
Cryopreservation of Natural Killer Cells Pre-Complexed with Innate Cell Engagers

Abstract
:1. Introduction
2. Material and Methods
2.1. Generation of Recombinant Antibodies, Soluble Antigens and Cell-Surface Antigens
2.2. Biophysical Characterization
2.3. Surface Plasmon Resonance
2.4. ELISA
2.5. Cell Lines and Cell Culture
2.6. Isolation of Peripheral Blood Mononuclear Cells and Enrichment of Human NK Cells
2.7. Flow Cytometric Analysis of ICE® Construct and Antibody Binding to Cells
2.8. Quantification of EpCAM Cell-Surface Expression
2.9. Flow Cytometric Analysis of ICE® Construct and Antibody Retention on the Surface of NK Cells
2.10. Cytotoxicity and NK-Cell Fratricide Assays
2.11. Pre-Complexing and Freeze/Thaw of NK Cell and ICE® Complexes
3. Results
3.1. Generation of Bispecific ICE® Constructs and Their Biophysical Characterization
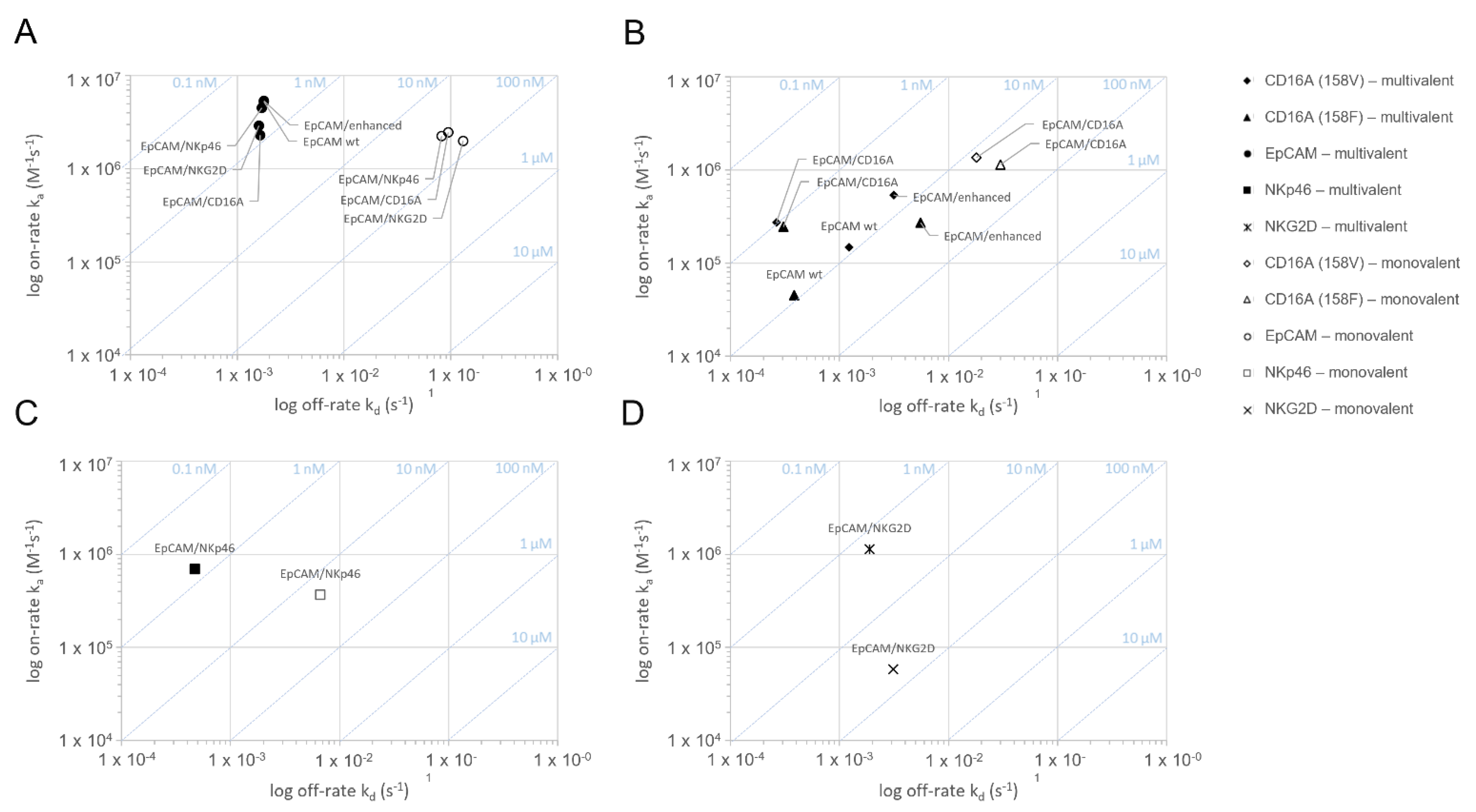
3.2. Surface Plasmon Resonance-Measured Kinetics of the Binding of ICE® Constructs to EpCAM and NK Cell Receptors
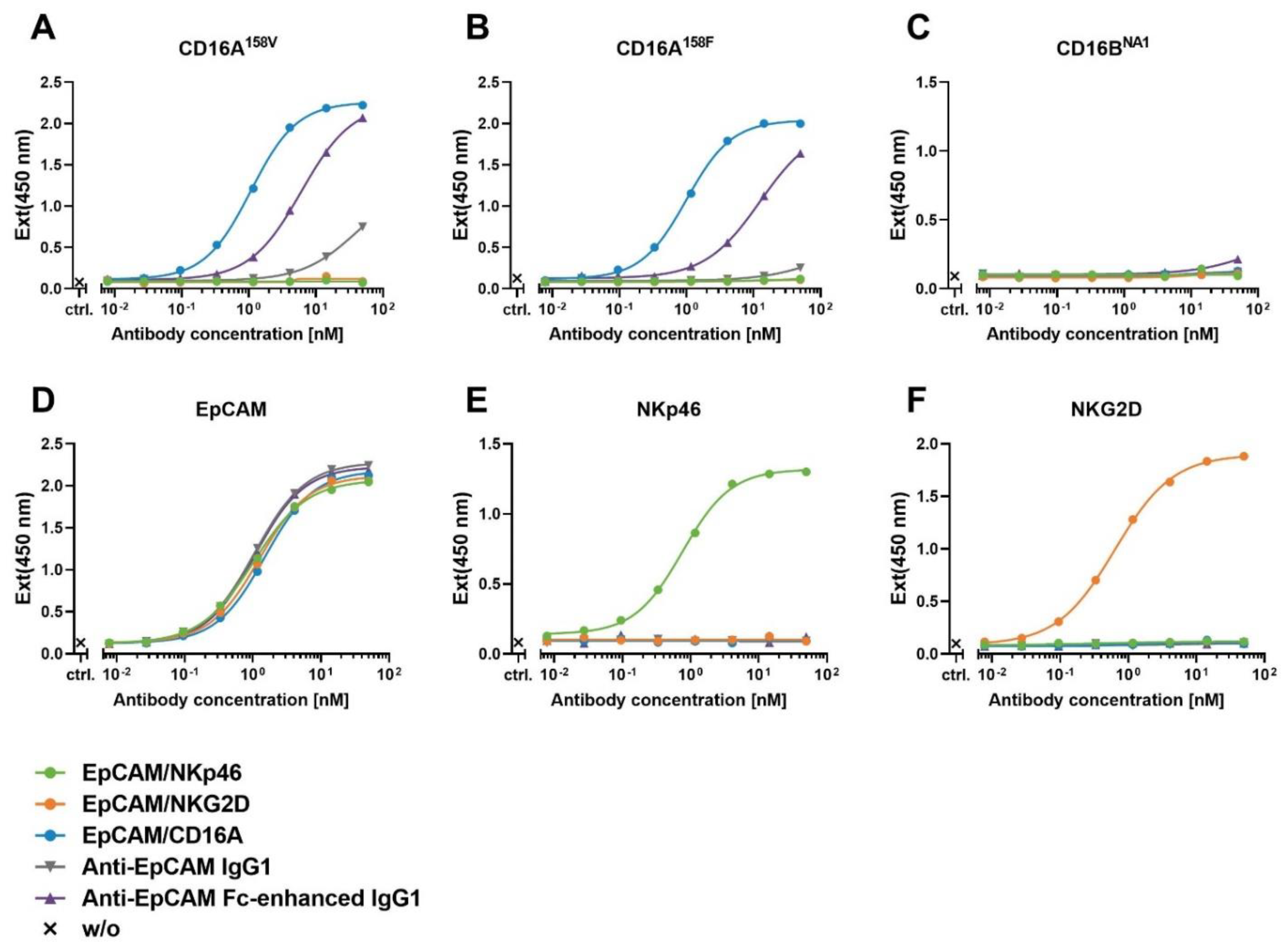
3.3. Assessment of the Binding and Specificity of ICE® Constructs to NK Cell Receptors Measured by Enzyme-Linked Immunosorbent Assay
3.4. Binding of ICE® Constructs to NK Cell Receptors Expressed on the Surface of CHO Cells and to Primary Human NK Cells
3.5. Binding of EpCAM-Specific ICE® Constructs to EpCAM+ Tumor Cell Lines
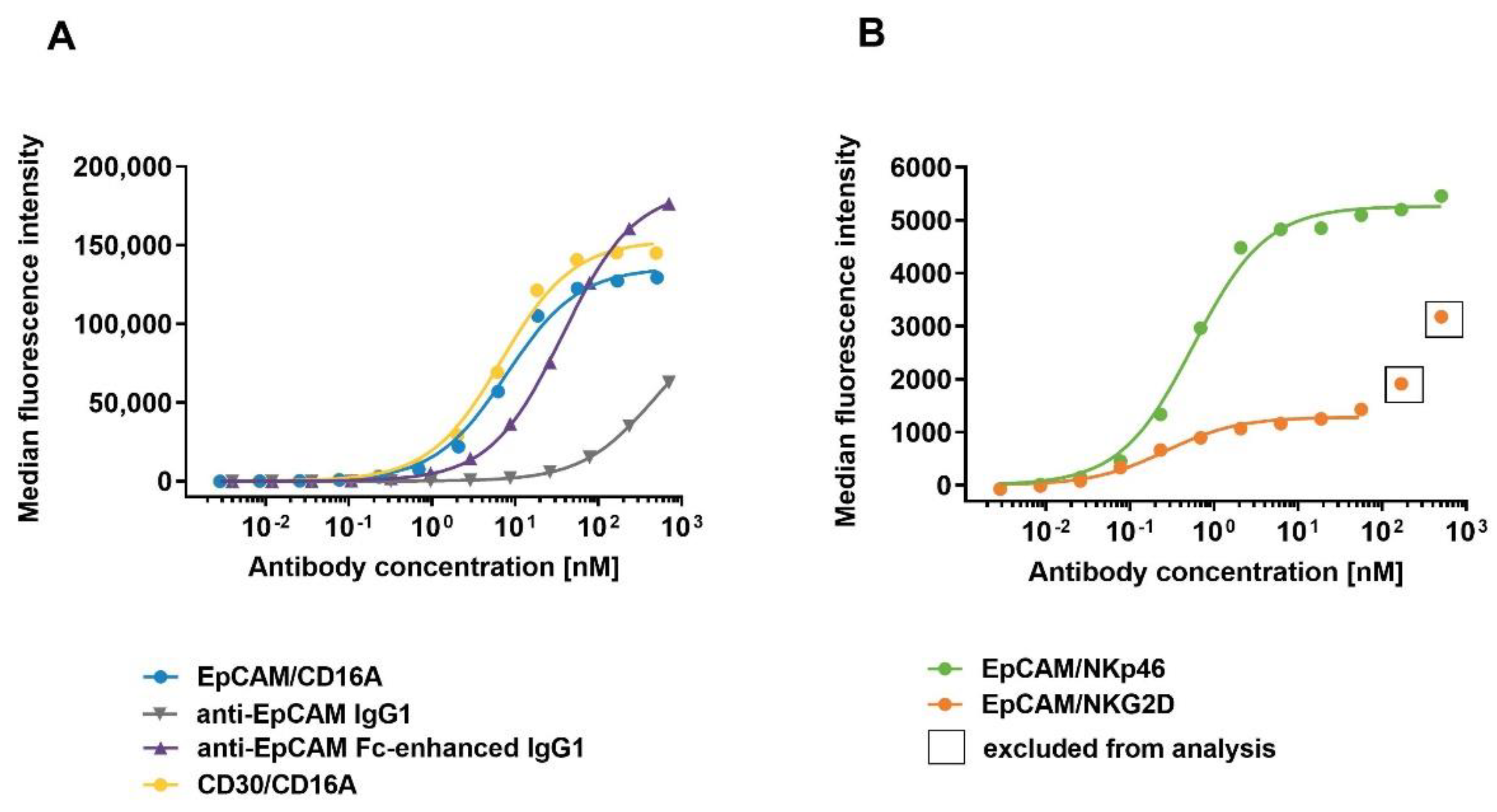
3.6. Assessment of NK Cell Fratricide Induced by the Binding of ICE® Constructs and Anti-EpCAM IgG1 Antibodies to CD16A, NKp46, and NKG2D
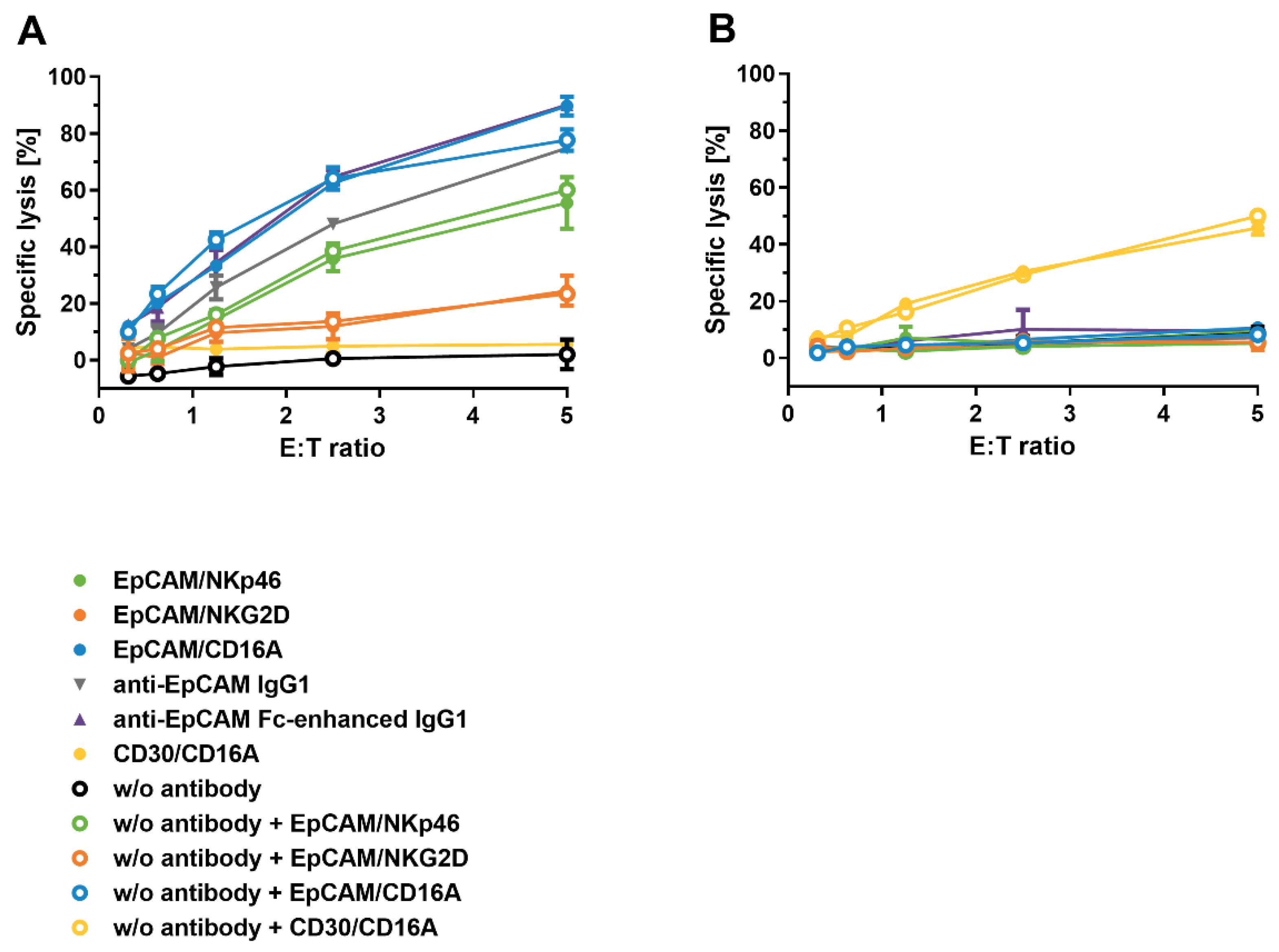
3.7. Target Tumor Cell Lysis Induced by ICE® Constructs
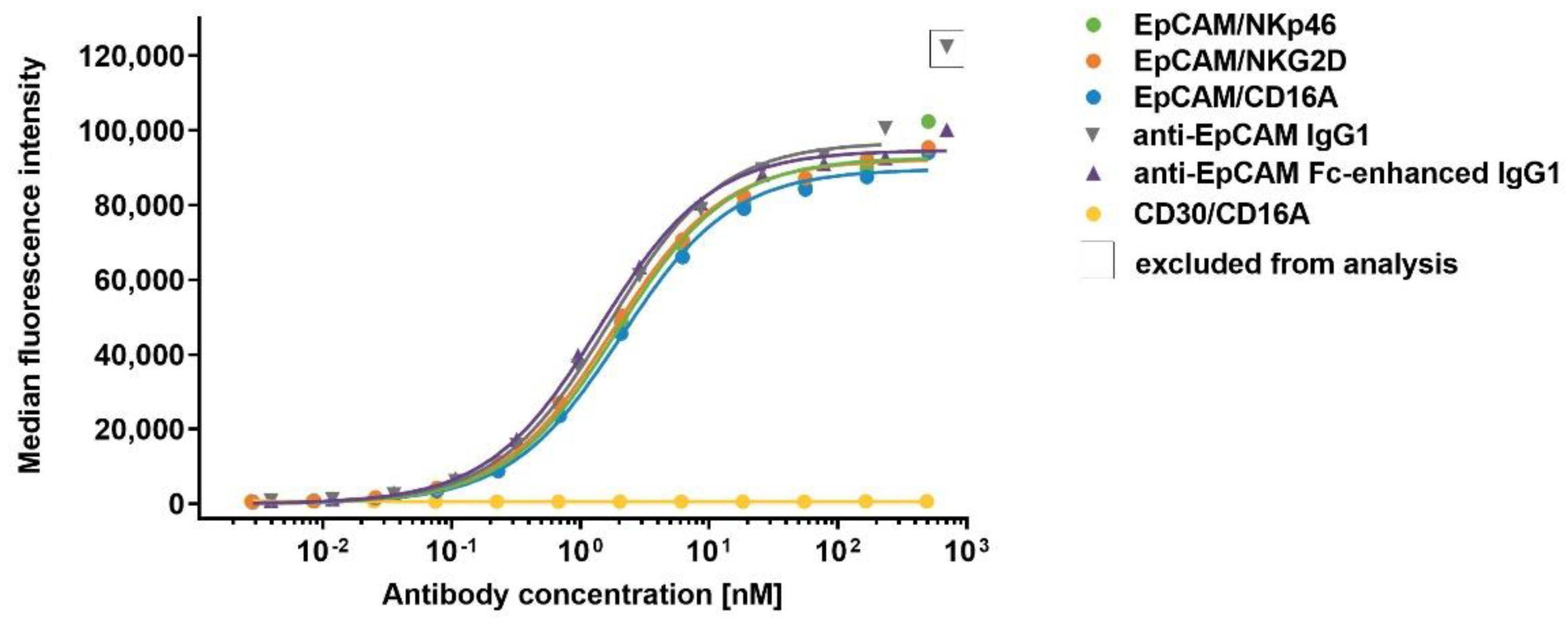
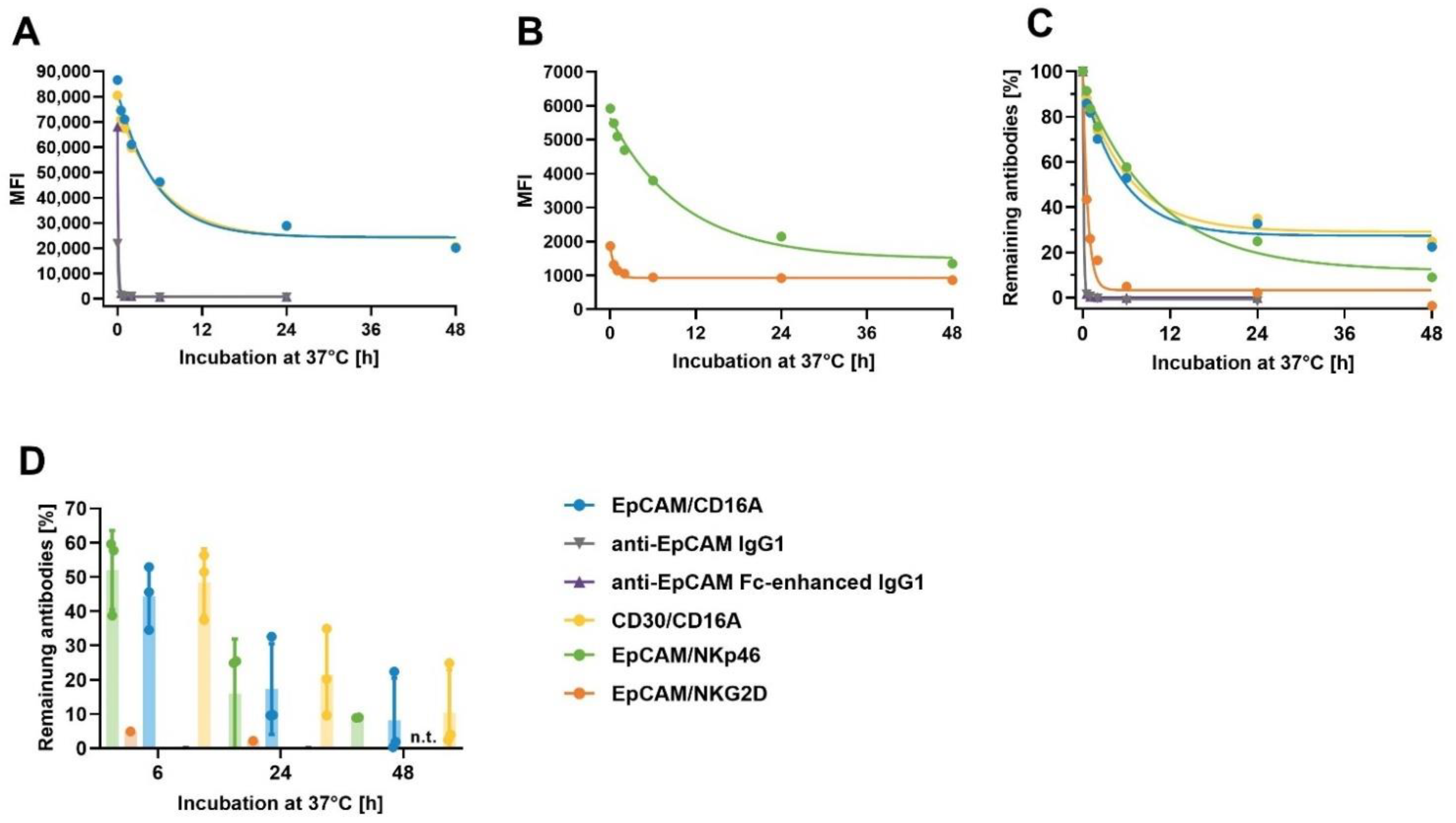
3.8. Assessment of the Cell-Surface Retention of ICE® Constructs on Primary NK Cells
3.9. Cytotoxic Activity of ICE®-Pre-Complexed NK Cells after Reconstitution from Cryopreservation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Demaria, O.; Cornen, S.; Daeron, M.; Morel, Y.; Medzhitov, R.; Vivier, E. Harnessing innate immunity in cancer therapy. Nature 2019, 574, 45–56. [Google Scholar] [CrossRef]
- Gasteiger, G.; D’Osualdo, A.; Schubert, D.A.; Weber, A.; Bruscia, E.M.; Hartl, D. Cellular innate immunity: An old game with new players. J. Innate Immun. 2017, 9, 111–125. [Google Scholar] [CrossRef] [PubMed]
- Vesely, M.D.; Kershaw, M.H.; Schreiber, R.D.; Smyth, M.J. Natural innate and adaptive immunity to cancer. Annu. Rev. Immunol. 2011, 29, 235–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivier, E.; Ugolini, S.; Blaise, D.; Chabannon, C.; Brossay, L. Targeting natural killer cells and natural killer T cells in cancer. Nat. Rev. Immunol. 2012, 12, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Melaiu, O.; Lucarini, V.; Cifaldi, L.; Fruci, D. Influence of the tumor microenvironment on NK cell function in solid tumors. Front. Immunol. 2019, 10, 3038. [Google Scholar] [CrossRef]
- Sivori, S.; Vacca, P.; Del Zotto, G.; Munari, E.; Mingari, M.C.; Moretta, L. Human NK cells: Surface receptors, inhibitory checkpoints, and translational applications. Cell. Mol. Immunol. 2019, 16, 430–441. [Google Scholar] [CrossRef]
- Sivori, S.; Pende, D.; Bottino, C.; Marcenaro, E.; Pessino, A.; Biassoni, R.; Moretta, L.; Moretta, A. NKp46 is the major triggering receptor involved in the natural cytotoxicity of fresh or cultured human NK cells. Correlation between surface density of NKp46 and natural cytotoxicity against autologous, allogeneic or xenogeneic target cells. Eur. J. Immunol. 1999, 29, 1656–1666. [Google Scholar] [CrossRef]
- Wensveen, F.M.; Jelencic, V.; Polic, B. NKG2D: A master regulator of immune cell responsiveness. Front. Immunol. 2018, 9, 441. [Google Scholar] [CrossRef]
- Capuano, C.; Pighi, C.; Battella, S.; De Federicis, D.; Galandrini, R.; Palmieri, G. Harnessing CD16-mediated NK cell functions to enhance therapeutic efficacy of tumor-targeting mAbs. Cancers 2021, 13, 2500. [Google Scholar] [CrossRef]
- Sakamoto, N.; Ishikawa, T.; Kokura, S.; Okayama, T.; Oka, K.; Ideno, M.; Sakai, F.; Kato, A.; Tanabe, M.; Enoki, T.; et al. Phase I clinical trial of autologous NK cell therapy using novel expansion method in patients with advanced digestive cancer. J. Transl. Med. 2015, 13, 277. [Google Scholar] [CrossRef] [Green Version]
- Iliopoulou, E.G.; Kountourakis, P.; Karamouzis, M.V.; Doufexis, D.; Ardavanis, A.; Baxevanis, C.N.; Rigatos, G.; Papamichail, M.; Perez, S.A. A phase I trial of adoptive transfer of allogeneic natural killer cells in patients with advanced non-small cell lung cancer. Cancer Immunol. Immunother. 2010, 59, 1781–1789. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.S.; Soignier, Y.; Panoskaltsis-Mortari, A.; McNearney, S.A.; Yun, G.H.; Fautsch, S.K.; McKenna, D.; Le, C.; Defor, T.E.; Burns, L.J.; et al. Successful adoptive transfer and in vivo expansion of human haploidentical NK cells in patients with cancer. Blood 2005, 105, 3051–3057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubnitz, J.E.; Inaba, H.; Ribeiro, R.C.; Pounds, S.; Rooney, B.; Bell, T.; Pui, C.H.; Leung, W. NKAML: A pilot study to determine the safety and feasibility of haploidentical natural killer cell transplantation in childhood acute myeloid leukemia. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2010, 28, 955–959. [Google Scholar] [CrossRef] [Green Version]
- Clinicaltrials.gov. Available online: https://clinicaltrials.gov/ct2/results?cond=&term=CAR-NK&cntry=&state=&city=&dist= (accessed on 15 December 2021).
- Liu, E.; Marin, D.; Banerjee, P.; Macapinlac, H.A.; Thompson, P.; Basar, R.; Nassif Kerbauy, L.; Overman, B.; Thall, P.; Kaplan, M.; et al. Use of CAR-transduced natural killer cells in CD19-positive lymphoid tumors. N. Engl. J. Med. 2020, 382, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Szmania, S.; Lapteva, N.; Garg, T.; Greenway, A.; Lingo, J.; Nair, B.; Stone, K.; Woods, E.; Khan, J.; Stivers, J.; et al. Ex vivo-expanded natural killer cells demonstrate robust proliferation in vivo in high-risk relapsed multiple myeloma patients. J. Immunother. 2015, 38, 24–36. [Google Scholar] [CrossRef] [Green Version]
- Lapteva, N.; Szmania, S.M.; van Rhee, F.; Rooney, C.M. Clinical grade purification and expansion of natural killer cells. Crit. Rev. Oncog. 2014, 19, 121–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, P.; Raftery, M.J.; Pecher, G. Engineering NK cells for CAR therapy-recent advances in gene transfer methodology. Front. Immunol. 2020, 11, 611163. [Google Scholar] [CrossRef] [PubMed]
- Felices, M.; Lenvik, T.R.; Davis, Z.B.; Miller, J.S.; Vallera, D.A. Generation of BiKEs and TriKEs to improve NK cell-mediated targeting of tumor cells. Methods Mol. Biol. 2016, 1441, 333–346. [Google Scholar] [CrossRef] [Green Version]
- Demaria, O.; Gauthier, L.; Debroas, G.; Vivier, E. Natural killer cell engagers in cancer immunotherapy: Next generation of immuno-oncology treatments. Eur. J. Immunol. 2021, 51, 1934–1942. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, L.; Morel, A.; Anceriz, N.; Rossi, B.; Blanchard-Alvarez, A.; Grondin, G.; Trichard, S.; Cesari, C.; Sapet, M.; Bosco, F.; et al. Multifunctional natural killer cell engagers targeting NKp46 trigger protective tumor immunity. Cell 2019, 177, 1701–1713.e16. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. NCT03214666. Available online: https://clinicaltrials.gov/ct2/show/NCT03214666 (accessed on 25 August 2021).
- ClinicalTrials.gov. NCT04143711. Available online: https://clinicaltrials.gov/ct2/show/NCT04143711 (accessed on 22 November 2021).
- Ellwanger, K.; Reusch, U.; Fucek, I.; Wingert, S.; Ross, T.; Muller, T.; Schniegler-Mattox, U.; Haneke, T.; Rajkovic, E.; Koch, J.; et al. Redirected optimized cell killing (ROCK(R)): A highly versatile multispecific fit-for-purpose antibody platform for engaging innate immunity. mAbs 2019, 11, 899–918. [Google Scholar] [CrossRef] [PubMed]
- Rothe, A.; Sasse, S.; Topp, M.S.; Eichenauer, D.A.; Hummel, H.; Reiners, K.S.; Dietlein, M.; Kuhnert, G. A phase 1 study of the bispecific anti-CD30/CD16A antibody construct AFM13 in patients with relapsed or refractory Hodgkin lymphoma. Blood 2015, 125, 4025–4031. [Google Scholar] [CrossRef] [PubMed]
- Sawas, A.; Chen, P.-H.; Lipschitz, M.; Rodig, S.; Vlad, G. Clinical and biological evaluation of the novel CD30/CD16A tetravalent bispecific antibody (AFM13) in relapsed or refractory CD30-positive lymphoma with cutaneous presentation: A biomarker phase Ib/IIa study (NCT03192202). Blood 2020, 136 (Suppl. S1), 2971. [Google Scholar] [CrossRef]
- Halama, N.; Braun, M.; Kahlert, C.; Spille, A.; Quack, C.; Rahbari, N.; Koch, M.; Weitz, J.; Kloor, M.; Zoernig, I.; et al. Natural killer cells are scarce in colorectal carcinoma tissue despite high levels of chemokines and cytokines. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2011, 17, 678–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavin, Y.; Kobayashi, S.; Leader, A.; Amir, E.D.; Elefant, N.; Bigenwald, C.; Remark, R.; Sweeney, R.; Becker, C.D.; Levine, J.H.; et al. Innate immune landscape in early lung adenocarcinoma by paired single-cell analyses. Cell 2017, 169, 750–765.e717. [Google Scholar] [CrossRef] [Green Version]
- Cai, L.; Zhang, Z.; Zhou, L.; Wang, H.; Fu, J.; Zhang, S.; Shi, M.; Zhang, H.; Yang, Y.; Wu, H.; et al. Functional impairment in circulating and intrahepatic NK cells and relative mechanism in hepatocellular carcinoma patients. Clin. Immunol. 2008, 129, 428–437. [Google Scholar] [CrossRef]
- Kerbauy, L.N.; Marin, N.D.; Kaplan, M.; Banerjee, P.P.; Berrien-Elliott, M.M.; Becker-Hapak, M.; Basar, R.; Foster, M.; Garcia Melo, L.; Neal, C.C.; et al. Combining AFM13, a bispecific CD30/CD16 antibody, with cytokine-activated blood and cord blood-derived NK cells facilitates CAR-like responses against CD30(+) malignancies. Clin. Cancer Res. 2021, 27, 3744–3756. [Google Scholar] [CrossRef]
- Ying, T.; Chen, W.; Gong, R.; Feng, Y.; Dimitrov, D.S. Soluble monomeric IgG1 Fc. J. Biol. Chem. 2012, 287, 19399–19408. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Chang, W.C.; Wong, C.W.; Colcher, D.; Sherman, M.; Ostberg, J.R.; Forman, S.J.; Riddell, S.R.; Jensen, M.C. A transgene-encoded cell surface polypeptide for selection, in vivo tracking, and ablation of engineered cells. Blood 2011, 118, 1255–1263. [Google Scholar] [CrossRef]
- Fuss, I.J.; Kanof, M.E.; Smith, P.D.; Zola, H. Isolation of whole mononuclear cells from peripheral blood and cord blood. Curr. Protoc. Immunol. 2009, 85, 1–7. [Google Scholar] [CrossRef]
- Reusch, U.; Burkhardt, C.; Fucek, I.; Le Gall, F.; Le Gall, M.; Hoffmann, K.; Knackmuss, S.H.; Kiprijanov, S.; Little, M.; Zhukovsky, E.A. A novel tetravalent bispecific TandAb (CD30/CD16A) efficiently recruits NK cells for the lysis of CD30+ tumor cells. mAbs 2014, 6, 728–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eder, M.; Knackmuss, S.; Le Gall, F.; Reusch, U.; Rybin, V.; Little, M.; Haberkorn, U.; Mier, W.; Eisenhut, M. 68Ga-labelled recombinant antibody variants for immuno-PET imaging of solid tumours. Eur. J. Nucl. Med. Mol. Imaging 2010, 37, 1397–1407. [Google Scholar] [CrossRef] [PubMed]
- Kakiuchi-Kiyota, S.; Ross, T.; Wallweber, H.; Kiefer, J.; Schutten, M.; Adedeji, A.; Cai, H.; Hendricks, R.; Cohen, S.; Myneni, S.; et al. A BCMA/CD16A bispecific innate cell engager for the treatment of multiple myeloma. Leukemia 2021, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lazar, G.A.; Dang, W.; Karki, S.; Vafa, O.; Peng, J.S.; Hyun, L.; Chan, C.; Chung, H.S.; Eivazi, A.; Yoder, S.C.; et al. Engineered antibody Fc variants with enhanced effector function. Proc. Natl. Acad. Sci. USA 2006, 103, 4005–4010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casneuf, T.; Xu, X.S.; Adams, H.C., 3rd; Axel, A.E.; Chiu, C.; Khan, I.; Ahmadi, T.; Yan, X.; Lonial, S.; Plesner, T.; et al. Effects of daratumumab on natural killer cells and impact on clinical outcomes in relapsed or refractory multiple myeloma. Blood Adv. 2017, 1, 2105–2114. [Google Scholar] [CrossRef]
- Basar, R.; Daher, M.; Rezvani, K. Next-generation cell therapies: The emerging role of CAR-NK cells. Blood Adv. 2020, 4, 5868–5876. [Google Scholar] [CrossRef]
- Heads, J.T.; Adams, R.; D’Hooghe, L.E.; Page, M.J.; Humphreys, D.P.; Popplewell, A.G.; Lawson, A.D.; Henry, A.J. Relative stabilities of IgG1 and IgG4 Fab domains: Influence of the light-heavy interchain disulfide bond architecture. Protein Sci. 2012, 21, 1315–1322. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Gunasekaran, K.; Wang, W.; Razinkov, V.; Sekirov, L.; Leng, E.; Sweet, H.; Foltz, I.; Howard, M.; Rousseau, A.M.; et al. Asymmetrical Fc engineering greatly enhances antibody-dependent cellular cytotoxicity (ADCC) effector function and stability of the modified antibodies. J. Biol. Chem. 2014, 289, 3571–3590. [Google Scholar] [CrossRef] [Green Version]
- Bryceson, Y.T.; March, M.E.; Ljunggren, H.G.; Long, E.O. Activation, coactivation, and costimulation of resting human natural killer cells. Immunol. Rev. 2006, 214, 73–91. [Google Scholar] [CrossRef] [Green Version]
- Mellor, J.D.; Brown, M.P.; Irving, H.R.; Zalcberg, J.R.; Dobrovic, A. A critical review of the role of Fc gamma receptor polymorphisms in the response to monoclonal antibodies in cancer. J. Hematol. Oncol. 2013, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Musolino, A.; Naldi, N.; Bortesi, B.; Pezzuolo, D.; Capelletti, M.; Missale, G.; Laccabue, D.; Zerbini, A.; Camisa, R.; Bisagni, G.; et al. Immunoglobulin G fragment C receptor polymorphisms and clinical efficacy of trastuzumab-based therapy in patients with HER-2/neu-positive metastatic breast cancer. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2008, 26, 1789–1796. [Google Scholar] [CrossRef] [PubMed]
- Bryceson, Y.T.; Ljunggren, H.G.; Long, E.O. Minimal requirement for induction of natural cytotoxicity and intersection of activation signals by inhibitory receptors. Blood 2009, 114, 2657–2666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pahl, J.H.W.; Koch, J.; Götz, J.J.; Arnold, A.; Reusch, U.; Gantke, T.; Rajkovic, E.; Treder, M.; Cerwenka, A. CD16A Activation of NK Cells Promotes NK Cell Proliferation and Memory-Like Cytotoxicity against Cancer Cells. Cancer Immunol. Res. 2018, 6, 517–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartlett, N.L.; Herrera, A.F.; Domingo-Domenech, E.; Mehta, A.; Forero-Torres, A.; Garcia-Sanz, R.; Armand, P.; Devata, S.; Izquierdo, A.R.; Lossos, I.S.; et al. A phase 1b study of AFM13 in combination with pembrolizumab in patients with relapsed or refractory Hodgkin lymphoma. Blood 2020, 136, 2401–2409. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Constructs | Interaction | KD [nM] | ||||
|---|---|---|---|---|---|---|
| EpCAM | CD16A158V | CD16A158F | NKp46 | NKG2D | ||
| EpCAM/CD16A | monovalent | 38.8 ± 5.6 | 13.9 ± 5.8 | 27.6 ± 10.1 | n.d. | n.d. |
| multivalent | 0.7 ± 0.02 | 1.0 ± 0.05 | 1.2 ± 0.12 | n.d. | n.d. | |
| EpCAM/NKp46 | monovalent | 36.7 ± 5.5 | n.d. | n.d. | 18.0 ± 1.3 | n.d. |
| multivalent | 0.4 ± 0.02 | n.d. | n.d. | 0.7 ± 0.01 | n.d. | |
| EpCAM/NKG2D | monovalent | 68.2 ± 22.6 | n.d. | n.d. | n.d. | 53.6 ± 4.6 |
| multivalent | 0.6 ± 0.03 | n.d. | n.d. | n.d. | 1.7 ± 0.2 | |
| Anti-EpCAM IgG1 | monovalent | n.t. | n.t. | n.t. | n.t. | n.t. |
| multivalent | 0.3 ± 0.004 | 8.2 ± 0.4 | 8.5 ± n.a. | n.d. | n.d. | |
| Anti-EpCAM Fc-enhanced IgG1 | monovalent | n.t. | n.t. | n.t. | n.t. | n.t. |
| multivalent | 0.3 ± 0.04 | 5.9 ± 0.8 | 20.7 ± 3.0 | n.d. | n.d. | |
| Construct | Mean Values of EC50 [nM] ± SD | |||||
|---|---|---|---|---|---|---|
| CD16A158V | CD16A158F | CD16BNA1 | EpCAM | NKp46 | NKG2D | |
| EpCAM/NKp46 | n.d. | n.d. | n.d. | 0.9 ± 0.2 | 0.7 ± 0.1 | n.d. |
| EpCAM/NKG2D | n.d. | n.d. | n.d. | 1.2 ± 0.0 | n.d. | 0.6 ± 0.1 |
| EpCAM/CD16A | 1.1 ± 0.1 | 1.0 ± 0.0 | n.d. | 1.5 ± 0.0 | n.d. | n.d. |
| Anti-EpCAM IgG1 | 42.1 ± 2.4 | 325.9 ± 154.0 | n.d. | 1.1 ± 0.0 | n.d. | n.d. |
| Anti-EpCAM Fc-enhanced IgG1 | 5.2 ± 0.7 | 10.0 ± 2.0 | 251.7 ± 189.8 | 1.0 ± 0.0 | n.d. | n.d. |
| Construct | KD [nM] CD16A158F | KD [nM] CD16A158V | KD [nM] CD16BNA1 | KD [nM] NKp46 | KD [nM] NKG2D |
|---|---|---|---|---|---|
| EpCAM/NKp46 | n.d. | n.d. | n.d. | 1.7 ± 0.6 | n.d. |
| EpCAM/NKG2D | n.d. | n.d. | n.d. | n.d. | 0.6 ± 0.3 |
| EpCAM/CD16A | 27.7 ± 2.0 | 22.7 ± 10.5 | n.d. | n.d. | n.d. |
| Anti-EpCAM IgG1 | 889 ± 396 | 308 ± 152 | 2692 ± 1928 | n.d. | n.d. |
| Anti-EpCAM Fc-enhanced IgG1 | 80.7 ± 40.8 | 26.3 ± 15.8 | 176 ± 71 | n.d. | n.d. |
| CD30/CD16A | 24.9 ± 3.2 | 22.5 ± 9.9 | n.d. | n.d. | n.d. |
| Construct | KD [nM] |
|---|---|
| EpCAM/NKp46 | 0.5 ± 0.2 |
| EpCAM/NKG2D | 0.2 ± 0.1 |
| EpCAM/CD16A | 7.9 ± 0.7 |
| Anti-EpCAM IgG1 | 1292 ± 766 |
| Anti-EpCAM Fc-enhanced IgG1 | 55.5 ± 16.4 |
| CD30/CD16A | 6.7 ± 1.5 |
| Construct | KD [nM] HCC-1954 | KD [nM] Detroit 562 | KD [nM] HCC-1187 |
|---|---|---|---|
| EpCAM/NKp46 | 1.6 ± 0.7 | 1.6 ± 0.3 | 1.5 ± 0.7 |
| EpCAM/NKG2D | 1.7 ± 0.7 | 1.6 ± 0.3 | 1.5 ± 0.7 |
| EpCAM/CD16A | 1.7 ± 0.6 | 2.0 ± 0.3 | 1.9 ± 1.0 |
| Anti-EpCAM IgG1 | 1.6 ± 0.6 | 1.5 ± 0.4 | 1.4 ± 0.7 |
| Anti-EpCAM Fc-enhanced IgG1 | 1.3 ± 0.6 | 1.2 ± 0.2 | 1.2 ± 0.6 |
| CD30/CD16A | n.d. | n.d. | n.d. |
| Construct | EC50 [pM] | Emax [%] |
|---|---|---|
| EpCAM/NKp46 | n.a. | 0.0 ± 0.0 |
| EpCAM/NKG2D | n.a. | 0.0 ± 0.0 |
| EpCAM/CD16A | 2797 ± 1209 | 11.8 ± 3.8 |
| CD30/CD16A | 20893 ± 23747 | 21.7 ± 14.8 |
| Anti-EpCAM IgG1 | n.a. | 13.1 ± 7.5 |
| Anti-EpCAM Fc-enhanced IgG1 | 43242 ± 70322 | 9.0 ± 4.4 |
| Anti-CD38 IgG1 | 73.6 ± 26.7 | 61.1 ± 6.1 |
| Construct | EpCAM−/CD30+ KARPAS-299 | EpCAM+/CD30− HCC-1954 | EpCAM+/CD30− Detroit 562 | EpCAM+/CD30− HCC-1187 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EC50 [pM] | Emax [%] | EC50 [pM] | Emax [%] | EC50 [pM] | Emax [%] | EC50 [pM] | Emax [%] | |||||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| EpCAM/NKp46 | 13994.0 | n.a. | 0.4 | 0.6 | 2.8 | 0.5 | 54.9 | 10.4 | 2.2 | 0.7 | 59.8 | 11.3 | 6.6 | 3.5 | 86.4 | 15.7 |
| EpCAM/NKG2D | n.a. | n.a. | 0.0 | 0.0 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. |
| EpCAM/CD16A | n.a. | n.a. | 0.0 | 0.0 | 2.7 | 0.7 | 64.6 | 5.4 | 1.6 | 0.4 | 70.6 | 12.0 | 2.6 | 1.3 | 101.5 | 9.8 |
| Anti-EpCAM IgG1 | 20717.0 | n.a. | 3.2 | 2.8 | 11.6 | 5.1 | 73.5 | 9.0 | 6.6 | 3.3 | 72.6 | 12.0 | 40.1 | 35.9 | 98.7 | 13.2 |
| Anti-EpCAM Fc-enhanced IgG1 | 5487.0 | n.a. | 1.8 | 3.1 | 3.2 | 0.6 | 69.0 | 8.3 | 1.6 | 0.3 | 69.2 | 12.8 | 6.6 | 4.7 | 100.9 | 6.2 |
| CD30/CD16A | 36.5 | 23.0 | 63.3 | 9.7 | n.a. | n.a. | 0.0 | 0.0 | n.a. | n.a. | 0.0 | 0.0 | n.a. | n.a. | 0.0 | 0.0 |
| ICE® Construct-NK Cell Complexes | Efficacy [%] of Target Cell Lysis at an E:T Ratio of 5:1 | |
|---|---|---|
| HCC-1187 Mean ± SD | KARPAS-299 Mean ± SD | |
| EpCAM/NKp46 | 46.1 ± 11.2 | 5.3 ± 4.0 |
| EpCAM/NKG2D | 14.3 ± 9.1 | 3.7 ± 5.5 |
| EpCAM/CD16A | 74.8 ± 15.3 | 3.1 ± 7.4 |
| Anti-EpCAM IgG1 | 39.2 ± 31.3 | 5.3 ± 2.5 |
| Anti-EpCAM Fc-enhanced IgG1 | 81.8 ± 8.3 | 4.8 ± 4.2 |
| CD30/CD16A | 0.6 ± 5.3 | 35.7 ± 19.3 |
| w/o antibody | 3.5 ± 3.9 | 5.9 ± 3.2 |
| w/o antibody + EpCAM/NKp46 | 57.4 ± 10.0 | 4.2 ± 2.5 |
| w/o antibody + EpCAM/NKG2D | 23.6 ± 1.3 | 3.1 ± 3.1 |
| w/o antibody + EpCAM/CD16A | 83.3 ± 5.0 | 4.8 ± 3.9 |
| w/o antibody + CD30/CD16A | n.t. | 52.1 ± 2.9 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reusch, U.; Ellwanger, K.; Fucek, I.; Müller, T.; Schniegler-Mattox, U.; Koch, J.; Tesar, M. Cryopreservation of Natural Killer Cells Pre-Complexed with Innate Cell Engagers. Antibodies 2022, 11, 12. https://doi.org/10.3390/antib11010012
Reusch U, Ellwanger K, Fucek I, Müller T, Schniegler-Mattox U, Koch J, Tesar M. Cryopreservation of Natural Killer Cells Pre-Complexed with Innate Cell Engagers. Antibodies. 2022; 11(1):12. https://doi.org/10.3390/antib11010012
Chicago/Turabian StyleReusch, Uwe, Kristina Ellwanger, Ivica Fucek, Thomas Müller, Ute Schniegler-Mattox, Joachim Koch, and Michael Tesar. 2022. "Cryopreservation of Natural Killer Cells Pre-Complexed with Innate Cell Engagers" Antibodies 11, no. 1: 12. https://doi.org/10.3390/antib11010012
APA StyleReusch, U., Ellwanger, K., Fucek, I., Müller, T., Schniegler-Mattox, U., Koch, J., & Tesar, M. (2022). Cryopreservation of Natural Killer Cells Pre-Complexed with Innate Cell Engagers. Antibodies, 11(1), 12. https://doi.org/10.3390/antib11010012