Carbon Dioxide Emissions from the Littoral Zone of a Chinese Reservoir

Abstract
:1. Introduction
2. Materials and Methods
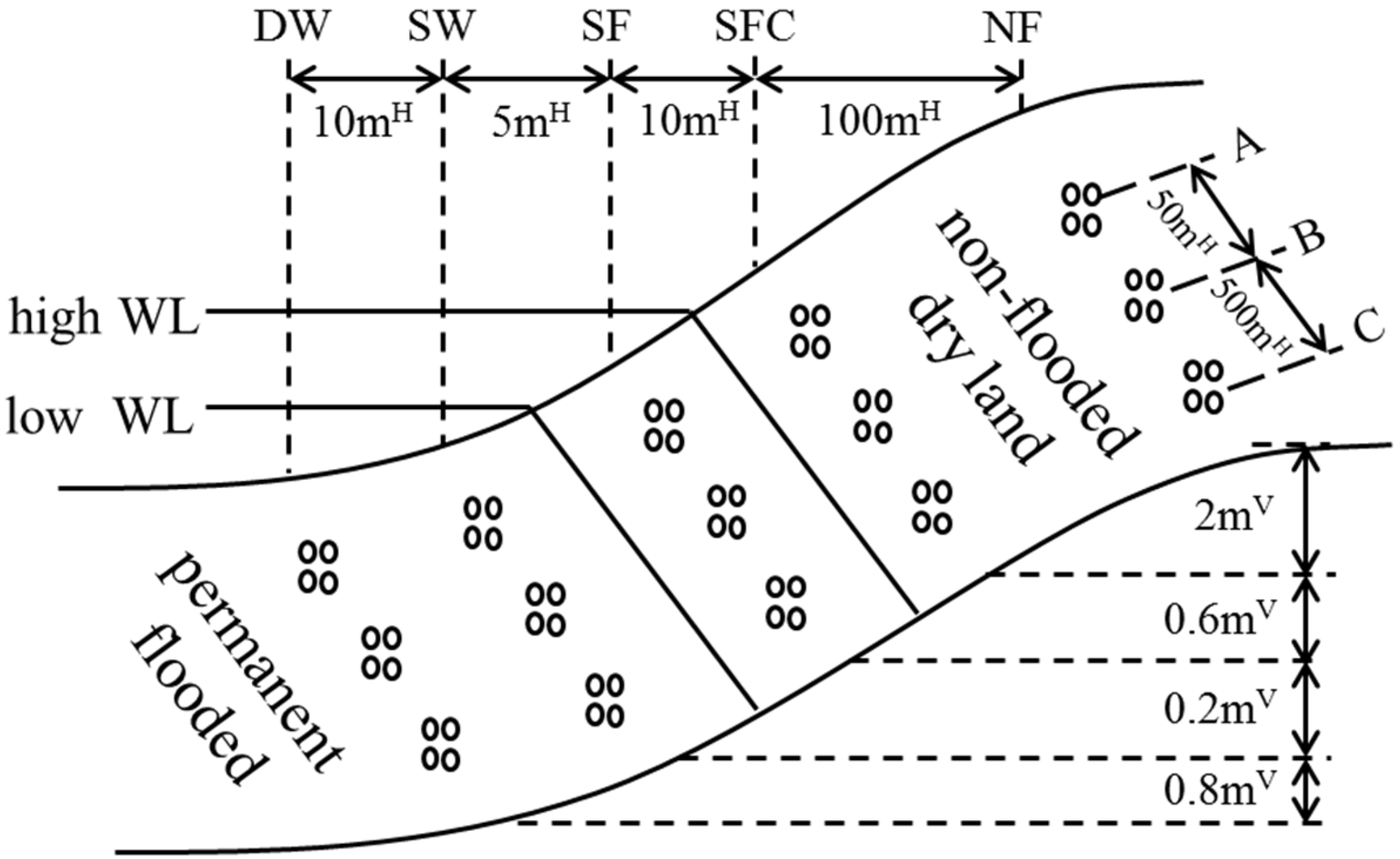
2.1. Study Area and Sampling Sites
2.2. CO2 Flux and Environmental Factors Measurements
2.3. Greenhouse Gases Balance Assessment
2.4. Statistical Analysis
3. Results
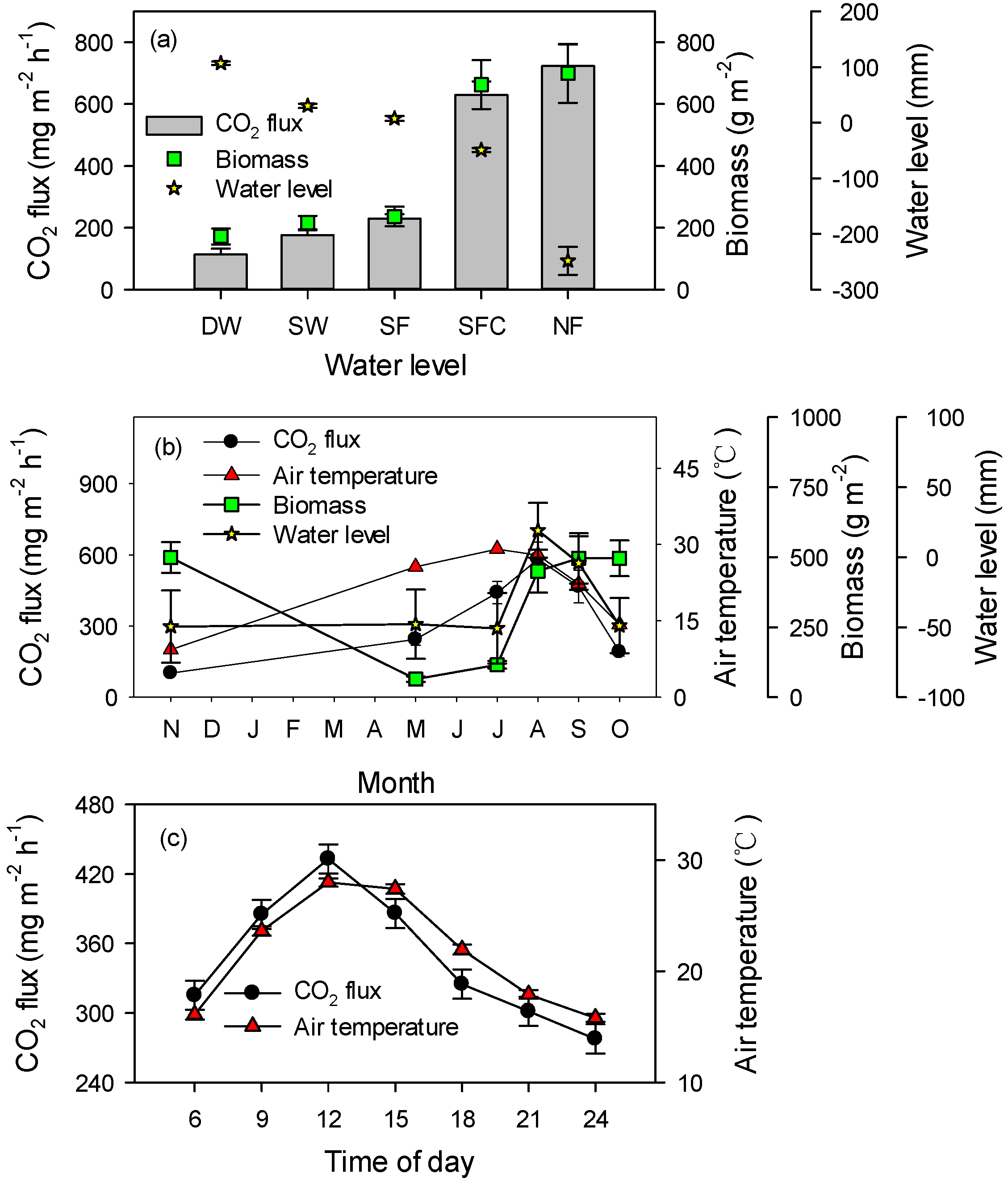
3.1. CO2 Flux and Environmental Influences
3.2. Greenhouse Gas Balance
4. Discussion
4.1. CO2 Flux
4.2. Influences of Environment
4.3. Comparison with Other Greenhouse Gases and the Source/Sink Question
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yang, L.; Lu, F.; Zhou, X.; Wang, X.; Duan, X.; Sun, B. Progress in the studies on the greenhouse gas emissions from reservoirs. Acta Ecol. Sin. 2014, 34, 204–212. [Google Scholar] [CrossRef]
- Zarfl, C.; Lumsdon, A.E.; Berlekamp, J.; Tydecks, L.; Tockner, K. A global boom in hydropower dam construction. Aquat. Sci. 2015, 77, 161–170. [Google Scholar] [CrossRef]
- St. Louis, V.L.; Kelly, C.A.; Duchemin, R.I.C.; Rudd, J.W.M.; Rosenberg, D.M. Reservoir surfaces as sources of greenhouse gases to the atmosphere: A global estimate. Bioscience 2000, 50, 766–775. [Google Scholar]
- Deemer, B.R.; Harrison, J.A.; Li, S.; Beaulieu, J.J.; Delsontro, T.; Barros, N.; Bezerraneto, J.F.; Powers, S.M.; Santos, M.A.D.; Vonk, J.A. Greenhouse gas emissions from reservoir water surfaces: A new global synthesis. Boioscience 2016, 66, 949–964. [Google Scholar] [CrossRef]
- Li, S.; Zhang, Q. Carbon emission from global hydroelectric reservoirs revisited. Environ. Sci.Pollut. Res. 2014, 21, 13636–13641. [Google Scholar] [CrossRef] [PubMed]
- López Bellido, J.; Tulonen, T.; Kankaala, P.; Ojala, A. CO2 and CH4 fluxes during spring and autumn mixing periods in a boreal lake (Pääjärvi, southern Finland). J. Geophys. Res. Biogeosci. 2009, 114. G04007. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Q.; Katul, G.G.; Cole, J.J.; Iii, F.S.C.; Macintyre, S. Large CO2 effluxes at night and during synoptic weather events significantly contribute to CO2 emissions from a reservoir. Environ. Res. Lett. 2016, 11, 064001. [Google Scholar] [CrossRef]
- Roland, F.; Vidal, L.O.; Pacheco, F.S.; Barros, N.O.; Assireu, A.; Ometto, J.; Cimbleris, A.C.P.; Cole, J.J. Variability of carbon dioxide flux from tropical (Cerrado) hydroelectric reservoirs. Aquat. Sci. 2010, 72, 283–293. [Google Scholar] [CrossRef]
- Gebremichael, A.W.; Osborne, B.; Orr, P. Flooding-related increases in CO2 and N2O emissions from a temperate coastal grassland ecosystem. Biogeosciences 2017, 14, 2611–2626. [Google Scholar] [CrossRef]
- Harrison, J.A.; Deemer, B.R.; Birchfield, M.K.; O’Malley, M.T. Reservoir water-level drawdowns accelerate and amplify methane emission. Environ. Sci. Technol. 2017, 51, 1267. [Google Scholar] [CrossRef] [PubMed]
- Oelbermann, M.; Schiff, S.L. Quantifying carbon dioxide and methane emissions and carbon dynamics from flooded boreal forest soil. J. Environ. Qual. 2008, 37, 2037–2047. [Google Scholar] [CrossRef] [PubMed]
- Catalan, N.; von Schiller, D.; Marce, R.; Koschorreck, M.; Gomez-Gener, L.; Obrador, B. Carbon dioxide efflux during the flooding phase of temporary ponds. Limnetica 2014, 33, 349–359. [Google Scholar]
- Dušek, J.; Čížková, H.; Czerný, R.; Taufarová, K.; Šmídová, M.; Janouš, D. Influence of summer flood on the net ecosystem exchange of CO2 in a temperate sedge-grass marsh. Agric. For. Meteorol. 2009, 149, 1524–1530. [Google Scholar] [CrossRef]
- Hu, Q.; Wu, Q.; Yao, B.; Xu, X. Ecosystem respiration and its components from a Carex meadow of Poyang Lake during the drawdown period. Atmos. Environ. 2015, 100, 124–132. [Google Scholar] [CrossRef]
- Jin, H.; Yoon, T.K.; Lee, S.H.; Kang, H.; Im, J.; Park, J.H. Enhanced greenhouse gas emission from exposed sediments along a hydroelectric reservoir during an extreme drought event. Environ. Res. Lett. 2016, 11, 124003. [Google Scholar] [CrossRef]
- Hou, C.; Song, C.; Li, Y.; Wang, J.; Song, Y.; Wang, X. Effects of water table changes on soil CO2, CH4 and N2O fluxes during the growing season in freshwater marsh of northeast China. Environ. Earth Sci. 2012, 69, 1–9. [Google Scholar] [CrossRef]
- Yang, M.; Geng, X.M.; Grace, J.; Jia, Y.F.; Liu, Y.Z.; Jiao, S.W.; Shi, L.L.; Lu, C.; Zhou, Y.; Lei, G.C. N2O fluxes from the littoral zone of a Chinese reservoir. Biogeosciences 2015, 12, 4711–4723. [Google Scholar] [CrossRef]
- Yang, M.; Geng, X.; Grace, J.; Lu, C.; Zhu, Y.; Zhou, Y.; Lei, G. Spatial and seasonal CH4 flux in the littoral zone of Miyun Reservoir near Beijing: The effects of water level and its fluctuation. PLoS ONE 2014, 9, e94275. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.Q. Beijing Chronicles of Water Conservancy, Volume III; Committee of the Beijing Chronicles of Water Conservancy: Beijing, China, 1989. [Google Scholar]
- Cao, R.; Li, C.; Liu, L.; Wang, J.; Yan, G. Extracting Miyun Reservoirs water area and monitoring its change based on a revised normalized different water index. Sci. Surv. Mapp. 2008, 33, 158–160. [Google Scholar]
- Burnham, K.P.; Anderson, D.R.; Huyvaert, K.P. AIC model selection and multimodel inference in behavioral ecology: Some background, observations, and comparisons. Behav. Ecol. Sociobiol. 2011, 65, 23–35. [Google Scholar] [CrossRef]
- Zou, J.W.; Huang, Y.; Zheng, Z.H.; Wang, Y.S.; Chen, Y.Q. Static opaque chamber-based technique for determination of net exchange of CO2 between terrestrial ecosystem and atmosphere. Chin. Sci. Bull. 2004, 49, 381–388. [Google Scholar] [CrossRef]
- Kuzyakov, Y. Separating microbial respiration of exudates from root respiration in non-sterile soils: A comparison of four methods. Soil. Biol. Biochem. 2002, 34, 1621–1631. [Google Scholar] [CrossRef]
- Stocker, T.F.; Qin, D.; Plattner, G.-K.; Tignor, M.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Intergovernmental Panel on Climate Change (IPCC). 2006 Ipcc Guidelines for National Greenhouse Gas Inventories. Volume 4 Agriculture, Forestry and other Land Use; Intergovernmental Panel on Climate Change: Bracknell, UK, 2006; Available online: http://www.Ipcc-nggip.Iges.Or.Jp/public/2006gl/index.htm (accessed on 17 September 2007).
- Tranvik, L.J.; Downing, J.A.; Cotner, J.B.; Loiselle, S.A.; Striegl, R.G.; Ballatore, T.J.; Dillon, P.; Finlay, K.; Fortino, K.; Knoll, L.B.; et al. Lakes and reservoirs as regulators of carbon cycling and climate. Limnol. Oceanogr. 2009, 54, 2298–2314. [Google Scholar] [CrossRef]
- Pace, M.L.; Prairie, Y.T. Respiration in lakes. In Respiration in Aquatic Ecosystems; Oxford University Press: New York, NY, USA, 2005; pp. 103–121. [Google Scholar]
- Soumis, N.; Duchemin, E.; Canuel, R.; Lucotte, M. Greenhouse gas emissions from reservoirs of the western United States Glob. Biogeochem. Cycles 2004, 18. GB3022. [Google Scholar] [CrossRef]
- Morales-Pineda, M.; Cózar, A.; Laiz, I.; Úbeda, B.; Gálvez, J.Á. Daily, biweekly, and seasonal temporal scales of pCO2 variability in two stratified Mediterranean reservoirs. J. Geophys. Res. Biogeosci. 2014, 119. 2013JG002317. [Google Scholar] [CrossRef]
- Jacinthe, P.A.; Filippelli, G.M.; Tedesco, L.P.; Raftis, R. Carbon storage and greenhouse gases emission from a fluvial reservoir in an agricultural landscape. Catena 2012, 94, 53–63. [Google Scholar] [CrossRef]
- Demarty, M.; Bastien, J.; Tremblay, A.; Hesslein, R.H.; Gill, R. Greenhouse gas emissions from boreal reservoirs in Manitoba and Québec, Canada, measured with automated systems. Environ. Sci. Technol. 2009, 43, 8908–8915. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, Z.; Xiao, Y.; Guo, J.; Wu, S.; Liu, J. Spatio-temporal variations of carbon dioxide and its gross emission regulated by artificial operation in a typical hydropower reservoir in China. Environ. Monit. Assess. 2014, 186, 3023–3039. [Google Scholar] [CrossRef] [PubMed]
- Metcalfe, D.B.; Fisher, R.A.; Wardle, D.A. Plant communities as drivers of soil respiration: Pathways,mechanisms, and significance for global change. Biogeosciences 2011, 8, 2047–2061. [Google Scholar] [CrossRef]
- Lloyd, J.; Taylor, J. On the temperature dependence of soil respiration. Funct. Ecol. 1994, 315–323. [Google Scholar] [CrossRef]
- Xu, L.K.; Baldocchi, D.D.; Tang, J.W. How soil moisture, rain pulses, and growth alter the response of ecosystem respiration to temperature. Glob. Biogeochem. Cycles 2004, 18. GB4002. [Google Scholar] [CrossRef]
- Banach, K.; Banach, A.M.; Lamers, L.P.; De Kroon, H.; Bennicelli, R.P.; Smits, A.J.; Visser, E.J. Differences in flooding tolerance between species from two wetland habitats with contrasting hydrology: Implications for vegetation development in future floodwater retention areas. Ann. Bot. 2009, 103, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Striker, G.G. Chapter 1—Flooding stress on plants: Anatomical, morphological and physiological responses. In Botany; Mworia, J., Ed.; InTech: Rijeka, Croatia, 2012; pp. 1–28. [Google Scholar]
- Luo, Y.; Zhou, X. Chapter 5—Controlling factors. In Soil Respiration and the Environment; Luo, Y., Zhou, X., Eds.; Academic Press: Burlington, VT, USA, 2006; pp. 79–105. [Google Scholar]
- McNicol, G.; Silver, W.L. Separate effects of flooding and anaerobiosis on soil greenhouse gas emissions and redox sensitive biogeochemistry. J. Geophys. Res.Biogeosci. 2014, 119, 557–566. [Google Scholar] [CrossRef]
- Bolpagni, R.; Folegot, S.; Laini, A.; Bartoli, M. Role of ephemeral vegetation of emerging river bottoms in modulating CO2 exchanges across a temperate large lowland river stretch. Aquat. Sci. 2017, 79, 149–158. [Google Scholar] [CrossRef]
- Austin, A.T.; Ballaré, C.L. Dual role of lignin in plant litter decomposition in terrestrial ecosystems. Proc. Nat. Acad. Sci. USA 2010, 107, 4618–4622. [Google Scholar] [CrossRef] [PubMed]
- Zenone, T.; Zona, D.; Gelfand, I.; Gielen, B.; Caminoserrano, M.; Ceulemans, R. CO2 uptake is offset by CH4 and N2O emissions in a poplar short-rotation coppice. Glob. Chang. Biol. Bioenergy 2016, 8, 524–538. [Google Scholar] [CrossRef]
- Merbold, L.; Eugster, W.; Stieger, J.; Zahniser, M.; Nelson, D.; Buchmann, N. Greenhouse gas budget (CO2, CH4 and N2O) of intensively managed grassland following restoration. Glob. Chang. Bioenergy 2014, 20, 1913. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Xu, L.; Huang, W.; Ju, X.; Cui, S. Carbon benefits completely offset by nitrogen fertilization induced greenhouse gas emissions in Chinese main cropping systems. In Proceedings of the 2016 International Nitrogen Initiative Conference “Solutions to Improve Nitrogen Use Efficiency for the World”; Melbourne, Australia, 4–8 December 2016. [Google Scholar]
- Tian, H.; Chen, G.; Lu, C.; Xu, X.; Hayes, D.J.; Wei, R.; Pan, S.; Huntzinger, D.N.; Wofsy, S.C. North American terrestrial CO2 uptake largely offset by CH4 and N2O emissions: Toward a full accounting of the greenhouse gas budget. Clim. Chang. 2015, 129, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Audet, J.; Elsgaard, L.; Kjaergaard, C.; Larsen, S.E.; Hoffmann, C.C. Greenhouse gas emissions from a Danish riparian wetland before and after restoration. Ecol. Eng. 2013, 57, 170–182. [Google Scholar] [CrossRef]
- Lee, S.C.; Christen, A.; Black, A.T.; Johnson, M.S.; Jassal, R.S.; Ketler, R.; Nesic, Z.; Merkens, M. Annual greenhouse gas budget for a bog ecosystem undergoing restoration by rewetting. Biogeosciences 2017, 14, 2799–2814. [Google Scholar] [CrossRef]
- Järveoja, J.; Peichl, M.; Maddison, M.; Soosaar, K.; Vellak, K.; Karofeld, E.; Teemusk, A.; Mander, Ü. Impact of water table level on annual carbon and greenhouse gas balances of a restored peat extraction area. Biogeosciences 2015, 13, 2637–2651. [Google Scholar] [CrossRef]
- Magnussen, S.; Reed, D. Knowledge Reference for National Forest Assessments–Modeling for Estimation and Monitoring; Food and Agriculture Organization of the United Nations: Rome, Italy, 2004. [Google Scholar]
- Maqbool, C.; Khan, A.B. Biomass and carbon content of emergent macrophytes in Lake Manasbal, Kashmir: Implications for carbon capture and sequestration. Int. J. Sci. Res. Pub. 2013, 3, 1–7. [Google Scholar]
- Tang, J.W.; Misson, L.; Gershenson, A.; Cheng, W.X.; Goldstein, A.H. Continuous measurements of soil respiration with and without roots in a ponderosa pine plantation in the Sierra Nevada Mountains. Agric. For. Meteorol. 2005, 132, 212–227. [Google Scholar] [CrossRef]
- Hanson, P.J.; Edwards, N.T.; Garten, C.T.; Andrews, J.A. Separating root and soil microbial contributions to soil respiration: A review of methods and observations. Biogeochemistry 2000, 48, 115–146. [Google Scholar] [CrossRef]
- Li, H.; Yang, M.; Lei, T.; Zhang, M.; Bridgewater, P.; Lu, C.; Geng, X.; Lei, G. Nitrous oxide emission from the littoral zones of the Miyun Reservoir near Beijing, China. Hydrol. Res. 2015, 46, 811–823. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Bulk Density (g cm−3) | pH | Total Carbon (g kg−1) | Total Nitrogen (g kg−1) | NH4+ (mg kg−1) | NO3− (mg kg−1) |
|---|---|---|---|---|---|---|
| DW-A | 1.03 ± 0.03 | 7.95 ± 0.01 | 19.81 ± 0.77 | 1.57 ± 0.06 | 22.98 ± 1.16 | 8.1 ± 1.24 |
| DW-B | 1.03 ± 0.03 | 7.95 ± 0.01 | 19.81 ± 0.77 | 1.57 ± 0.06 | 23 ± 0 | 8.1 ± 0 |
| DW-C | 1.03 ± 0.03 | 7.84 ± 0.08 | 22.63 ± 0.48 | 1.8 ± 0.03 | 22.93 ± 0.64 | 5.95 ± 0.3 |
| SW-A | 1.38 ± 0.03 | 8.03 ± 0.04 | 8.2 ± 0.65 | 0.65 ± 0.04 | 24.36 ± 0.44 | 6.89 ± 0.56 |
| SW-B | 1.38 ± 0.02 | 8.13 ± 0.1 | 9.19 ± 0.29 | 0.66 ± 0.02 | 25.72 ± 0.62 | 7.55 ± 0.37 |
| SW-C | 1.4 ± 0.05 | 8.09 ± 0.06 | 9.81 ± 1.45 | 0.81 ± 0.13 | 32.79 ± 6.22 | 5.94 ± 0.74 |
| SF-A | 1.39 ± 0.04 | 8.13 ± 0.06 | 7.23 ± 0.27 | 0.56 ± 0.02 | 24.67 ± 3.73 | 6.13 ± 0.48 |
| SF-B | 1.43 ± 0.02 | 7.98 ± 0.03 | 8.22 ± 0.64 | 0.67 ± 0.04 | 28.44 ± 2.42 | 6.59 ± 1.01 |
| SF-C | 1.43 ± 0.05 | 8.03 ± 0.02 | 7.13 ± 0.55 | 0.64 ± 0.03 | 30.01 ± 3.75 | 6.6 ± 0.92 |
| SFC-A | 1.5 ± 0.04 | 8.61 ± 0.02 | 8.2 ± 0.23 | 0.72 ± 0.01 | 4.13 ± 0.23 | 7.86 ± 0.41 |
| SFC-B | 1.49 ± 0.02 | 8.48 ± 0.06 | 8.13 ± 0.44 | 0.71 ± 0.04 | 4.23 ± 0.39 | 7.61 ± 0.19 |
| SFC-C | 1.4 ± 0.04 | 7.76 ± 0.03 | 3.45 ± 0.15 | 0.36 ± 0.01 | 3.08 ± 0.09 | 7.54 ± 0.38 |
| NF-A | 1.66 ± 0.08 | 8.27 ± 0.08 | 2.19 ± 0.62 | 0.14 ± 0.06 | 19.22 ± 0.76 | 5.76 ± 1.71 |
| NF-B | 1.62 ± 0.02 | 7.59 ± 0.06 | 5.5 ± 0.3 | 0.47 ± 0.02 | 24.16 ± 1.17 | 4.39 ± 0.32 |
| NF-C | 1.45 ± 0.01 | 8.15 ± 0.05 | 6.22 ± 0.18 | 0.57 ± 0.02 | 27.07 ± 1.29 | 12.81 ± 0.66 |
| Effect | Degrees of Freedom | F-Value | p-Value |
|---|---|---|---|
| Water level | 4 | 644.6 | <0.001 |
| Month | 5 | 226.3 | <0.001 |
| Time of day | 6 | 19.2 | <0.001 |
| Water level × Month | 17 | 97.4 | <0.001 |
| Water level × Time of day | 24 | 4.1 | <0.001 |
| Month × Time of day | 30 | 1.8 | 0.004 |
| Water level × Month × Time of day | 102 | 1.1 | 0.225 |
| Error | 2068 |
| Gases | Model | SS | K | AIC Value | |
|---|---|---|---|---|---|
| CO2 (SAR) | 1 | F = −1352.65 + 0.11B + 9.05T − 0.52WD + 174.03PH − 3.24NH − 32.97TC + 364.52TN | 5.7 × 107 | 8 | 1390.8 |
| 2 | F = −1459.12 + 0.12B + 9.21T − 0.55WD + 186.69PH − 3.25NH − 3.76TC | 5.8 × 107 | 7 | 1391.2 | |
| 3 | F = −1872.2 + 0.12B + 9.49T − 0.64WD + 231.36PH − 3.04NH | 5.9 × 107 | 6 | 1391.4 | |
| CH4 | 1 | F = 19.580 + 3.255TN + 0.091T + 0.009WD − 4.121PH + 7.833BD | 1942 | 6 | 264.0 |
| 2 | F = 18.641 + 0.256TC + 0.091T + 0.008WD − 3.960PH + 7.611BD | 1945 | 6 | 264.2 | |
| 3 | F = 17.992 + 3.068TN + 0.091T + 0.008WD − 3.663PH + 6.956BD − 0.105NO | 1926 | 7 | 264.8 | |
| N2O | 1 | F = 0.001 × (−37.096 + 3.458NO + 0.551T − 0.019WD + 0.302NH) | 1.5 × 105 | 5 | 875.774 |
| 2 | F = 0.001 × (−29.243 + 3.350NO + 0.547T − 0.018WD) | 1.5 × 105 | 4 | 875.776 | |
| 3 | F = 0.001 × (−26.057 + 3.129NO + 0.495T) | 1.6 × 105 | 3 | 876.087 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, M.; Grace, J.; Geng, X.; Guan, L.; Zhang, Y.; Lei, J.; Lu, C.; Lei, G. Carbon Dioxide Emissions from the Littoral Zone of a Chinese Reservoir. Water 2017, 9, 539. https://doi.org/10.3390/w9070539
Yang M, Grace J, Geng X, Guan L, Zhang Y, Lei J, Lu C, Lei G. Carbon Dioxide Emissions from the Littoral Zone of a Chinese Reservoir. Water. 2017; 9(7):539. https://doi.org/10.3390/w9070539
Chicago/Turabian StyleYang, Meng, John Grace, Xuemeng Geng, Lei Guan, Yamian Zhang, Jialin Lei, Cai Lu, and Guangchun Lei. 2017. "Carbon Dioxide Emissions from the Littoral Zone of a Chinese Reservoir" Water 9, no. 7: 539. https://doi.org/10.3390/w9070539
APA StyleYang, M., Grace, J., Geng, X., Guan, L., Zhang, Y., Lei, J., Lu, C., & Lei, G. (2017). Carbon Dioxide Emissions from the Littoral Zone of a Chinese Reservoir. Water, 9(7), 539. https://doi.org/10.3390/w9070539