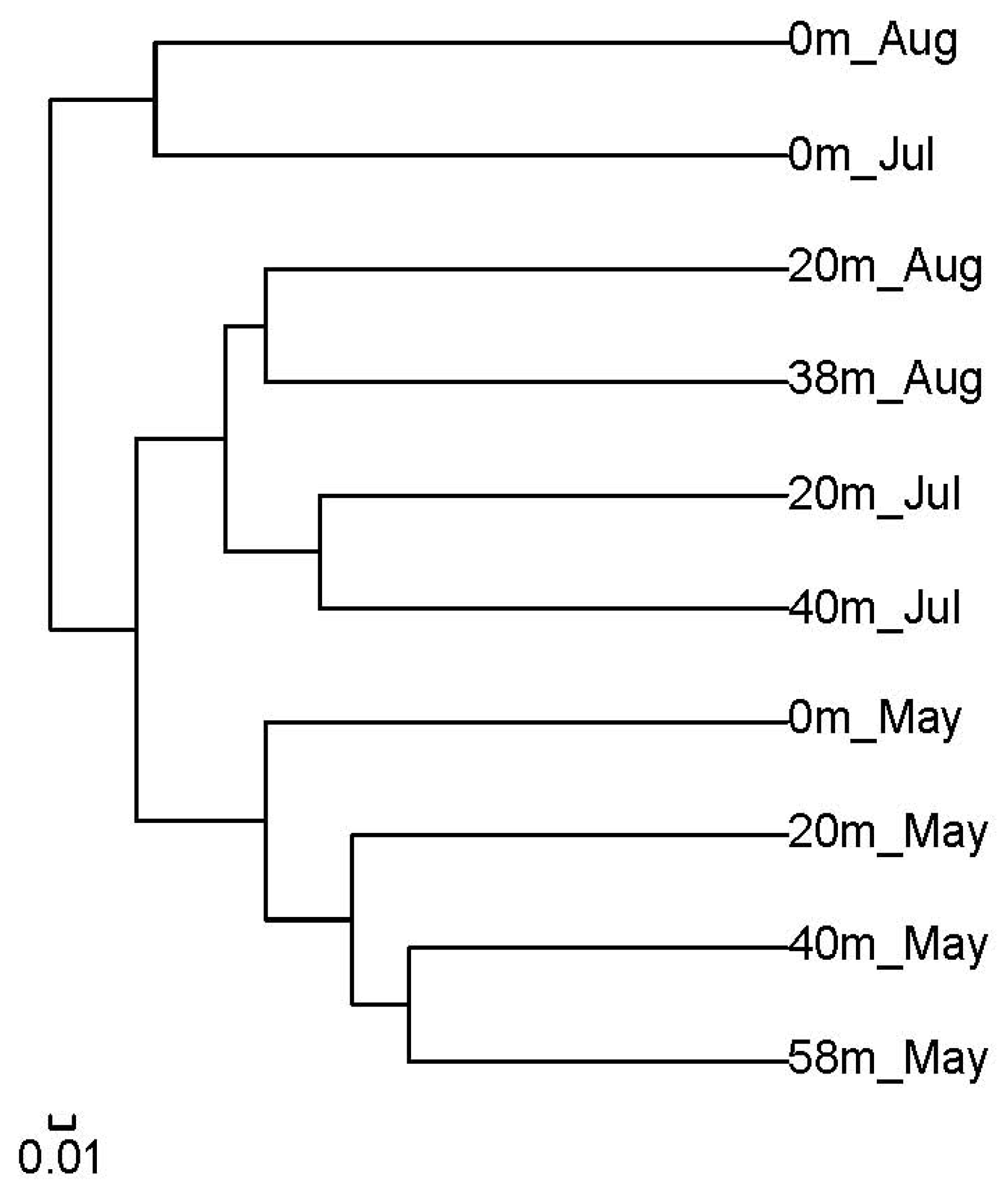
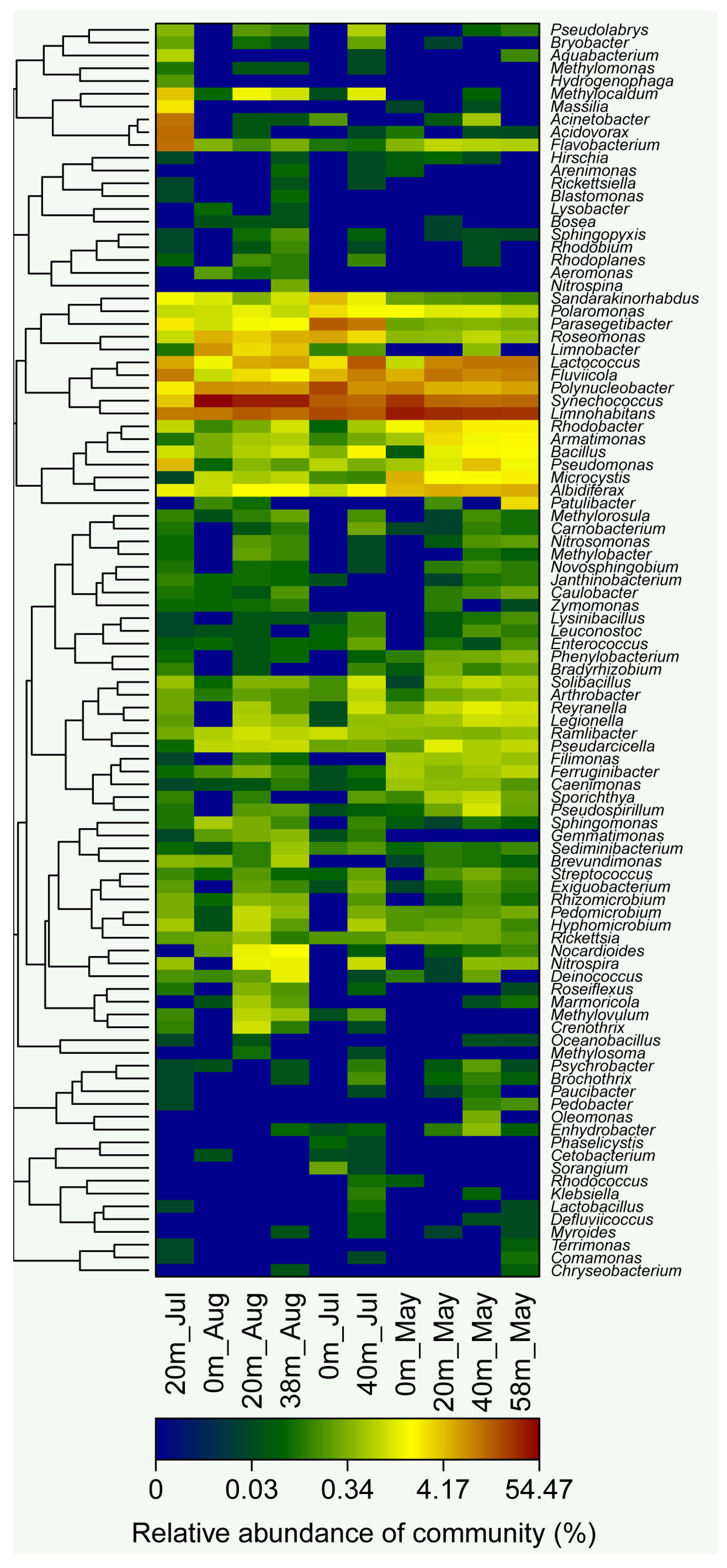
3.3. Pyrosequencing Results of Bacterial Community Composition

| Water Layer | Operational Taxonomic Units (OTUs) | Chao 1 Richness Estimate | Abundance-Based Coverage Estimators (ACE) | Shannon-Wiener Index (H') | Coverage |
|---|---|---|---|---|---|
| Mixing Layer (n = 4) | 302 ± 39 A | 375.5 ± 47.6 A | 369.8 ± 35.2 A | 4.11 ± 0.26 A | 0.99 |
| Epilimnion (n = 2) | 218 ± 13 B | 339.5 ± 6.4 A | 384.0 ± 56.6 A | 3.32 ± 0.24 B | 0.99 |
| Metalimnion (n = 2) | 356 ± 8 A | 497.5 ± 27.6 B | 477.5 ± 31.8 A | 4.25 ± 0.13 A | 0.99 |
| Hypolimnion (n = 2) | 364 ± 8 A | 483.0 ± 14.1 B | 469.0 ± 2.8 A | 4.13 ± 0.18 A | 0.99 |



4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Huang, T.; Li, X.; Rijnaarts, H.; Grotenhuis, T.; Ma, W.; Sun, X.; Xu, J. Effects of storm runoff on the thermal regime and water quality of a deep, stratified reservoir in a temperate monsoon zone, in Northwest China. Sci. Total Environ. 2014, 485, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Qian, X.; Han, B.-P.; Luo, L.-C.; Hamilton, D.P. Effects of local climate and hydrological conditions on the thermal regime of a reservoir at tropic of cancer, in Southern China. Water Res. 2012, 46, 2591–2604. [Google Scholar] [CrossRef] [PubMed]
- Elçi, Ş. Effects of thermal stratification and mixing on reservoir water quality. Limnology 2008, 9, 135–142. [Google Scholar]
- Becker, V.; Huszar, V.; Crossetti, L. Responses of phytoplankton functional groups to the mixing regime in a deep subtropical reservoir. Hydrobiologia 2009, 628, 137–151. [Google Scholar] [CrossRef]
- Yu, Z.; Yang, J.; Amalfitano, S.; Yu, X.; Liu, L. Effects of water stratification and mixing on microbial community structure in a subtropical deep reservoir. Sci. Rep. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- McGinnis, D.F.; Little, J.C. Predicting diffused-bubble oxygen transfer rate using the discrete-bubble model. Water Res. 2002, 36, 4627–4635. [Google Scholar] [CrossRef]
- Lawson, R.; Anderson, M.A. Stratification and mixing in lake elsinore, california: An assessment of axial flow pumps for improving water quality in a shallow eutrophic lake. Water Res. 2007, 41, 4457–4467. [Google Scholar] [CrossRef] [PubMed]
- Newton, R.J.; Jones, S.E.; Eiler, A.; McMahon, K.D.; Bertilsson, S. A guide to the natural history of freshwater lake bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 14–49. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, Y.; Kojima, H.; Oguri, K.; Kitazato, H.; Fukui, M. Vertical and temporal shifts in microbial communities in the water column and sediment of saline meromictic lake kaiike (Japan), as determined by a 16s rDNA-based analysis, and related to physicochemical gradients. Environ. Microbiol. 2004, 6, 622–637. [Google Scholar] [CrossRef] [PubMed]
- Röske, K.; Röske, I.; Uhlmann, D. Characterization of the bacterial population and chemistry in the bottom sediment of a laterally subdivided drinking water reservoir system. Limnol. Ecol. Manag. Inland Waters 2008, 38, 367–377. [Google Scholar] [CrossRef]
- Meuser, J.E.; Baxter, B.K.; Spear, J.R.; Peters, J.W.; Posewitz, M.C.; Boyd, E.S. Contrasting patterns of community assembly in the stratified water column of great salt lake, Utah. Microb. Ecol. 2013, 66, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Pommier, T.; Neal, P.R.; Gasol, J.M.; Coll, M.; Acinas, S.G.; Pedrós-Alió, C. Spatial patterns of bacterial richness and evenness in the nw mediterranean sea explored by pyrosequencing of the 16s rRNA. Aquat. Microb. Ecol. 2010, 61, 221–233. [Google Scholar] [CrossRef]
- Qian, P.-Y.; Wang, Y.; Lee, O.O.; Lau, S.C.K.; Yang, J.; Lafi, F.F.; Al-Suwailem, A.; Wong, T.Y.H. Vertical stratification of microbial communities in the red sea revealed by 16s rDNA pyrosequencing. ISME J. 2010, 5, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.-H.; Dobbs, F.C. Comparison of two kinds of biolog microplates (GN and ECO) in their ability to distinguish among aquatic microbial communities. J. Microbiol. Meth. 1999, 36, 203–213. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, B.; Zhang, W.; Hu, C.; An, S. Effects of plant and influent C:N:P ratio on microbial diversity in pilot-scale constructed wetlands. Ecol. Eng. 2010, 36, 441–449. [Google Scholar] [CrossRef]
- Rogers, B.; Tate, R. Temporal analysis of the soil microbial community along a toposequence in pineland soils. Soil Biol. Biochem. 2001, 33, 1389–1401. [Google Scholar] [CrossRef]
- Comeau, A.M.; Harding, T.; Galand, P.E.; Vincent, W.F.; Lovejoy, C. Vertical distribution of microbial communities in a perennially stratified arctic lake with saline, anoxic bottom waters. Sci. Rep. 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Boucher, D.; Jardillier, L.; Debroas, D. Succession of bacterial community composition over two consecutive years in two aquatic systems: A natural lake and a lake-reservoir. FEMS Microbiol. Ecol. 2006, 55, 79–97. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Li, G.; Wang, C.; Jing, Y.; Zhu, Y.; Zhang, S.; Liu, Y. Community dynamics of prokaryotic and eukaryotic microbes in an estuary reservoir. Sci. Rep. 2014, 4, 6966. [Google Scholar] [CrossRef] [PubMed]
- Pouliot, J.; Galand, P.E.; Lovejoy, C.; Vincent, W.F. Vertical structure of archaeal communities and the distribution of ammonia monooxygenase a gene variants in two meromictic high arctic lakes. Environ. Microbiol. 2009, 11, 687–699. [Google Scholar] [CrossRef] [PubMed]
- Kubo, K.; Kojima, H.; Fukui, M. Vertical distribution of major sulfate-reducing bacteria in a shallow eutrophic meromictic lake. Syst. Appl. Microbiol. 2014, 37, 510–519. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Li, X.; Zhang, M.; Huang, T.; Liu, W. Comparison of water-lifting aerator type for algae inhibition in stratified source water reservoirs. Ecol. Eng. 2014, 73, 624–634. [Google Scholar] [CrossRef]
- Huang, T.; Ma, Y.; Cong, H.; Tan, P. Application of the technology of water lifting and aeration on improving water quality in a deep canyon reservoir—A case study from Northern China. Desalination Water Treat. 2013, 52, 1636–1646. [Google Scholar] [CrossRef]
- Wilhelm, S.; Adrian, R. Impact of summer warming on the thermal characteristics of a polymictic lake and consequences for oxygen, nutrients and phytoplankton. Freshw. Biol. 2008, 53, 226–237. [Google Scholar] [CrossRef]
- Shade, A.; Jones, S.E.; McMahon, K.D. The influence of habitat heterogeneity on freshwater bacterial community composition and dynamics. Environ. Microbiol. 2008, 10, 1057–1067. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Guo, Q.; Huang, T.; Tan, P. Response characteristics of water quality to seasonal thermal stratification in Jinpen reservoir along the heihe river, Xi’an city in China. J. Hydraul. Eng. 2013, 44, 406–415. (In Chinese) [Google Scholar]
- Li, X.; Huang, T.; Ma, W.; Sun, X.; Zhang, H. Effects of rainfall patterns on water quality in a stratified reservoir subject to eutrophication: Implications for management. Sci. Total Environ. 2015, 521, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Wen, G.; Ma, J.; Huang, T.-L.; Egli, T. Using coagulation to restrict microbial re-growth in tap water by phosphate limitation in water treatment. J. Hazar. Mater. 2014, 280, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Hammes, F.; Egli, T. Cytometric methods for measuring bacteria in water: Advantages, pitfalls and applications. Anal. Bioanal. Chem. 2010, 397, 1083–1095. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Huang, T.; Liu, T. Sediment enzyme activities and microbial community diversity in an oligotrophic drinking water reservoir, Eastern China. PloS ONE 2013, 8, e78571. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Huang, T.; Chen, S.; Guo, L.; Liu, T.; Yang, X. Microbial community functional diversity and enzymatic activity in the sediments of drinking water reservoirs, Northwest China. Desalination Water Treat. 2014, 52, 1608–1614. [Google Scholar] [CrossRef]
- Zak, J.C.; Willig, M.R.; Moorhead, D.L.; Wildman, H.G. Functional diversity of microbial communities: A quantitative approach. Soil Biol. Biochem. 1994, 26, 1101–1108. [Google Scholar] [CrossRef]
- Zhu, D.; Tanabe, S.-H.; Yang, C.; Zhang, W.; Sun, J. Bacterial community composition of South China sea sediments through pyrosequencing-based analysis of 16s rRNA genes. PloS ONE 2013, 8, e78501. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I. Qiime allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D. A high-throughput DNA sequence aligner for microbial ecology studies. PLoS ONE 2009, 4, e8230. [Google Scholar] [CrossRef] [PubMed]
- Hamady, M.; Walker, J.J.; Harris, J.K.; Gold, N.J.; Knight, R. Error-correcting barcoded primers for pyrosequencing hundreds of samples in multiplex. Nat. Methods 2008, 5, 235–237. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.R.; Chai, B.; Farris, R.J.; Wang, Q.; Kulam-Syed-Mohideen, A.; McGarrell, D.M.; Bandela, A.; Cardenas, E.; Garrity, G.M.; Tiedje, J.M. The ribosomal database project (RDP-II): Introducing myrdp space and quality controlled public data. Nucleic Acids Res. 2007, 35, D169–D172. [Google Scholar] [CrossRef] [PubMed]
- Crawford, P.A.; Crowley, J.R.; Sambandam, N.; Muegge, B.D.; Costello, E.K.; Hamady, M.; Knight, R.; Gordon, J.I. Regulation of myocardial ketone body metabolism by the gut microbiota during nutrient deprivation. Proceed. Natl. Acad. Sci. 2009, 106, 11276–11281. [Google Scholar] [CrossRef] [PubMed]
- Chao, A.; Bunge, J. Estimating the number of species in a stochastic abundance model. Biometrics 2002, 58, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.-T.; Peng, X.; Deng, G.-H.; Sheng, H.-F.; Wang, Y.; Zhou, H.-W.; Tam, N.F.-Y. Illumina sequencing of 16s rRNA tag revealed spatial variations of bacterial communities in a mangrove wetland. Microb. Ecol. 2013, 66, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. Unifrac: An effective distance metric for microbial community comparison. ISME J. 2011, 5, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Stevens, M.H.H.; Oksanen, M.J.; Suggests, M. The Vegan Package. Available online: http://www.researchgate.net/profile/Gavin_Simpson/publication/228339454_The_vegan_package/links/0912f50be86bc29a7f000000.pdf (accessed on 18 February 2015).
- Zhang, H.-H.; Huang, T.-L.; Chen, S.-N.; Yang, X.; Lv, K.; Sekar, R. Abundance and diversity of bacteria in oxygen minimum drinking water reservoir sediments studied by quantitative PCR and pyrosequencing. Microbol. Ecol. 2015, 69, 618–629. [Google Scholar] [CrossRef] [PubMed]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Schoener, T.W. Resource partitioning in ecological communities. Science 1974, 185, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Valdespino-Castillo, P.M.; Merino-Ibarra, M.; Jiménez-Contreras, J.; Castillo-Sandoval, F.S.; Ramírez-Zierold, J.A. Community metabolism in a deep (stratified) tropical reservoir during a period of high water-level fluctuations. Environ. Monit. Assess. 2014, 186, 6505–6520. [Google Scholar] [CrossRef] [PubMed]
- Bauer, M.; Kube, M.; Teeling, H.; Richter, M.; Lombardot, T.; Allers, E.; Würdemann, C.A.; Quast, C.; Kuhl, H.; Knaust, F. Whole genome analysis of the marine bacteroidetes “gramella forsetii” reveals adaptations to degradation of polymeric organic matter. Environ. Microbiol. 2006, 8, 2201–2213. [Google Scholar] [CrossRef] [PubMed]
- Grossart, H.P.; Levold, F.; Allgaier, M.; Simon, M.; Brinkhoff, T. Marine diatom species harbour distinct bacterial communities. Environ. Microbiol. 2005, 7, 860–873. [Google Scholar] [CrossRef] [PubMed]
- Hupfer, M.; Lewandowski, J. Oxygen controls the phosphorus release from lake sediments—A long-lasting paradigm in limnology. Int. Rev. Hydrobiol. 2008, 93, 415–432. [Google Scholar] [CrossRef]
- Song, L.-Y.; Wang, Y.-Q. Investigation of microbial community structure of a shallow lake after one season copper sulfate algaecide treatment. Microbiol. Res. 2015, 170, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Hiorns, W.D.; Methe, B.A.; Nierzwicki-Bauer, S.A.; Zehr, J.P. Bacterial diversity in adirondack mountain lakes as revealed by 16s rRNA gene sequences. Appl. Environ. Microbiol. 1997, 63, 2957–2960. [Google Scholar] [PubMed]
- Pernthaler, J.; Glöckner, F.-O.; Unterholzner, S.; Alfreider, A.; Psenner, R.; Amann, R. Seasonal community and population dynamics of pelagic bacteria and archaea in a high mountain lake. Appl. Environ. Microbiol. 1998, 64, 4299–4306. [Google Scholar] [PubMed]
- Eiler, A.; Bertilsson, S. Composition of freshwater bacterial communities associated with cyanobacterial blooms in four swedish lakes. Environ. Microbiol. 2004, 6, 1228–1243. [Google Scholar] [CrossRef] [PubMed]
- Kasalický, V.; Jezbera, J.; Hahn, M.W.; Šimek, K. The diversity of the limnohabitans genus, an important group of freshwater bacterioplankton, by characterization of 35 isolated strains. PLoS ONE 2013, 8, e58209. [Google Scholar] [CrossRef] [PubMed]
- Hahn, M.W.; Pöckl, M.; Wu, Q.L. Low intraspecific diversity in a polynucleobacter subcluster population numerically dominating bacterioplankton of a freshwater pond. Appl. Environ. Microbiol. 2005, 71, 4539–4547. [Google Scholar] [CrossRef] [PubMed]
- Bickel, S.L.; Tang, K.W. Carbon substrate usage by zooplankton-associated bacteria, phytoplankton-associated bacteria, and free-living bacteria under aerobic and anaerobic conditions. Mar. Biol. 2014, 161, 2233–2242. [Google Scholar] [CrossRef]
- Garland, J.L.; Mills, A.L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Appl. Environ. Microbiol. 1991, 57, 2351–2359. [Google Scholar] [PubMed]
- Kersters, I.; Van Vooren, L.; Verschuere, L.; Vauterin, L.; Wouters, A.; Mergaert, J.; Swings, J.; Verstraete, W. Utility of the biolog system for the characterization of heterotrophic microbial communities. Syst. Appl. Microbiol. 1997, 20, 439–447. [Google Scholar] [CrossRef]
- Garland, J.L. Analytical approaches to the characterization of samples of microbial communities using patterns of potential C source utilization. Soil Biol. Biochem. 1996, 28, 213–221. [Google Scholar] [CrossRef]
- Fredrickson, J.K.; Balkwill, D.L.; Zachara, J.M.; Li, S.-M.W.; Brockman, F.J.; Simmons, M.A. Physiological diversity and distributions of heterotrophic bacteria in deep cretaceous sediments of the atlantic coastal plain. Appl. Environ. Microbiol. 1991, 57, 402–411. [Google Scholar] [PubMed]
- Fuller, M.E.; Scow, K.M.; Lau, S.; Ferris, H. Trichloroethylene (TCE) and toluene effects on the structure and function of the soil community. Soil Biol. Biochem. 1997, 29, 75–89. [Google Scholar] [CrossRef]
- Bossio, D.A.; Scow, K.M. Impact of carbon and flooding on the metabolic diversity of microbial communities in soils. Appl. Environ. Microbiol. 1995, 61, 4043–4050. [Google Scholar] [PubMed]
- Haack, S.K.; Garchow, H.; Klug, M.J.; Forney, L.J. Analysis of factors affecting the accuracy, reproducibility, and interpretation of microbial community carbon source utilization patterns. Appl. Environ. Microbiol. 1995, 61, 1458–1468. [Google Scholar] [PubMed]
- Sun, W.; Cupples, A.M. Diversity of five anaerobic toluene-degrading microbial communities investigated using stable isotope probing. Appl. Environ. Microbiol. 2012, 78, 972–980. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Huang, T.; Zhang, H. Effects of Seasonal Thermal Stratification on the Functional Diversity and Composition of the Microbial Community in a Drinking Water Reservoir. Water 2015, 7, 5525-5546. https://doi.org/10.3390/w7105525
Yang X, Huang T, Zhang H. Effects of Seasonal Thermal Stratification on the Functional Diversity and Composition of the Microbial Community in a Drinking Water Reservoir. Water. 2015; 7(10):5525-5546. https://doi.org/10.3390/w7105525
Chicago/Turabian StyleYang, Xiao, Tinglin Huang, and Haihan Zhang. 2015. "Effects of Seasonal Thermal Stratification on the Functional Diversity and Composition of the Microbial Community in a Drinking Water Reservoir" Water 7, no. 10: 5525-5546. https://doi.org/10.3390/w7105525
APA StyleYang, X., Huang, T., & Zhang, H. (2015). Effects of Seasonal Thermal Stratification on the Functional Diversity and Composition of the Microbial Community in a Drinking Water Reservoir. Water, 7(10), 5525-5546. https://doi.org/10.3390/w7105525