
Integrated Mosquito Management in Experimental Constructed Wetlands: Efficacy of Small-Stature Macrophytes and Fluctuating Hydroperiod


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Vegetation
2.3. Water Quality Monitoring
2.4. Mosquitoes and Invertebrates
3. Results
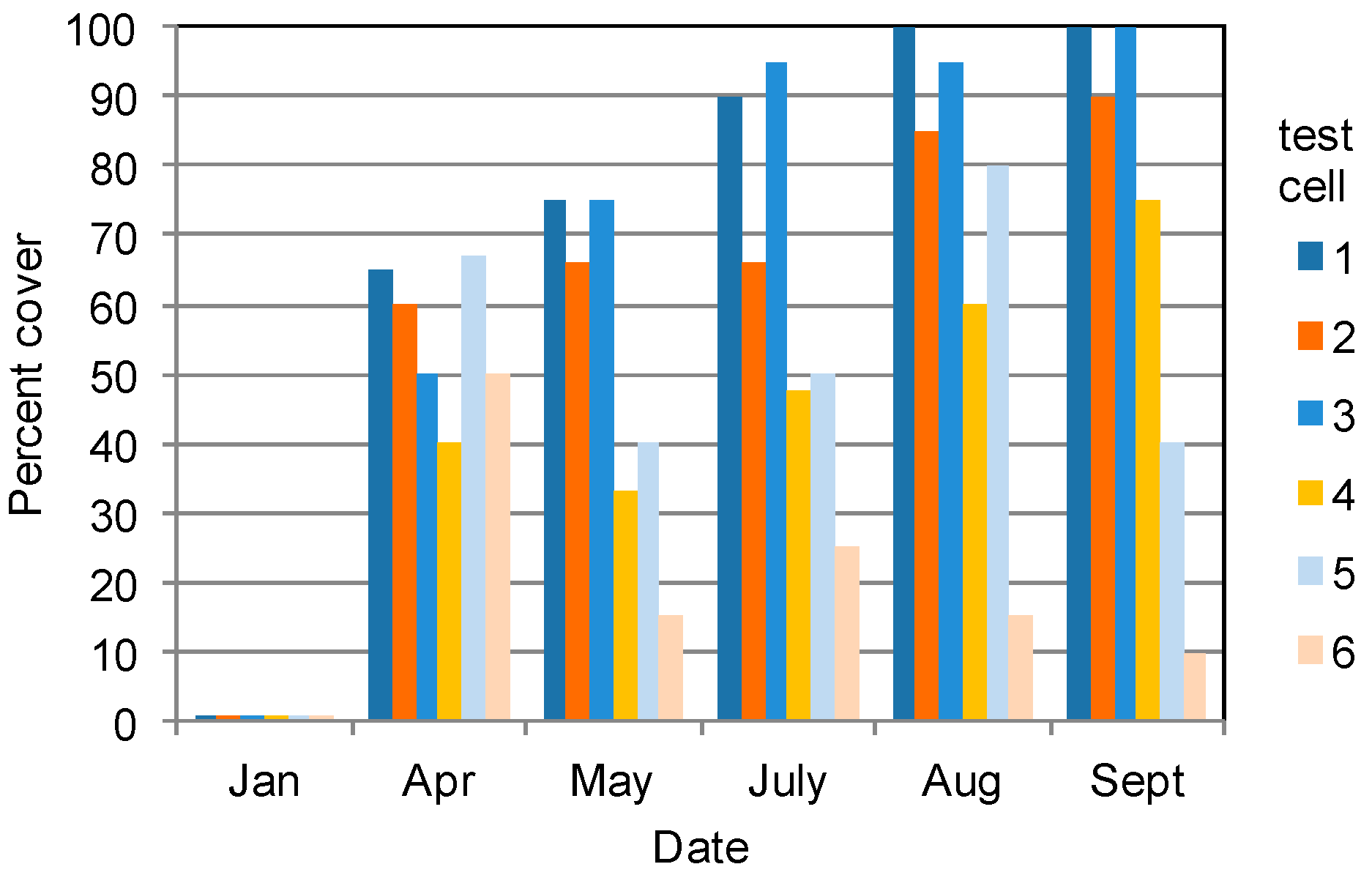
3.1. Vegetation
3.2. Water Quality
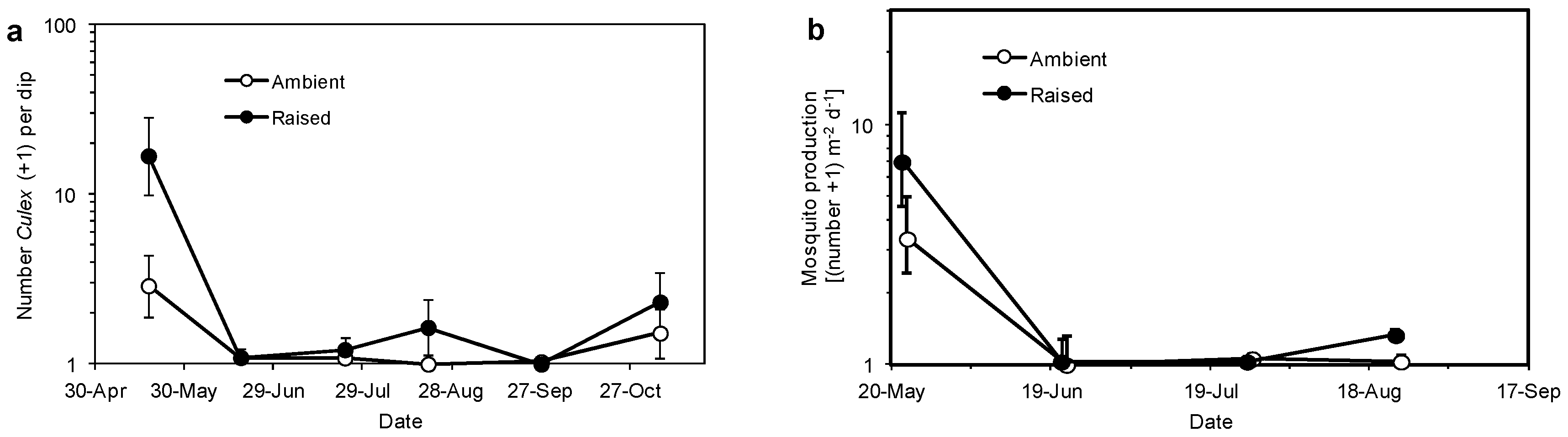
3.3. Mosquitoes and Invertebrates
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Brix, H. Do macrophytes play a role in constructed wetland treatment wetlands? Water Sci. Technol. 1997, 35, 11–17. [Google Scholar] [CrossRef]
- Stottmeister, U.; Wießner, A.; Kuschk, P.; Kappelmeyer, U.; Kästner, M.; Bederski, O.; Müller, R.A.; Moormann, H. Effects of plants and microorganisms in constructed wetlands for wastewater treatment. Biotechnol. Adv. 2003, 22, 93–117. [Google Scholar] [CrossRef] [PubMed]
- Kadlec, R.H.; Wallace, S.D. Treatment Wetlands, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Walton, W.E. Multipurpose constructed treatment wetlands in the arid southwestern United States: Are the benefits worth the risks? In Treatment Wetlands for Water Quality Improvement, Proceedings of the Quebec 2000 Conference, Quebec City, QC, Canada, 6–12 August 2000; Pries, J., Ed.; Selected Papers. CH2M Hill Canada Limited, Pandora Press: Waterloo, ON, Canada, 2002; pp. 115–123. [Google Scholar]
- Walton, W.E. Managing Mosquitoes in Surface-Flow Constructed Treatment Wetlands; Publication Number 8117; Division of Agriculture and Natural Resources, University of California: Davis, CA, USA, 2003; Available online: http://anrcatalog.ucdavis.edu/pdf/8117.pdf (accessed on 20 August 2016).
- Walton, W.E. Design and management of free water surface constructed wetlands to minimize mosquito production. Wetl. Ecol. Manag. 2012, 20, 173–195. [Google Scholar] [CrossRef]
- Walton, W.E.; Popko, D.A.; Van Dam, A.R.; Merrill, A.; Lythgoe, J.; Hess, B. Width of planting beds for emergent vegetation influences mosquito production from a constructed wetland in California (USA). Ecol. Eng. 2012, 42, 150–159. [Google Scholar] [CrossRef]
- Walton, W.E.; Popko, D.A.; Van Dam, A.R.; Merrill, A. Distribution of Culex species in vegetation bands of a constructed wetland undergoing integrated mosquito management. J. Am. Mosq. Control Assoc. 2013, 29, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Berkelhamer, R.C.; Bradley, T.J. Mosquito larval development in container habitats: The role of rotting Scirpus californicus. J. Am. Mosq. Control Assoc. 1989, 5, 258–260. [Google Scholar] [PubMed]
- Walton, W.E.; Jiannino, J.A. Vegetation management to stimulate denitrification increases mosquito abundance in multipurpose constructed treatment wetlands. J. Am. Mosq. Control Assoc. 2005, 21, 22–27. [Google Scholar] [CrossRef]
- Smith, G.C.; Kukkonen, I. A new lectotype for Scirpus maritimus (Cyperaceae). Taxon 1999, 48, 355–357. [Google Scholar] [CrossRef]
- United States Department of Agriculture (USDA); National Resources Conservation Service (NRCS). The PLANTS Database; National Plant Data Team: Greensboro, NC, USA, 2011. Available online: http://plants.usda.gov/java/profile?symbol=SCMA8 (accessed on 20 August 2016).
- Hoag, C. Plant Guide. Alkali Bulrush Schoenoplectus maritimus (L.) Lye; USDA-Natural Resources Conservation Service, Idaho Plant Materials Center: Aberdeen, ID, USA, 2000. Available online: http://plants.usda.gov/plantguide/pdf/pg_scma8.pdf (accessed on 20 August 2016).
- Kantrud, H.A. The Alkali (Scirpus maritimus L.) and Saltmarsh (S. robustus Pursh) Bulrushes: A Literature Review. Information and Technology Report 6; National Biological Service, Northern Prairie Wildlife Research Center: Jamestown, ND, USA, 1996. Available online: https://web.archive.org/web/20130902225308/http://www.npwrc.usgs.gov/resource/plants/bulrush (accessed on 24 August 2016). [Google Scholar]
- Miller, M.R.; Burns, E.G.; Wickland, B.E.; Eadie, J.M. Diet and body mass of wintering ducks in adjacent brackish and freshwater habitats. Waterbirds 2009, 32, 374–378. [Google Scholar] [CrossRef]
- Mills, W.R.; Bradford, S.M.; Rigby, M.; Wehner, M.P. Groundwater recharge at the Orange County Water District. In Wastewater Reclamation and Reuse; Asano, T., Ed.; Technomic Publishing Company: Lancaster, PA, USA, 1998; pp. 1105–1141. [Google Scholar]
- Skrenter, R.G. Instrumentation Handbook for Water and Wastewater Treatment Plants; ATSM D 5242-92; Lewis Publishers: Chelsea, MI, USA, 1988. [Google Scholar]
- Popko, D.A.; Walton, W.E. Small-stature emergent macrophytes and crepuscular sprinkler disturbance reduce mosquito abundance in wetland mesocosms. J. Vector Ecol. 2013, 38, 379–389. [Google Scholar] [CrossRef] [PubMed]
- American Public Health Association (APHA). Standard Methods for the Examination of Water and Wastewater, 19th ed.; American Public Health Association: Washington, DC, USA, 1995. [Google Scholar]
- Meyer, R.P.; Durso, S.L. Identification of the Mosquitoes of California; Mosquito and Vector Control Association of California: Sacramento, CA, USA, 1998. [Google Scholar]
- Merritt, R.W.; Cummins, K.W.; Berg, M.B. An Introduction to the Aquatic Insects of North America, 4th ed.; Kendall Hunt Publishing Co.: Dubuque, IA, USA, 2008. [Google Scholar]
- Walton, W.E.; Workman, P.D.; Keiper, J.B. An inexpensive collapsible pyramidal emergence trap for the assessment of wetland insect populations. In Proceedings of the 67th Annual Conference of the Mosquito and Vector Control Association of California, Anaheim, CA, USA, 24–27 January 1999; Mosquito and Vector Control Association of California: Sacramento, CA, USA, 1999; pp. 15–17. [Google Scholar]
- Duguma, D.; Walton, W.E. Effects of nutrients on mosquitoes and an emergent macrophyte, Schoenoplectus maritimus, for use in treatment wetlands. J. Vector Ecol. 2014, 39, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.J.; Rafferty, P.S. Clonal variation in response to salinity and flooding stress in four marsh macrophytes of the northern gulf of Mexico, USA. Environ. Exp. Bot. 2006, 56, 301–313. [Google Scholar] [CrossRef]
- Noack, T.J.; Pierce, R.M. The swamps of Cactus: Year 1 lessons learned treating high-strength wastewater with a constructed wetland. In Proceedings of the Texas Water Conference, San Antonio, TX, USA, 10–13 April 2012.
- Deegan, B.; Harrington, T.J.; Dundon, P. Effects of salinity and inundation regime on growth and distribution of Schoenoplectus triqueter. Aquat. Bot. 2005, 81, 199–211. [Google Scholar] [CrossRef]
- Clevering, O.A.; van Gulik, W.M.G. Restoration of Scirpus lacustris and Scirpus maritimus stands in a former tidal area. Aquat. Bot. 1997, 55, 229–246. [Google Scholar] [CrossRef]
- Mall, R.E. Soil-Water-Salt Relationships of Waterfowl Food Plants in the Suisun Marsh of California; Wildlife Bulletin Number 1; California Department of Fish and Game: Sacramento, CA, USA, 1969.
- Lieffers, V.J.; Shay, J.M. The effects of water level on the growth and reproduction of Scirpus maritimus var. paludosus. Can. J. Bot. 1981, 59, 118–121. [Google Scholar] [CrossRef]
- Shay, J.M.; Shay, C.T. Prairie marshes in western Canada, with specific reference to the ecology of five emergent macrophytes. Can. J. Bot. 1986, 64, 443–454. [Google Scholar] [CrossRef]
- Crawford, R.M.M. Oxygen availability as an ecological limit to plant distribution. Adv. Ecol. Res. 1992, 23, 93–171. [Google Scholar]
- Clevering, O.A.; Hundscheid, M.P. Plastic and non-plastic variation in growth of newly established clones of Scirpus (Bolboschoenus) maritimus L. grown at different water depths. Aquat. Bot. 1998, 62, 1–17. [Google Scholar] [CrossRef]
- Coops, H.; van den Brink, F.W.B.; van der Velde, G. Growth and morphological responses of four helophyte species in an experimental water-depth gradient. Aquat. Bot. 1996, 54, 11–24. [Google Scholar] [CrossRef]
- Lillebø, A.I.; Pardal, M.A.; Neto, J.M.; Marques, J.C. Salinity as the major factor affecting Scirpus maritimus annual dynamics. Evidence from field data and greenhouse experiment. Aquat. Bot. 2003, 77, 111–120. [Google Scholar] [CrossRef]
- Clevering, O.A.; Blom, C.W.P.M.; van Vierssen, W. Growth and morphology of Scirpus lacustris and S. maritimus seedlings as affected by water level and light availability. Funct. Ecol. 1996, 10, 289–296. [Google Scholar] [CrossRef]
- Keiper, J.B.; Jiannino, J.A.; Beehler, J.; Walton, W.E. Distribution and abundance of Culicidae and Chironomidae (Diptera) following storm damage in a southern California constructed wetlands. In Proceedings of the 67th Annual Conference of the Mosquito and Vector Control Association of California, Anaheim, CA, USA, 24–27 January 1999; Mosquito and Vector Control Association of California: Sacramento, CA, USA, 1999; pp. 47–54. [Google Scholar]
- Keiper, J.B.; Jiannino, J.A.; Sanford, M.R.; Walton, W.E. Effect of vegetation management on the abundance of mosquitoes at a constructed treatment wetland in southern California. In Proceedings of the 70th Annual Conference of the Mosquito and Vector Control Association of California, Fish Camp, CA, USA, 27–30 January 2002; Mosquito and Vector Control Association of California: Sacramento, CA, USA, 2003; pp. 35–43. [Google Scholar]
- Walton, W.E. Effectiveness of pyramidal emergence traps for estimating production of Culex mosquitoes (Diptera: Culicidae). J. N. Am. Benthol. Soc. 2009, 28, 676–682. [Google Scholar] [CrossRef]
- Duguma, D.; Rugman-Jones, P.; Kaufman, M.G.; Hall, M.W.; Neufeld, J.D.; Stouthamer, R.; Walton, W.E. Bacterial communities associated with Culex mosquito larvae and two emergent aquatic plants of bioremediation importance. PLoS ONE 2013, 8, e72522. [Google Scholar] [CrossRef] [PubMed]
- Hinojosa-Garro, D.; Mason, C.F.; Underwood, G.J.C. Influence of macrophyte spatial architecture on periphyton and macroinvertebrate community structure in shallow water bodies under contrasting land management. Fundam. Appl. Limnol. 2010, 177, 19–37. [Google Scholar] [CrossRef]
- Thullen, J.S.; Sartoris, J.J.; Walton, W.E. Effects of vegetation management in constructed wetland treatment cells on water quality and mosquito production. Ecol. Eng. 2002, 18, 441–457. [Google Scholar] [CrossRef]
- Knight, R.L.; Walton, W.E.; O’Meara, G.F.; Reisen, W.K.; Wass, R. Strategies for effective mosquito control in constructed treatment wetlands. Ecol. Eng. 2003, 21, 211–232. [Google Scholar] [CrossRef]
- Rey, J.R.; Walton, W.E.; Wolfe, R.J.; Connelly, C.R.; O’Connell, S.M.; Berg, J.A.; Sakolsky-Hoopes, G.E.; Laderman, A.D. North American wetlands and mosquito control. Int. J. Environ. Res. Public Health 2012, 9, 4537–4605. [Google Scholar] [CrossRef] [PubMed]
- Bowler, P.; Bretz, W.; (University of California-Irvine, Irvine, CA, USA). Personal communication, 2012.
- Mokry, L.; (Alan Plummer Associates Inc., College Station, TX, USA). Personal communication, 2014.
- Rollins, G.L. A Guide to Waterfowl Habitat Management in Suisun Marsh; California Department of Fish and Game: Sacramento, CA, USA, 1981.




{kind=link}
{kind=link}
{kind=link}
| Height and Density | November 2012 | July 2013 | ||
|---|---|---|---|---|
| Mean ± SD | N | Mean ± SD | N | |
| B. maritimus height (cm) | ||||
| Raised | 79.8 ± 13.0 | 142 | 122.1 ± 16.5 | 107 |
| Ambient | 83.7 ± 15.4 | 128 | 118.0 ± 17.3 | 91 |
| B. maritimus density (number m−2) * | ||||
| Raised | 56.3 ± 46.2 | 90 | 16.4 ± 39.1 | 90 |
| Ambient | 37.0 ± 47.0 | 90 | 14.4 ± 40.6 | 90 |
| E. crus-galli height (cm) | _ | _ | ||
| Raised | 192.9 ± 71.1 | 371 | ||
| Ambient | 167.9 ± 45.3 | 128 | ||
| E. crus-galli density (number m−2) | ||||
| Raised | 103.6 ± 99.4 | 90 | ||
| Ambient | 49.3 ± 94.5 | 90 | ||
| Total N | NH4-N | NO2-N | NO3-N | TKN | Total P | COD | |
|---|---|---|---|---|---|---|---|
| Before manipulation: November 2012 | |||||||
| Inflow | 8.56 | 0.27 | 0.17 | 5.52 | 1.21 | 90.0 | |
| Outflow | 6.26 | 0.09 | 0.18 | 4.63 | 0.94 | 101.0 | |
| RE | 27% | 68% | –7% | 16% | 19% | –13% | |
| During manipulation: December 2012–February 2013 | |||||||
| Inflow | 5.61 | 0.28 | 0.15 | 3.05 | 1.20 | 117.7 | |
| Raised | 5.19 | 0.75 | 0.22 | 2.25 | 0.84 | 153.1 | |
| RE | 8% | –166% | –43% | 26% | 30% | –30% | |
| Ambient | 4.47 | 0.29 | 0.13 | 2.22 | 0.84 | 106.6 | |
| RE | 20% | –3% | 13% | 27% | 30% | 9% | |
| After manipulation: April–May 2013 | |||||||
| Inflow | 5.97 | 0.33 | 0.16 | 3.67 | 1.31 | 131.2 | |
| Raised | 3.82 | 0.15 | 0.15 | 2.06 | 1.32 | 118.3 | |
| RE | 36% | 55% | 6% | 44% | –1% | 10% | |
| Ambient | 4.00 | 0.21 | 0.14 | 2.36 | 1.09 | 99.7 | |
| RE | 33% | 35% | 15% | 36% | 16% | 24% | |
| After manipulation: June–August 2013 | |||||||
| Inflow | 6.21 | 0.19 | 4.45 * | 1.76 | 1.48 | 96.9 | |
| Raised | 4.21 | 0.12 | 2.70 * | 1.52 | 1.38 | 81.5 | |
| RE | 32% | 37% | 39% | 14% | 6% | 16% | |
| Ambient | 4.40 | 0.11 | 2.76 * | 1.64 | 1.42 | 78.5 | |
| RE | 29% | 41% | 38% | 7% | 4% | 19% | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walton, W.E.; Duguma, D.; Tao, M.; Popko, D.A.; Nygren, S. Integrated Mosquito Management in Experimental Constructed Wetlands: Efficacy of Small-Stature Macrophytes and Fluctuating Hydroperiod. Water 2016, 8, 421. https://doi.org/10.3390/w8100421
Walton WE, Duguma D, Tao M, Popko DA, Nygren S. Integrated Mosquito Management in Experimental Constructed Wetlands: Efficacy of Small-Stature Macrophytes and Fluctuating Hydroperiod. Water. 2016; 8(10):421. https://doi.org/10.3390/w8100421
Chicago/Turabian StyleWalton, William E., Dagne Duguma, Min Tao, David A. Popko, and Scott Nygren. 2016. "Integrated Mosquito Management in Experimental Constructed Wetlands: Efficacy of Small-Stature Macrophytes and Fluctuating Hydroperiod" Water 8, no. 10: 421. https://doi.org/10.3390/w8100421
APA StyleWalton, W. E., Duguma, D., Tao, M., Popko, D. A., & Nygren, S. (2016). Integrated Mosquito Management in Experimental Constructed Wetlands: Efficacy of Small-Stature Macrophytes and Fluctuating Hydroperiod. Water, 8(10), 421. https://doi.org/10.3390/w8100421