
Design and Characterization of Novel Antibody-Cytokine Fusion Proteins Based on Interleukin-21


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Cloning, Expression and Protein Purification
2.2. Protein Characterization
2.3. Bioactivity Measurement
2.4. Cell Lines
2.5. Immunofluorescence Study
2.6. Biodistribution Experiments
2.7. Ethical Statement
3. Results
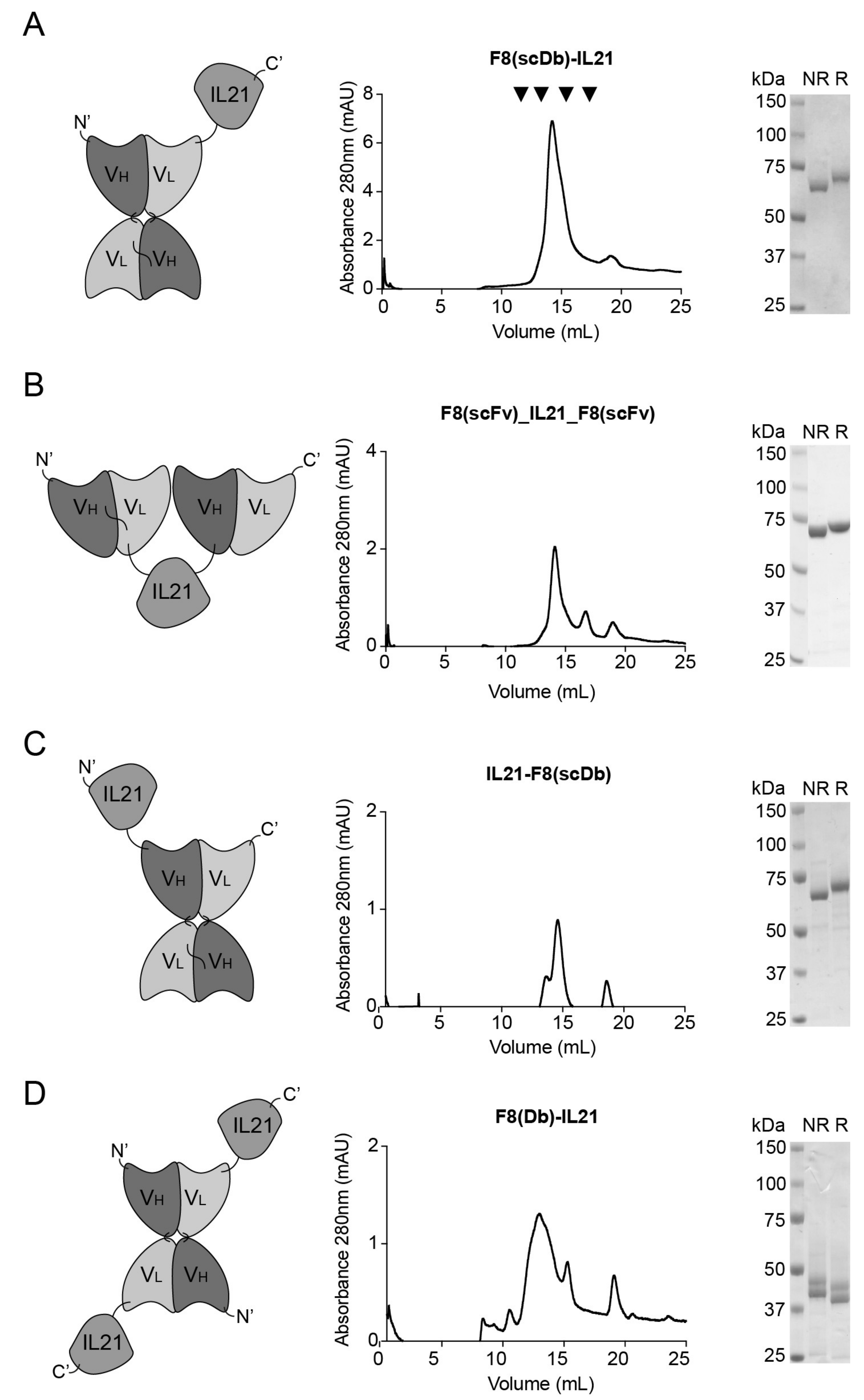
3.1. Generation and Format Screening of Novel IL21-Based Antibody Fusion Proteins
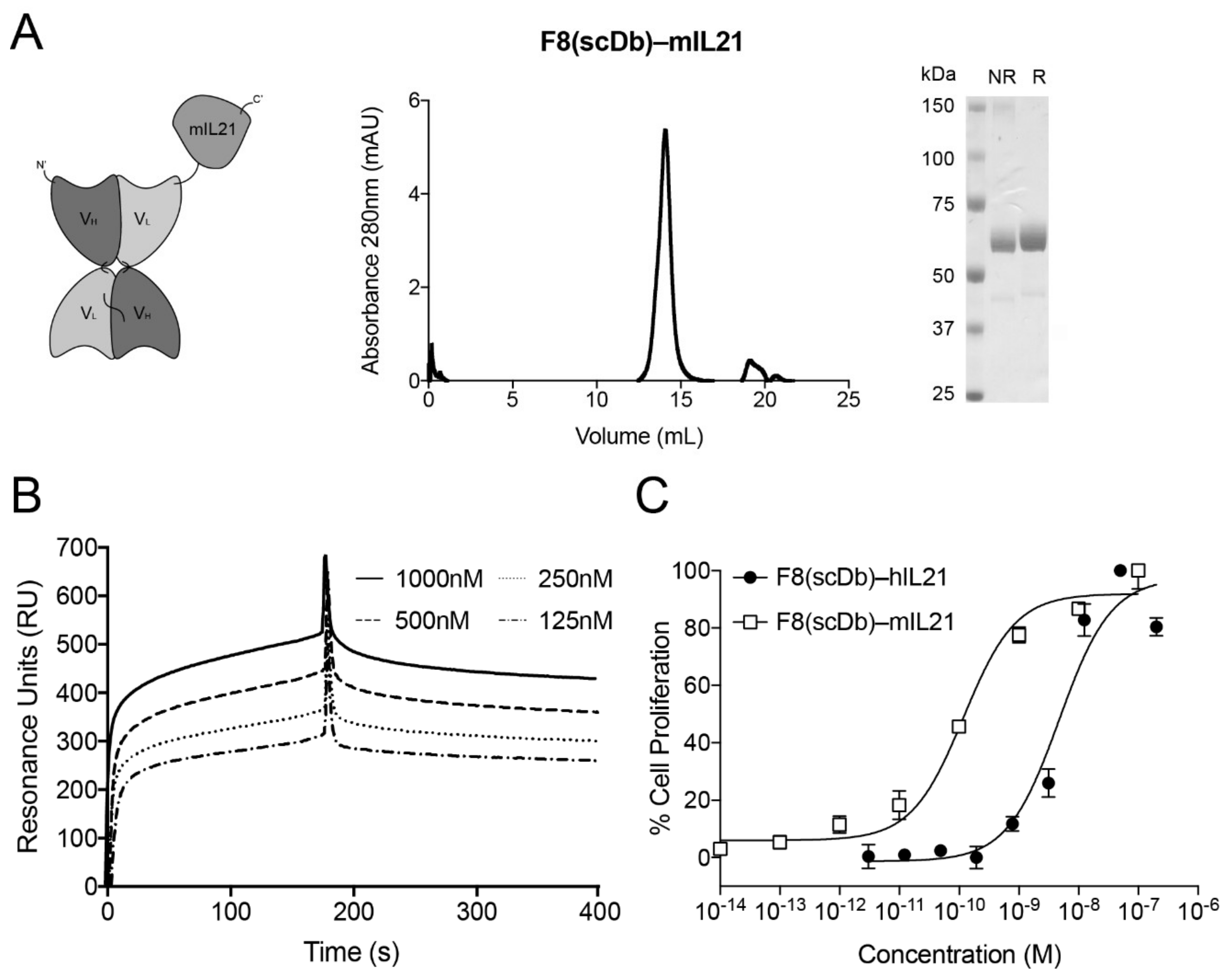
3.2. Biochemical Properties and Activity of F8(scDb)-IL21
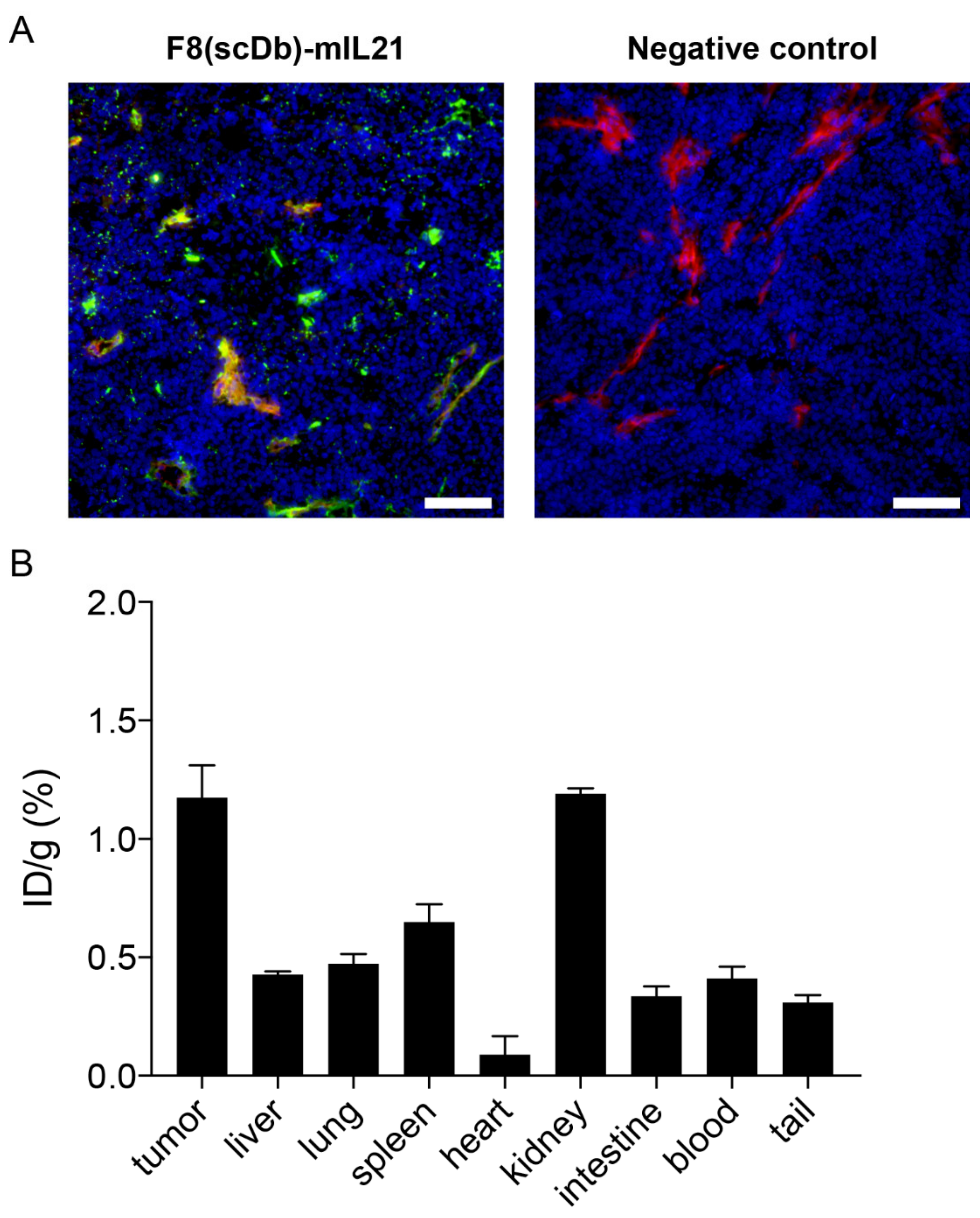
3.3. In Vitro Immunofluorescence Analysis
3.4. In Vivo Tumor Targeting Performance
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neri, D. Antibody–Cytokine Fusions: Versatile Products for the Modulation of Anticancer Immunity. Cancer Immunol. Res. 2019, 7, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Danielli, R.; Patuzzo, R.; Di Giacomo, A.M.; Gallino, G.; Maurichi, A.; Di Florio, A.; Cutaia, O.; Lazzeri, A.; Fazio, C.; Miracco, C.; et al. Intralesional administration of L19-IL2/L19-TNF in stage III or stage IVM1a melanoma patients: Results of a phase II study. Cancer Immunol. Immunother. 2015, 64, 999–1009. [Google Scholar] [CrossRef] [PubMed]
- Weiss, T.; Puca, E.; Silginer, M.; Hemmerle, T.; Pazahr, S.; Bink, A.; Weller, M.; Neri, D.; Roth, P. Immunocytokines are a promising immunotherapeutic approach against glioblastoma. Sci. Transl. Med. 2020, 12, eabb2311. [Google Scholar] [CrossRef] [PubMed]
- Rotte, A.; Bhandaru, M.; Zhou, Y.; McElwee, K.J. Immunotherapy of melanoma: Present options and future promises. Cancer Metastasis Rev. 2015, 34, 115–128. [Google Scholar] [CrossRef]
- Pestka, S.; Krause, C.D.; Walter, M.R. Interferons, interferon-like cytokines, and their receptors. Immunol. Rev. 2004, 202, 8–32. [Google Scholar] [CrossRef]
- Baldo, B.A. Side Effects of Cytokines Approved for Therapy. Drug Saf. 2014, 37, 921–943. [Google Scholar] [CrossRef]
- Neri, D.; Sondel, P.M. Immunocytokines for cancer treatment: Past, present and future. Curr. Opin. Immunol. 2016, 40, 96–102. [Google Scholar] [CrossRef] [Green Version]
- Briukhovetska, D.; Dörr, J.; Endres, S.; Libby, P.; Dinarello, C.A.; Kobold, S. Interleukins in cancer: From biology to therapy. Nat. Cancer 2021, 21, 481–499. [Google Scholar] [CrossRef]
- Runbeck, E.; Crescioli, S.; Karagiannis, S.; Papa, S. Utilizing Immunocytokines for Cancer Therapy. Antibodies 2021, 10, 10. [Google Scholar] [CrossRef]
- Lode, H.N.; Reisfeld, R.A. Targeted Cytokines for Cancer Immunotherapy. Immunol. Res. 2000, 21, 279–288. [Google Scholar] [CrossRef]
- List, T.; Neri, D. Immunocytokines: A review of molecules in clinical development for cancer therapy. Clin. Pharmacol. Adv. Appl. 2013, 5, 29–45. [Google Scholar] [CrossRef] [Green Version]
- Leonard, W.J.; Wan, C.K. IL-21 Signaling in Immunity. F1000Research 2016, 5, 1–10. [Google Scholar] [CrossRef]
- Spolski, R.; Leonard, W.J. Interleukin-21: Basic Biology and Implications for Cancer and Autoimmunity. Annu. Rev. Immunol. 2008, 26, 57–79. [Google Scholar] [CrossRef] [Green Version]
- Moroz, A.; Eppolito, C.; Li, Q.; Tao, J.; Clegg, C.H.; Shrikant, P.A. IL-21 Enhances and Sustains CD8+ T Cell Responses to Achieve Durable Tumor Immunity: Comparative Evaluation of IL-2, IL-15, and IL-21. J. Immunol. 2004, 173, 900–909. [Google Scholar] [CrossRef] [Green Version]
- Shen, S.; Sckisel, G.; Sahoo, A.; Lalani, A.; Otter, D.D.; Pearson, J.; Devoss, J.; Cheng, J.; Casey, S.C.; Case, R.; et al. Engineered IL-21 Cytokine Muteins Fused to Anti-PD-1 Antibodies Can Improve CD8+ T Cell Function and Anti-tumor Immunity. Front. Immunol. 2020, 11, 832. [Google Scholar] [CrossRef]
- Davis, I.D.; Skrumsager, B.K.; Cebon, J.; Nicholaou, T.; Barlow, J.W.; Moller, N.P.; Skak, K.; Lundsgaard, D.; Frederiksen, K.S.; Thygesen, P.; et al. An open-label, two-arm, phase I trial of recombinant human interleukin-21 in patients with metastatic melanoma. Clin. Cancer Res. 2007, 13, 3630–3636. [Google Scholar] [CrossRef] [Green Version]
- Deng, S.; Sun, Z.; Qiao, J.; Liang, Y.; Liu, L.; Dong, C.; Shen, A.; Wang, Y.; Tang, H.; Fu, Y.-X.; et al. Targeting tumors with IL-21 reshapes the tumor microenvironment by proliferating PD-1intTim-3–CD8+ T cells. JCI Insight 2020, 5, e132000. [Google Scholar] [CrossRef] [Green Version]
- Hashmi, M.H.; Van Veldhuizen, P.J. Interleukin-21: Updated review of Phase i and II clinical trials in metastatic renal cell car-cinoma, metastatic melanoma and relapsed/refractory indolent non-Hodgkin’s lymphoma. Expert Opin. Biol. Ther. 2010, 10, 807–817. [Google Scholar] [CrossRef]
- Davis, I.D.; Brady, B.; Kefford, R.F.; Millward, M.; Cebon, J.; Skrumsager, B.K.; Mouritzen, U.; Hansen, L.T.; Skak, K.; Lundsgaard, D.; et al. Clinical and biological efficacy of recombinant human interleukin-21 in patients with stage TV Malignant melanoma without prior treatment: A phase Lla trial. Clin. Cancer Res. 2009, 15, 2123–2129. [Google Scholar] [CrossRef] [Green Version]
- Borsi, L.; Castellani, P.; Allemanni, G.; Neri, D.; Zardi, L. Preparation of phage antibodies to the ED-A domain of human fibron-ectin. Exp. Cell Res. 1998, 240, 244–251. [Google Scholar] [CrossRef]
- Villa, A.; Trachsel, E.; Kaspar, M.; Schliemann, C.; Sommavilla, R.; Rybak, J.N.; Rösli, C.; Borsi, L.; Neri, D. A high-affinity human monoclonal antibody specific to the alternatively spliced EDA domain of fibronectin efficiently targets tumor neo-vasculature in vivo. Int. J. Cancer 2008, 122, 2405–2413. [Google Scholar] [CrossRef]
- Ongaro, T.; Gouyou, B.; Stringhini, M.; Corbellari, R.; Neri, D.; Villa, A. A novel format for recombinant antibody-interleukin-2 fu-sion proteins exhibits superior tumor-targeting properties in vivo. Oncotarget 2020, 11, 3698–3711. [Google Scholar] [CrossRef]
- Tarli, L.; Balza, E.; Viti, F.; Borsi, L.; Castellani, P.; Berndorff, D.; Dinkelborg, L.; Neri, D.; Zardi, L. A High-Affinity Human Antibody That Targets Tumoral Blood Vessels. Blood 1999, 94, 192–198. [Google Scholar] [CrossRef]
- Pasche, N.; Wulhfard, S.; Pretto, F.; Carugati, E.; Neri, D. The antibody-based delivery of interleukin-12 to the tumor neovascula-ture eradicates murine models of cancer in combination with paclitaxel. Clin. Cancer Res. 2012, 18, 4092–4103. [Google Scholar] [CrossRef] [Green Version]
- Corbellari, R.; Stringhini, M.; Mock, J.; Ongaro, T.; Villa, A.; Neri, D.; De Luca, R. A novel Antibody-IL15 fusion protein selectively localizes to tumors, synergizes with TNF-based immunocytokine, and inhibits metastasis. Mol. Cancer Ther. 2021, 20, 859–871. [Google Scholar] [CrossRef]
- Ziffels, B.; Grötsch, A.; Al-Bayati, L.; Neri, D. Targeted delivery of calreticulin to ED-A fibronectin leads to tumor-growth retardation. J. Biotechnol. 2018, 290, 53–58. [Google Scholar] [CrossRef]
- De Luca, R.; Neri, D. Potentiation of PD-L1 blockade with a potency-matched dual cytokine–antibody fusion protein leads to cancer eradication in BALB/c-derived tumors but not in other mouse strains. Cancer Immunol. Immunother. 2018, 67, 1381–1391. [Google Scholar] [CrossRef]
- Scheerlinck, J.-P.Y. Functional and structural comparison of cytokines in different species. Veter.-Immunol. Immunopathol. 1999, 72, 39–44. [Google Scholar] [CrossRef]
- Ibrahim, I.; El-Kolaly, M.; Aboumanei, M.; Abdelbary, A. 125I labeling of clomiphene and biodistribution studies for possible use as a model in breast cancer imaging. Appl. Radiat. Isot. 2016, 115, 37–44. [Google Scholar] [CrossRef]
- Durante, A.C.R.; Sobral, D.V.; Miranda, A.C.C.; de Almeida, É.V.; Fuscaldi, L.L.; de Barboza, M.R.F.F.; Malavolta, L. Comparative study of two oxidizing agents, chloramine T and Iodo-Gen®, for the radiolabeling of β-CIT with iodine-131: Relevance for Parkinson’s disease. Pharmaceuticals 2019, 12, 25. [Google Scholar] [CrossRef] [Green Version]
- Shima, K.; Sawazaki, N.; Tanaka, R.; Tarui, S.; Nishikawa, M. Effect of an Exposure to Chloramine-T on the Immunoreactivity of Glucagon1. Endocrinology 1975, 96, 1254–1260. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Jona, J.; Yamada, A.; Dittert, L. Chloramine-T in Radiolabeling Techniques II. A Nondestructive Method for Radiolabeling Biomolecules by Halogenation. Anal. Biochem. 1995, 224, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Ebbinghaus, C.; Ronca, R.; Kaspar, M.; Grabulovski, D.; Berndt, A.; Kosmehl, H.; Zardi, L.; Neri, D. Engineered vascular-targeting anti-body-interferon-γ fusion protein for cancer therapy. Int. J. Cancer 2005, 116, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Kaspar, M.; Trachsel, E.; Neri, D. The antibody-mediated targeted delivery of interleukin-15 and GM-CSF to the tumor neovas-culature inhibits tumor growth and metastasis. Cancer Res. 2007, 67, 4940–4948. [Google Scholar] [CrossRef] [Green Version]
- Ongaro, T.; Matasci, M.; Cazzamalli, S.; Gouyou, B.; De Luca, R.; Neri, D.; Villa, A. A novel anti-cancer L19-interleukin-12 fusion protein with an optimized peptide linker efficiently localizes in vivo at the site of tumors. J. Biotechnol. 2018, 291, 17–25. [Google Scholar] [CrossRef]
- Holder, P.; Lim, S.; Huang, C.; Sharma, P.; Dagdas, Y.; Bulutoglu, B.; Sockolosky, J. Engineering interferons and interleukins for cancer immunotherapy. Adv. Drug Deliv. Rev. 2022, 182, 114112. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Nitto, C.; Neri, D.; Weiss, T.; Weller, M.; De Luca, R. Design and Characterization of Novel Antibody-Cytokine Fusion Proteins Based on Interleukin-21. Antibodies 2022, 11, 19. https://doi.org/10.3390/antib11010019
Di Nitto C, Neri D, Weiss T, Weller M, De Luca R. Design and Characterization of Novel Antibody-Cytokine Fusion Proteins Based on Interleukin-21. Antibodies. 2022; 11(1):19. https://doi.org/10.3390/antib11010019
Chicago/Turabian StyleDi Nitto, Cesare, Dario Neri, Tobias Weiss, Michael Weller, and Roberto De Luca. 2022. "Design and Characterization of Novel Antibody-Cytokine Fusion Proteins Based on Interleukin-21" Antibodies 11, no. 1: 19. https://doi.org/10.3390/antib11010019
APA StyleDi Nitto, C., Neri, D., Weiss, T., Weller, M., & De Luca, R. (2022). Design and Characterization of Novel Antibody-Cytokine Fusion Proteins Based on Interleukin-21. Antibodies, 11(1), 19. https://doi.org/10.3390/antib11010019