


Establishment and Functional Characterization of Murine Monoclonal Antibodies Recognizing Neuritin
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Cloning, Expression and Purification of Recombinant Protein
2.3. Transient Transfection of HEK293 Cells
2.4. Establishment of Monoclonal Antibodies
2.5. Immunization of Mice
2.6. Production of Fab Fragments
2.7. Enzyme-Linked Immunosorbent Assays
2.8. Flow Cytometry
2.9. Statistical Analysis
3. Results
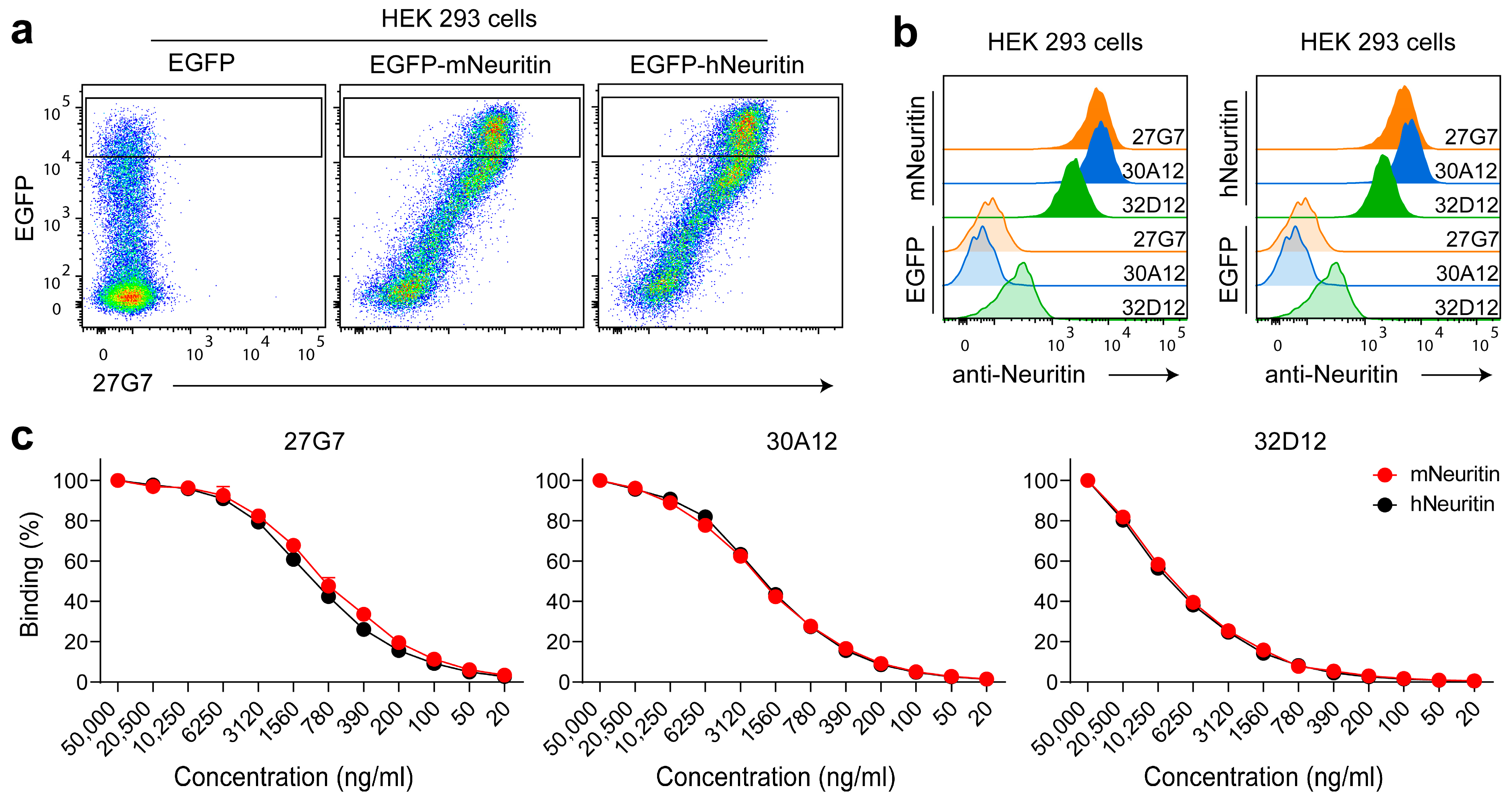
3.1. Establishment and Validation of Mouse Anti-Neuritin Monoclonal Antibodies (mAbs)
3.2. Application of the mAbs in ELISA Assays
3.3. Clones 27G7, 30A12 and 32D12 Recognize Surface-Bound Neuritin Expressed by Murine Follicular Regulatory T
3.4. The mAb Block Binding of Recombinant Neuritin to Germinal Center B Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nedivi, E.; Fieldust, S.; E Theill, L.; Hevron, D. A set of genes expressed in response to light in the adult cerebral cortex and regulated during development. Proc. Natl. Acad. Sci. USA 1996, 93, 2048–2053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naeve, G.S.; Ramakrishnan, M.; Kramer, R.; Hevroni, D.; Citri, Y.; Theill, L.E. Neuritin: A gene induced by neural activity and neurotrophins that promotes neuritogenesis. Proc. Natl. Acad. Sci. USA 1997, 94, 2648–2653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nedivi, E.; Wu, G.-Y.; Cline, H.T. Promotion of Dendritic Growth by CPG15, an Activity-Induced Signaling Molecule. Science 1998, 281, 1863–1866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putz, U.; Harwell, C.; Nedivi, E. Soluble CPG15 expressed during early development rescues cortical progenitors from apoptosis. Nat. Neurosci. 2005, 8, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Picard, N.; Leslie, J.H.; Trowbridge, S.K.; Subramanian, J.; Nedivi, E.; Fagiolini, M. Aberrant Development and Plasticity of Excitatory Visual Cortical Networks in the Absence of cpg15. J. Neurosci. 2014, 34, 3517–3522. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Li, X.; Shan, L.; Zhu, J.; Chen, R.; Li, Y.; Yuan, W.; Yang, L.; Huang, J. Recombinant hNeuritin Promotes Structural and Functional Recovery of Sciatic Nerve Injury in Rats. Front. Neurosci. 2016, 10, 589. [Google Scholar] [CrossRef] [Green Version]
- Azuchi, Y.; Namekata, K.; Shimada, T.; Guo, X.; Kimura, A.; Harada, C.; Saito, A.; Yamagata, K.; Harada, T. Role of neuritin in retinal ganglion cell death in adult mice following optic nerve injury. Sci. Rep. 2018, 8, 10132. [Google Scholar] [CrossRef] [Green Version]
- Fujino, T.; Leslie, J.H.; Eavri, R.; Chen, J.L.; Lin, W.C.; Flanders, G.H.; Borok, E.; Horvath, T.L.; Nedivi, E. CPG15 regulates synapse stability in the developing and adult brain. Genes Dev. 2011, 25, 2674–2685. [Google Scholar] [CrossRef] [Green Version]
- Yao, J.-J.; Zhao, Q.-R.; Lu, J.-M.; Mei, Y.-A. Functions and the related signaling pathways of the neurotrophic factor neuritin. Acta Pharmacol. Sin. 2018, 39, 1414–1420. [Google Scholar] [CrossRef]
- Dong, H.; Luo, X.; Niu, Y.; Yu, N.; Gao, R.; Wang, H.; Yang, L.; Huang, J. Neuritin 1 expression in human normal tissues and its association with various human cancers. Int. J. Clin. Exp. Pathol. 2018, 11, 1956–1964. [Google Scholar]
- Han, D.; Qin, B.; Liu, G.; Liu, T.; Ji, G.; Wu, Y.; Yu, L. Characterization of neuritin as a novel angiogenic factor. Biochem. Biophys. Res. Commun. 2011, 415, 608–612. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, Y.; Zhou, J. Neuritin Promotes Bone Marrow-Derived Mesenchymal Stem Cell Migration to Treat Diabetic Peripheral Neuropathy. Mol. Neurobiol. 2022, 59, 6666–6683. [Google Scholar] [CrossRef] [PubMed]
- Kamada, S.; Ikeda, K.; Suzuki, T.; Sato, W.; Kitayama, S.; Kawakami, S.; Ichikawa, T.; Horie, K.; Inoue, S. Clinicopathological and Preclinical Patient-Derived Model Studies Define High Expression of NRN1 as a Diagnostic and Therapeutic Target for Clear Cell Renal Cell Carcinoma. Front. Oncol. 2021, 11, 758503. [Google Scholar] [CrossRef] [PubMed]
- Bosserhoff, A.K.; Schneider, N.; Ellmann, L.; Heinzerling, L.; Kuphal, S. The neurotrophin Neuritin1 (cpg15) is involved in melanoma migration, attachment independent growth, and vascular mimicry. Oncotarget 2016, 8, 1117–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, M.; Li, Y.; Zhong, C.; Li, Y.; Niu, J.; Gong, J. Overexpression of neuritin in gastric cancer. Oncol. Lett. 2015, 10, 3832–3836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Zhao, Y.; Wang, C.-G.; Fei, Z.; Wang, Y.; Li, L.; Li, L.; Zhen, H.-N. Neuritin expression and its relation with proliferation, apoptosis, and angiogenesis in human astrocytoma. Med. Oncol. 2010, 28, 907–912. [Google Scholar] [CrossRef]
- Gonzalez-Figueroa, P.; Roco, J.A.; Papa, I.; Villacís, L.N.; Stanley, M.; Linterman, M.A.; Dent, A.; Canete, P.F.; Vinuesa, C.G. Follicular regulatory T cells produce neuritin to regulate B cells. Cell 2021, 184, 1775–1789.e19. [Google Scholar] [CrossRef]
- Georgiev, H.; Peng, C.; Huggins, M.A.; Jameson, S.C.; Hogquist, K.A. Classical MHC expression by DP thymocytes impairs the selection of non-classical MHC restricted innate-like T cells. Nat. Commun. 2021, 12, 2308. [Google Scholar] [CrossRef]
- Schmidt, T.G.; Batz, L.; Bonet, L.; Carl, U.; Holzapfel, G.; Kiem, K.; Matulewicz, K.; Niermeier, D.; Schuchardt, I.; Stanar, K. Development of the Twin-Strep-tag® and its application for purification of recombinant proteins from cell culture supernatants. Protein Expr. Purif. 2013, 92, 54–61. [Google Scholar] [CrossRef]
- Georgiev, H.; Ravens, I.; Papadogianni, G.; Halle, S.; Malissen, B.; Loots, G.; Forster, R.; Bernhardt, G. Shared and Unique Features Distinguishing Follicular T Helper and Regulatory Cells of Peripheral Lymph Node and Peyer’s Patches. Front. Immunol. 2018, 9, 714. [Google Scholar] [CrossRef]
- Jameson, B.; Wolf, H. The antigenic index: A novel algorithm for predicting antigenic determinants. Bioinformatics 1988, 4, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Wei, Y.; Zhang, W.; Li, X.; Zhu, J.; Shan, L.; Liu, C.; Yuan, W.; Huang, J. Production of polyclonal antibody against human Neuritin and its application of immunodetection. Prep. Biochem. Biotechnol. 2019, 49, 209–214. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papadogianni, G.; Ravens, I.; Hassan, A.; Flatley, A.; Feederle, R.; Bernhardt, G.; Georgiev, H. Establishment and Functional Characterization of Murine Monoclonal Antibodies Recognizing Neuritin. Antibodies 2023, 12, 28. https://doi.org/10.3390/antib12020028
Papadogianni G, Ravens I, Hassan A, Flatley A, Feederle R, Bernhardt G, Georgiev H. Establishment and Functional Characterization of Murine Monoclonal Antibodies Recognizing Neuritin. Antibodies. 2023; 12(2):28. https://doi.org/10.3390/antib12020028
Chicago/Turabian StylePapadogianni, Georgia, Inga Ravens, Ahmed Hassan, Andrew Flatley, Regina Feederle, Günter Bernhardt, and Hristo Georgiev. 2023. "Establishment and Functional Characterization of Murine Monoclonal Antibodies Recognizing Neuritin" Antibodies 12, no. 2: 28. https://doi.org/10.3390/antib12020028
APA StylePapadogianni, G., Ravens, I., Hassan, A., Flatley, A., Feederle, R., Bernhardt, G., & Georgiev, H. (2023). Establishment and Functional Characterization of Murine Monoclonal Antibodies Recognizing Neuritin. Antibodies, 12(2), 28. https://doi.org/10.3390/antib12020028