
Development of a Novel Anti-CD44 Variant 8 Monoclonal Antibody C44Mab-94 against Gastric Carcinomas

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
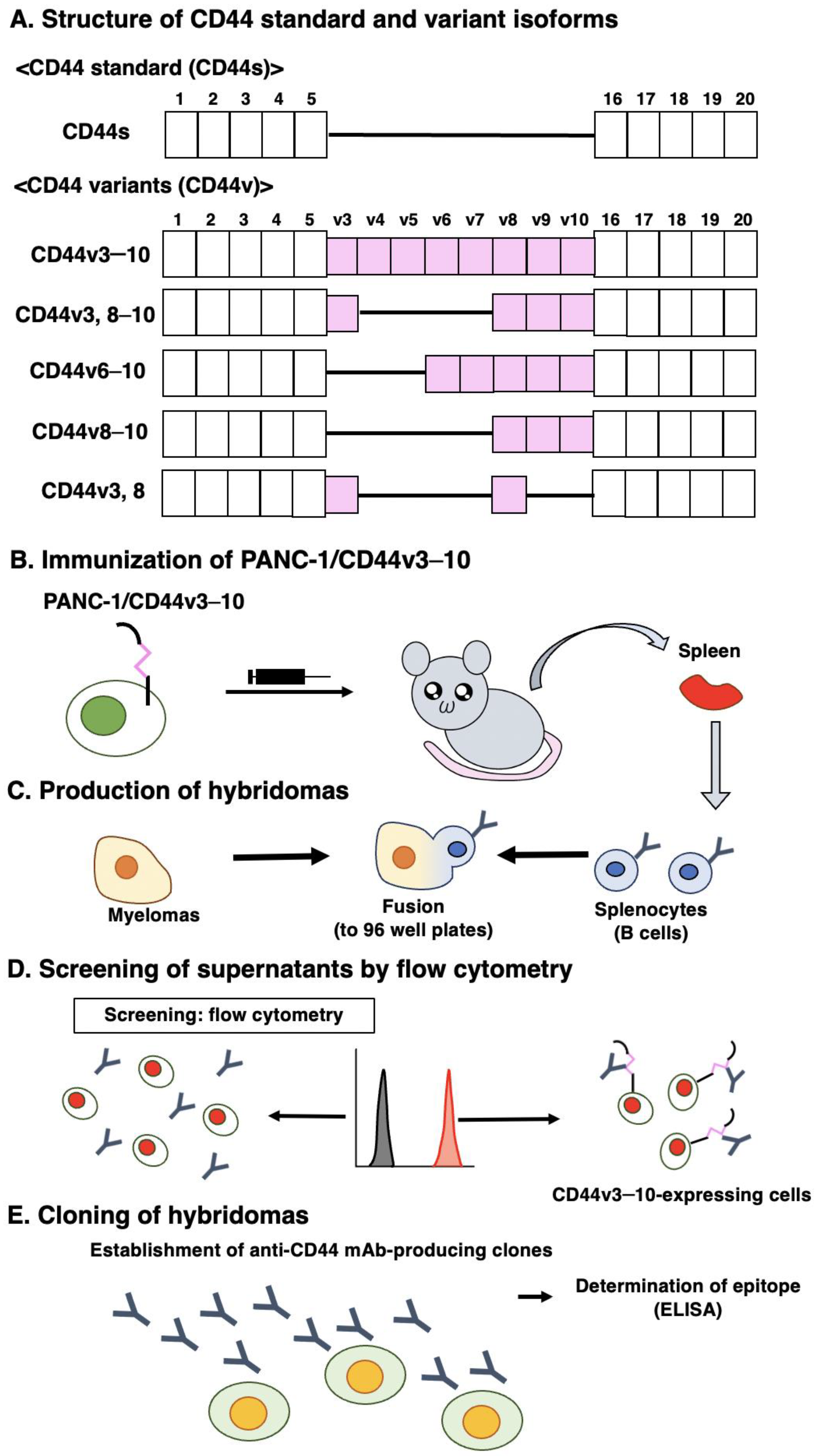
2.1. Cell Lines
2.2. Construction of Plasmid DNA and Establishment of Stable Transfectants
2.3. Production of Hybridomas
2.4. Enzyme-Linked Immunosorbent Assay (ELISA)
2.5. Flow Cytometry
2.6. Western Blot Analysis
2.7. Immunohistochemical Analysis
3. Results
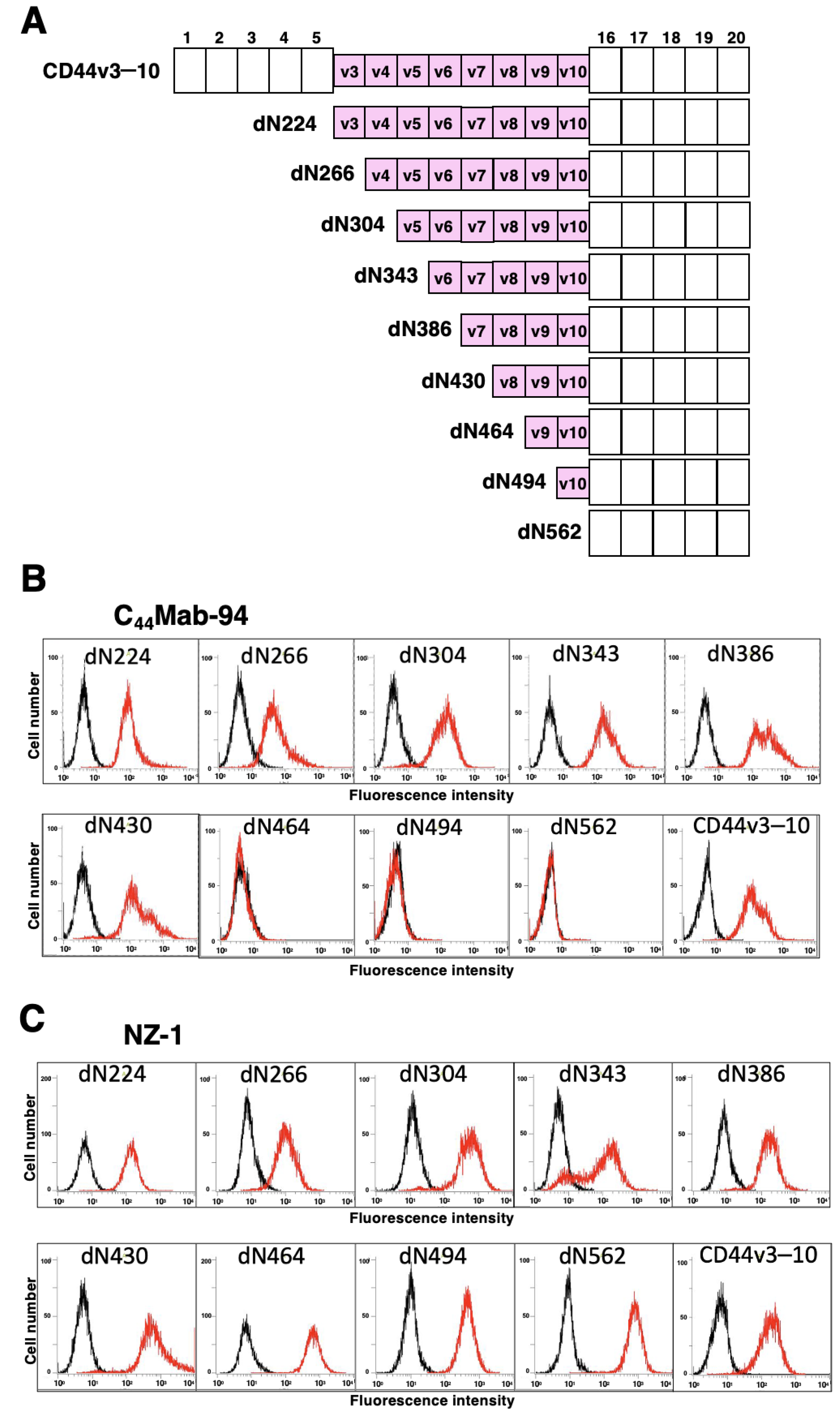
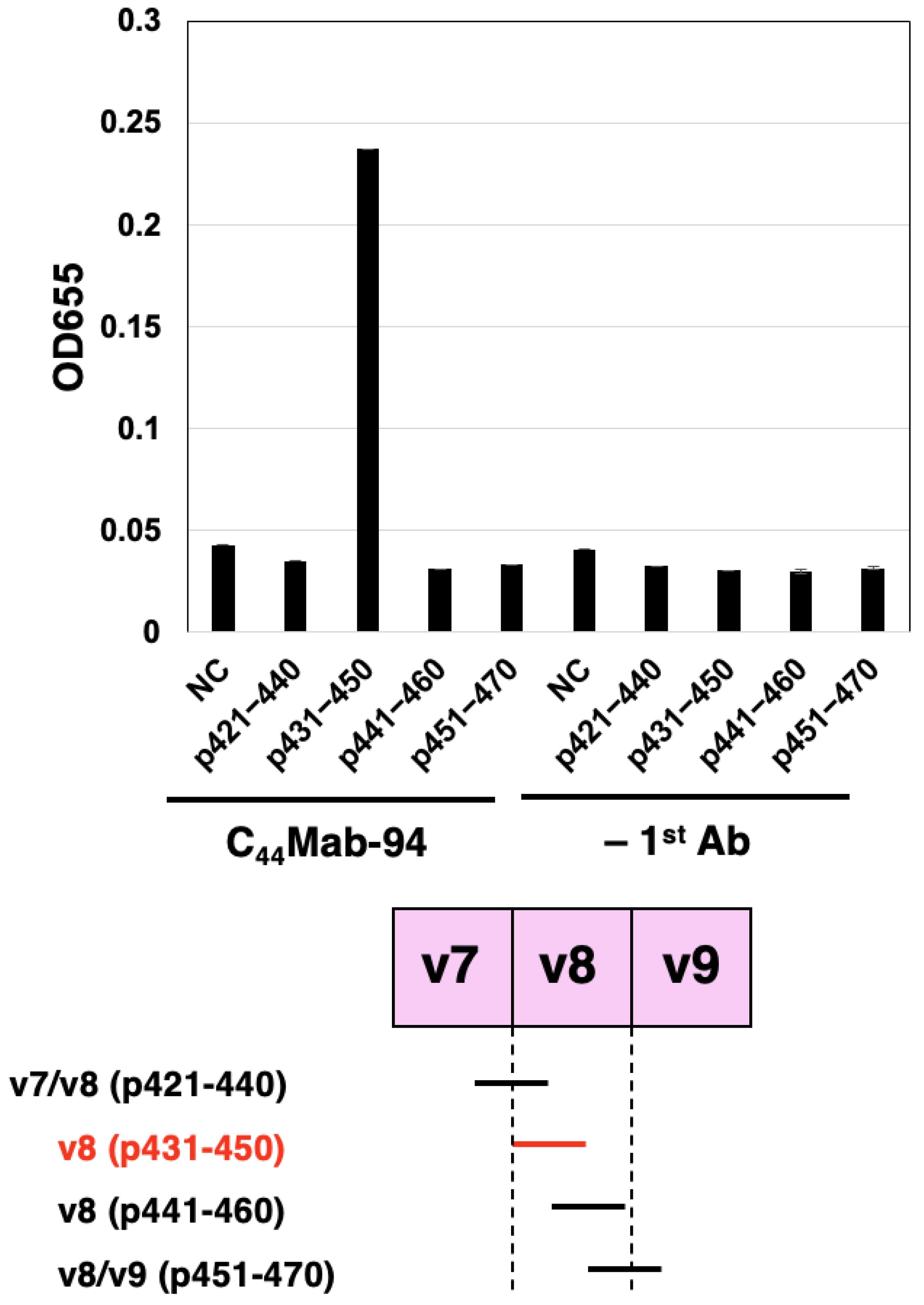
3.1. Establishment of an Anti-CD44v8 mAb, C44Mab-94
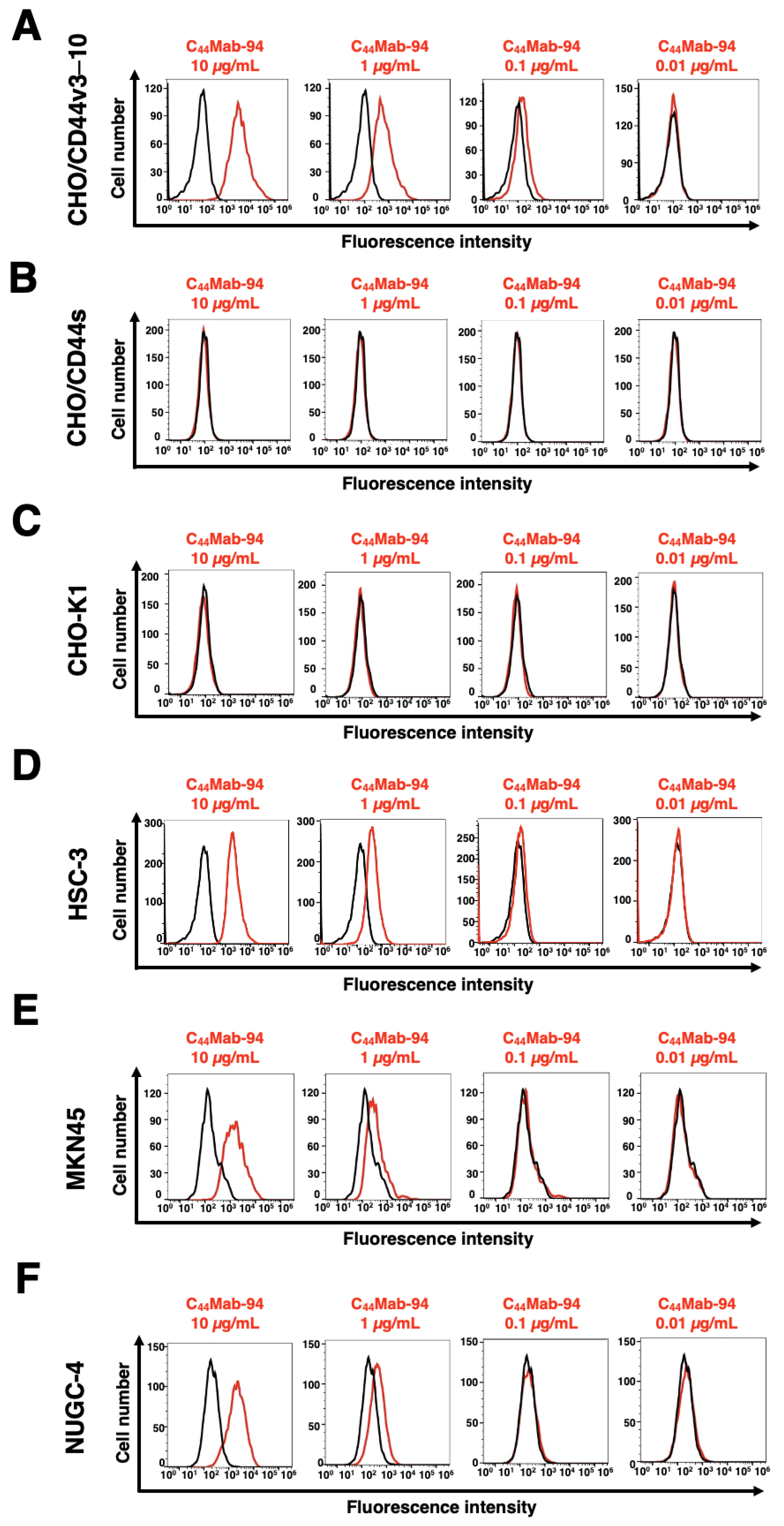
3.2. Flow Cytometric Analysis of C44Mab-94 against CD44-Expressing Cells
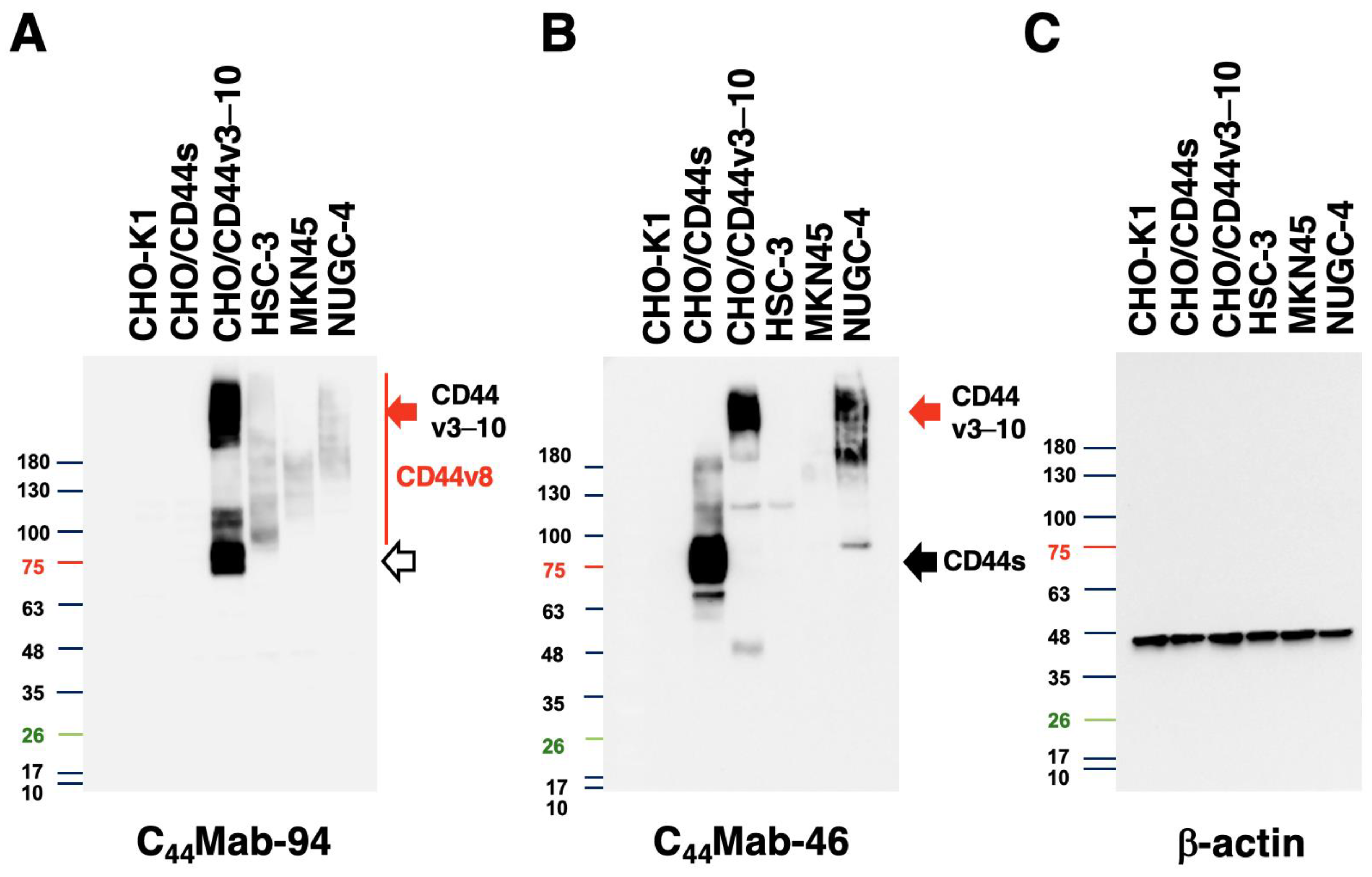
3.3. Western Blot Analysis
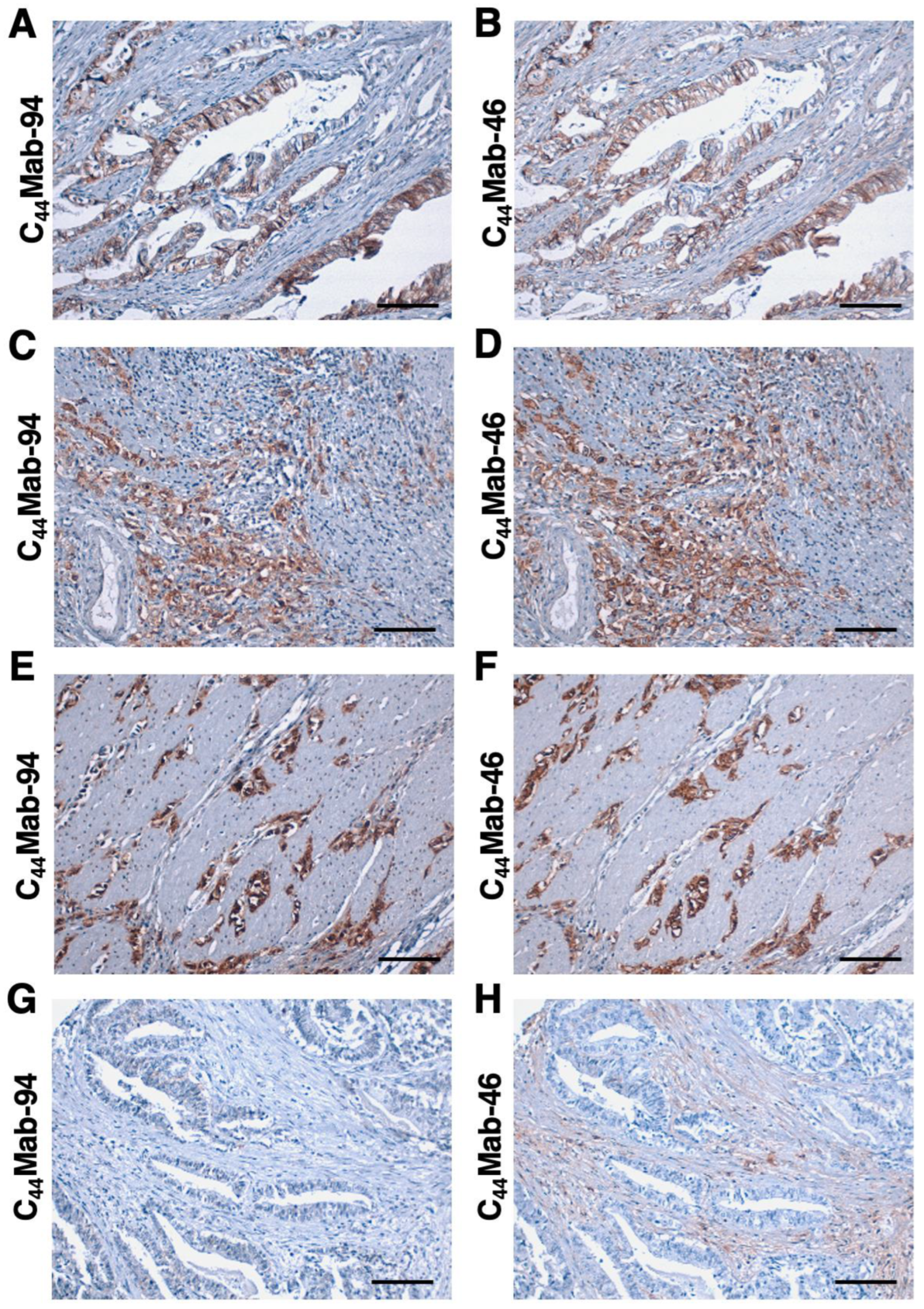
3.4. Immunohistochemical Analysis Using C44Mab-94 against Tumor Tissues
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- López, M.J.; Carbajal, J.; Alfaro, A.L.; Saravia, L.G.; Zanabria, D.; Araujo, J.M.; Quispe, L.; Zevallos, A.; Buleje, J.L.; Cho, C.E.; et al. Characteristics of gastric cancer around the world. Crit. Rev. Oncol. Hematol. 2023, 181, 103841. [Google Scholar] [CrossRef] [PubMed]
- Lauren, P. The two histological main types of gastric carcinoma: Diffuse and so-called intestinal-type carcinoma. an attempt at a histo-clinical classification. Acta Pathol. Microbiol. Scand. 1965, 64, 31–49. [Google Scholar] [CrossRef] [PubMed]
- Kushima, R. The updated WHO classification of digestive system tumours-gastric adenocarcinoma and dysplasia. Pathologe 2022, 43, 8–15. [Google Scholar] [CrossRef]
- Cristescu, R.; Lee, J.; Nebozhyn, M.; Kim, K.M.; Ting, J.C.; Wong, S.S.; Liu, J.; Yue, Y.G.; Wang, J.; Yu, K.; et al. Molecular analysis of gastric cancer identifies subtypes associated with distinct clinical outcomes. Nat. Med. 2015, 21, 449–456. [Google Scholar] [CrossRef]
- The Cancer Genome Atlas Research Network. Comprehensive molecular characterization of gastric adenocarcinoma. Nature 2014, 513, 202–209. [Google Scholar] [CrossRef] [Green Version]
- Shitara, K.; Bang, Y.J.; Iwasa, S.; Sugimoto, N.; Ryu, M.H.; Sakai, D.; Chung, H.C.; Kawakami, H.; Yabusaki, H.; Lee, J.; et al. Trastuzumab Deruxtecan in Previously Treated HER2-Positive Gastric Cancer. N. Engl. J. Med. 2020, 382, 2419–2430. [Google Scholar] [CrossRef]
- Kang, Y.K.; Boku, N.; Satoh, T.; Ryu, M.H.; Chao, Y.; Kato, K.; Chung, H.C.; Chen, J.S.; Muro, K.; Kang, W.K.; et al. Nivolumab in patients with advanced gastric or gastro-oesophageal junction cancer refractory to, or intolerant of, at least two previous chemotherapy regimens (ONO-4538-12, ATTRACTION-2): A randomised, double-blind, placebo-controlled, phase 3 trial. Lancet 2017, 390, 2461–2471. [Google Scholar] [CrossRef]
- Monster, J.L.; Kemp, L.J.S.; Gloerich, M.; van der Post, R.S. Diffuse gastric cancer: Emerging mechanisms of tumor initiation and progression. Biochim. Biophys. Acta Rev. Cancer 2022, 1877, 188719. [Google Scholar] [CrossRef]
- Tanaka, Y.; Chiwaki, F.; Kojima, S.; Kawazu, M.; Komatsu, M.; Ueno, T.; Inoue, S.; Sekine, S.; Matsusaki, K.; Matsushita, H.; et al. Multi-omic profiling of peritoneal metastases in gastric cancer identifies molecular subtypes and therapeutic vulnerabilities. Nat. Cancer 2021, 2, 962–977. [Google Scholar] [CrossRef]
- Tsao, L.C.; Force, J.; Hartman, Z.C. Mechanisms of Therapeutic Antitumor Monoclonal Antibodies. Cancer Res. 2021, 81, 4641–4651. [Google Scholar] [CrossRef] [PubMed]
- Günthert, U.; Hofmann, M.; Rudy, W.; Reber, S.; Zöller, M.; Haussmann, I.; Matzku, S.; Wenzel, A.; Ponta, H.; Herrlich, P. A new variant of glycoprotein CD44 confers metastatic potential to rat carcinoma cells. Cell 1991, 65, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Ponta, H.; Sherman, L.; Herrlich, P.A. CD44: From adhesion molecules to signalling regulators. Nat. Rev. Mol. Cell Biol. 2003, 4, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Zuo, X.; Wei, D. Concise Review: Emerging Role of CD44 in Cancer Stem Cells: A Promising Biomarker and Therapeutic Target. Stem Cells Transl. Med. 2015, 4, 1033–1043. [Google Scholar] [CrossRef]
- Chen, C.; Zhao, S.; Karnad, A.; Freeman, J.W. The biology and role of CD44 in cancer progression: Therapeutic implications. J. Hematol. Oncol. 2018, 11, 64. [Google Scholar] [CrossRef] [Green Version]
- Liao, C.; Wang, Q.; An, J.; Chen, J.; Li, X.; Long, Q.; Xiao, L.; Guan, X.; Liu, J. CD44 Glycosylation as a Therapeutic Target in Oncology. Front. Oncol. 2022, 12, 883831. [Google Scholar] [CrossRef] [PubMed]
- Mishra, M.N.; Chandavarkar, V.; Sharma, R.; Bhargava, D. Structure, function and role of CD44 in neoplasia. J. Oral Maxillofac. Pathol. 2019, 23, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Slevin, M.; Krupinski, J.; Gaffney, J.; Matou, S.; West, D.; Delisser, H.; Savani, R.C.; Kumar, S. Hyaluronan-mediated angiogenesis in vascular disease: Uncovering RHAMM and CD44 receptor signaling pathways. Matrix Biol. 2007, 26, 58–68. [Google Scholar] [CrossRef]
- Hassn Mesrati, M.; Syafruddin, S.E.; Mohtar, M.A.; Syahir, A. CD44: A Multifunctional Mediator of Cancer Progression. Biomolecules 2021, 11, 1850. [Google Scholar] [CrossRef]
- Guo, Q.; Yang, C.; Gao, F. The state of CD44 activation in cancer progression and therapeutic targeting. FEBS J. 2021, 289, 7970–7986. [Google Scholar] [CrossRef]
- Zöller, M. CD44: Can a cancer-initiating cell profit from an abundantly expressed molecule? Nat. Rev. Cancer 2011, 11, 254–267. [Google Scholar] [CrossRef]
- Jackson, D.G.; Bell, J.I.; Dickinson, R.; Timans, J.; Shields, J.; Whittle, N. Proteoglycan forms of the lymphocyte homing receptor CD44 are alternatively spliced variants containing the v3 exon. J. Cell Biol. 1995, 128, 673–685. [Google Scholar] [CrossRef]
- Bennett, K.L.; Jackson, D.G.; Simon, J.C.; Tanczos, E.; Peach, R.; Modrell, B.; Stamenkovic, I.; Plowman, G.; Aruffo, A. CD44 isoforms containing exon V3 are responsible for the presentation of heparin-binding growth factor. J. Cell Biol. 1995, 128, 687–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matzke, A.; Sargsyan, V.; Holtmann, B.; Aramuni, G.; Asan, E.; Sendtner, M.; Pace, G.; Howells, N.; Zhang, W.; Ponta, H.; et al. Haploinsufficiency of c-Met in cd44−/− mice identifies a collaboration of CD44 and c-Met in vivo. Mol. Cell. Biol. 2007, 27, 8797–8806. [Google Scholar] [CrossRef] [Green Version]
- Orian-Rousseau, V.; Chen, L.; Sleeman, J.P.; Herrlich, P.; Ponta, H. CD44 is required for two consecutive steps in HGF/c-Met signaling. Genes Dev. 2002, 16, 3074–3086. [Google Scholar] [CrossRef] [Green Version]
- Ishimoto, T.; Nagano, O.; Yae, T.; Tamada, M.; Motohara, T.; Oshima, H.; Oshima, M.; Ikeda, T.; Asaba, R.; Yagi, H.; et al. CD44 variant regulates redox status in cancer cells by stabilizing the xCT subunit of system xc− and thereby promotes tumor growth. Cancer Cell 2011, 19, 387–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagiwara, M.; Kikuchi, E.; Tanaka, N.; Kosaka, T.; Mikami, S.; Saya, H.; Oya, M. Variant isoforms of CD44 involves acquisition of chemoresistance to cisplatin and has potential as a novel indicator for identifying a cisplatin-resistant population in urothelial cancer. BMC Cancer 2018, 18, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagami, T.; Yamade, M.; Suzuki, T.; Uotani, T.; Tani, S.; Hamaya, Y.; Iwaizumi, M.; Osawa, S.; Sugimoto, K.; Baba, S.; et al. High expression level of CD44v8-10 in cancer stem-like cells is associated with poor prognosis in esophageal squamous cell carcinoma patients treated with chemoradiotherapy. Oncotarget 2018, 9, 34876–34888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, S.; Itai, S.; Nakamura, T.; Yanaka, M.; Kaneko, M.K.; Kato, Y. Detection of high CD44 expression in oral cancers using the novel monoclonal antibody, C44Mab-5. Biochem. Biophys. Rep. 2018, 14, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Goto, N.; Suzuki, H.; Tanaka, T.; Asano, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-CD44 Monoclonal Antibody for Multiple Applications against Esophageal Squamous Cell Carcinomas. Int. J. Mol. Sci. 2022, 23, 5535. [Google Scholar] [CrossRef]
- Takei, J.; Asano, T.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Epitope Mapping of the Anti-CD44 Monoclonal Antibody (C44Mab-46) Using Alanine-Scanning Mutagenesis and Surface Plasmon Resonance. Monoclon. Antib. Immunodiagn. Immunother. 2021, 40, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Asano, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Epitope Mapping System: RIEDL Insertion for Epitope Mapping Method. Monoclon. Antib. Immunodiagn. Immunother. 2021, 40, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Takei, J.; Kaneko, M.K.; Ohishi, T.; Hosono, H.; Nakamura, T.; Yanaka, M.; Sano, M.; Asano, T.; Sayama, Y.; Kawada, M.; et al. A defucosylated antiCD44 monoclonal antibody 5mG2af exerts antitumor effects in mouse xenograft models of oral squamous cell carcinoma. Oncol. Rep. 2020, 44, 1949–1960. [Google Scholar] [CrossRef]
- Suzuki, H.; Tanaka, T.; Goto, N.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-CD44 Variant 4 Monoclonal Antibody C44Mab-108 for Immunohistochemistry. Curr. Issues Mol. Biol. 2023, 45, 121. [Google Scholar] [CrossRef]
- Kudo, Y.; Suzuki, H.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-CD44 variant 5 Monoclonal Antibody C44Mab-3 for Multiple Applications against Pancreatic Carcinomas. Antibodies 2023, 12, 31. [Google Scholar] [CrossRef]
- Ejima, R.; Suzuki, H.; Tanaka, T.; Asano, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-CD44 Variant 6 Monoclonal Antibody C44Mab-9 for Multiple Applications against Colorectal Carcinomas. Int. J. Mol. Sci. 2023, 24, 4007. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Ozawa, K.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-CD44 Variant 7/8 Monoclonal Antibody, C44Mab-34, for Multiple Applications against Oral Carcinomas. Biomedicines 2023, 11, 1099. [Google Scholar] [CrossRef] [PubMed]
- Tawara, M.; Suzuki, H.; Goto, N.; Tanaka, T.; Kaneko, M.K.; Kato, Y. A Novel Anti-CD44 Variant 9 Monoclonal Antibody C44Mab-1 was Developed for Immunohistochemical Analyses Against Colorectal Cancers. Curr. Issues Mol. Biol. 2023, 45, 238. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Iimori, M.; Edahiro, K.; Fujimoto, Y.; Matsuoka, K.; Oki, E.; Maehara, Y.; Mori, M.; Kitao, H. CD44v3,8-10 is essential for Slug-dependent vimentin gene expression to acquire TGF-β1-induced tumor cell motility. Cancer Sci. 2022, 113, 2654–2667. [Google Scholar] [CrossRef]
- Ludwig, N.; Szczepanski, M.J.; Gluszko, A.; Szafarowski, T.; Azambuja, J.H.; Dolg, L.; Gellrich, N.C.; Kampmann, A.; Whiteside, T.L.; Zimmerer, R.M. CD44(+) tumor cells promote early angiogenesis in head and neck squamous cell carcinoma. Cancer Lett. 2019, 467, 85–95. [Google Scholar] [CrossRef]
- Heider, K.H.; Sproll, M.; Susani, S.; Patzelt, E.; Beaumier, P.; Ostermann, E.; Ahorn, H.; Adolf, G.R. Characterization of a high-affinity monoclonal antibody specific for CD44v6 as candidate for immunotherapy of squamous cell carcinomas. Cancer Immunol. Immunother. 1996, 43, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Heider, K.H.; Mulder, J.W.; Ostermann, E.; Susani, S.; Patzelt, E.; Pals, S.T.; Adolf, G.R. Splice variants of the cell surface glycoprotein CD44 associated with metastatic tumour cells are expressed in normal tissues of humans and cynomolgus monkeys. Eur. J. Cancer 1995, 31, 2385–2391. [Google Scholar] [CrossRef] [PubMed]
- Dall, P.; Hekele, A.; Ikenberg, H.; Göppinger, A.; Bauknecht, T.; Pfleiderer, A.; Moll, J.; Hofmann, M.; Ponta, H.; Herrlich, P. Increasing incidence of CD44v7/8 epitope expression during uterine cervical carcinogenesis. Int. J. Cancer 1996, 69, 79–85. [Google Scholar] [CrossRef]
- Woerner, S.M.; Givehchian, M.; Dürst, M.; Schneider, A.; Costa, S.; Melsheimer, P.; Lacroix, J.; Zöller, M.; Doeberitz, M.K. Expression of CD44 splice variants in normal, dysplastic, and neoplastic cervical epithelium. Clin. Cancer Res. 1995, 1, 1125–1132. [Google Scholar] [PubMed]
- Gansauge, F.; Gansauge, S.; Zobywalski, A.; Scharnweber, C.; Link, K.H.; Nussler, A.K.; Beger, H.G. Differential expression of CD44 splice variants in human pancreatic adenocarcinoma and in normal pancreas. Cancer Res. 1995, 55, 5499–5503. [Google Scholar] [PubMed]
- Verel, I.; Heider, K.H.; Siegmund, M.; Ostermann, E.; Patzelt, E.; Sproll, M.; Snow, G.B.; Adolf, G.R.; van Dongen, G.A. Tumor targeting properties of monoclonal antibodies with different affinity for target antigen CD44V6 in nude mice bearing head-and-neck cancer xenografts. Int. J. Cancer 2002, 99, 396–402. [Google Scholar] [CrossRef]
- Riechelmann, H.; Sauter, A.; Golze, W.; Hanft, G.; Schroen, C.; Hoermann, K.; Erhardt, T.; Gronau, S. Phase I trial with the CD44v6-targeting immunoconjugate bivatuzumab mertansine in head and neck squamous cell carcinoma. Oral Oncol. 2008, 44, 823–829. [Google Scholar] [CrossRef]
- Tijink, B.M.; Buter, J.; de Bree, R.; Giaccone, G.; Lang, M.S.; Staab, A.; Leemans, C.R.; van Dongen, G.A. A phase I dose escalation study with anti-CD44v6 bivatuzumab mertansine in patients with incurable squamous cell carcinoma of the head and neck or esophagus. Clin. Cancer Res. 2006, 12, 6064–6072. [Google Scholar] [CrossRef] [Green Version]
- Fox, S.B.; Fawcett, J.; Jackson, D.G.; Collins, I.; Gatter, K.C.; Harris, A.L.; Gearing, A.; Simmons, D.L. Normal human tissues, in addition to some tumors, express multiple different CD44 isoforms. Cancer Res. 1994, 54, 4539–4546. [Google Scholar]
- Mereiter, S.; Martins, Á.M.; Gomes, C.; Balmaña, M.; Macedo, J.A.; Polom, K.; Roviello, F.; Magalhães, A.; Reis, C.A. O-glycan truncation enhances cancer-related functions of CD44 in gastric cancer. FEBS Lett. 2019, 593, 1675–1689. [Google Scholar] [CrossRef]
- Hirata, K.; Suzuki, H.; Imaeda, H.; Matsuzaki, J.; Tsugawa, H.; Nagano, O.; Asakura, K.; Saya, H.; Hibi, T. CD44 variant 9 expression in primary early gastric cancer as a predictive marker for recurrence. Br. J. Cancer 2013, 109, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Shitara, K.; Doi, T.; Nagano, O.; Imamura, C.K.; Ozeki, T.; Ishii, Y.; Tsuchihashi, K.; Takahashi, S.; Nakajima, T.E.; Hironaka, S.; et al. Dose-escalation study for the targeting of CD44v+ cancer stem cells by sulfasalazine in patients with advanced gastric cancer (EPOC1205). Gastric. Cancer 2017, 20, 341–349. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Suzuki, H.; Ohishi, T.; Asano, T.; Tanaka, T.; Yanaka, M.; Nakamura, T.; Yoshikawa, T.; Kawada, M.; Kaneko, M.K.; et al. Antitumor activities of a defucosylated anti-EpCAM monoclonal antibody in colorectal carcinoma xenograft models. Int. J. Mol. Med. 2023, 51, 18. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Huang, H.; Tang, Y.; Li, Q.; Wang, J.; Li, D.; Zhong, Z.; Zou, P.; You, Y.; Cao, Y.; et al. CD44v6 chimeric antigen receptor T cell specificity towards AML with FLT3 or DNMT3A mutations. Clin. Transl. Med. 2022, 12, e1043. [Google Scholar] [CrossRef]
- Greco, B.; Malacarne, V.; De Girardi, F.; Scotti, G.M.; Manfredi, F.; Angelino, E.; Sirini, C.; Camisa, B.; Falcone, L.; Moresco, M.A.; et al. Disrupting N-glycan expression on tumor cells boosts chimeric antigen receptor T cell efficacy against solid malignancies. Sci. Transl. Med. 2022, 14, eabg3072. [Google Scholar] [CrossRef] [PubMed]
- Porcellini, S.; Asperti, C.; Corna, S.; Cicoria, E.; Valtolina, V.; Stornaiuolo, A.; Valentinis, B.; Bordignon, C.; Traversari, C. CAR T Cells Redirected to CD44v6 Control Tumor Growth in Lung and Ovary Adenocarcinoma Bearing Mice. Front. Immunol. 2020, 11, 99. [Google Scholar] [CrossRef] [Green Version]
- Holm, F.; Hellqvist, E.; Mason, C.N.; Ali, S.A.; Delos-Santos, N.; Barrett, C.L.; Chun, H.J.; Minden, M.D.; Moore, R.A.; Marra, M.A.; et al. Reversion to an embryonic alternative splicing program enhances leukemia stem cell self-renewal. Proc. Natl. Acad. Sci. USA 2015, 112, 15444–15449. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Kaneko, M.K. A cancer-specific monoclonal antibody recognizes the aberrantly glycosylated podoplanin. Sci. Rep. 2014, 4, 5924. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, M.K.; Ohishi, T.; Kawada, M.; Kato, Y. A cancer-specific anti-podocalyxin monoclonal antibody (60-mG2a-f) exerts antitumor effects in mouse xenograft models of pancreatic carcinoma. Biochem. Biophys. Rep. 2020, 24, 100826. [Google Scholar] [CrossRef]
- Suzuki, H.; Kaneko, M.K.; Kato, Y. Roles of Podoplanin in Malignant Progression of Tumor. Cells 2022, 11, 575. [Google Scholar] [CrossRef]
- Chalise, L.; Kato, A.; Ohno, M.; Maeda, S.; Yamamichi, A.; Kuramitsu, S.; Shiina, S.; Takahashi, H.; Ozone, S.; Yamaguchi, J.; et al. Efficacy of cancer-specific anti-podoplanin CAR-T cells and oncolytic herpes virus G47Δ combination therapy against glioblastoma. Mol. Ther.-Oncolytics 2022, 26, 265–274. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Age | Sex | Pathology Diagnosis | TNM | Grade | Stage | C44Mab-94 | C44Mab-46 |
|---|---|---|---|---|---|---|---|---|
| 1 | 55 | F | Adenocarcinoma | T2N0M0 | 1 | IB | - | + |
| 2 | 51 | F | Adenocarcinoma | T2N0M0 | - | IB | - | - |
| 3 | 71 | M | Adenocarcinoma | T3N1M0 | 1 | IIB | - | ++ |
| 4 | 63 | M | Adenocarcinoma | T3N0M0 | 1 | IIA | - | - |
| 5 | 61 | M | Adenocarcinoma | T2N0M0 | 1 | IB | - | - |
| 6 | 61 | M | Adenocarcinoma | T2N0M0 | 1 | IB | - | + |
| 7 | 60 | M | Adenocarcinoma | T3N2M0 | 1 | IIIA | - | - |
| 8 | 54 | M | Adenocarcinoma | T3N2M0 | 1 | IIIA | + | ++ |
| 9 | 46 | F | Adenocarcinoma | T3N0M0 | 1 | IIA | - | + |
| 10 | 66 | M | Mucinous adenocarcinoma | T3N0M0 | 2–3 | IIA | - | + |
| 11 | 56 | M | Adenocarcinoma | T2N0M0 | 2 | IB | ++ | + |
| 12 | 52 | F | Adenocarcinoma | T3N0M0 | 2 | IIA | + | + |
| 13 | 70 | M | Adenocarcinoma | T3N0M0 | 2 | IIA | - | + |
| 14 | 71 | M | Adenocarcinoma | T2N0M0 | 2 | IB | - | ++ |
| 15 | 61 | M | Adenocarcinoma | T3N0M0 | 2 | IIA | - | - |
| 16 | 75 | M | Adenocarcinoma | T3N1M0 | 2 | IIB | - | - |
| 17 | 72 | F | Adenocarcinoma | T3N0M0 | 2 | IIA | + | + |
| 18 | 60 | M | Adenocarcinoma | T3N0M0 | 2 | IIA | - | - |
| 19 | 63 | F | Adenocarcinoma | T3N0M0 | 2 | IIA | - | - |
| 20 | 69 | M | Adenocarcinoma | T2N0M0 | 2 | IB | - | - |
| 21 | 54 | F | Adenocarcinoma | T3N0M0 | 2 | IIA | - | - |
| 22 | 50 | F | Adenocarcinoma | T3N0M0 | 3 | IIA | - | - |
| 23 | 64 | M | Adenocarcinoma | T3N0M0 | 3 | IIA | - | ++ |
| 24 | 59 | M | Adenocarcinoma | T2N0M0 | 2 | IB | ++ | +++ |
| 25 | 59 | M | Adenocarcinoma | T2N0M0 | 2 | IB | - | - |
| 26 | 44 | M | Adenocarcinoma | T3N0M0 | 2 | IIA | - | - |
| 27 | 76 | M | Adenocarcinoma | T3N0M0 | 2 | IIA | - | + |
| 28 | 56 | M | Adenocarcinoma | T3N0M0 | 2 | IIA | - | + |
| 29 | 56 | M | Adenocarcinoma | T2N0M0 | 2 | IB | + | + |
| 30 | 58 | M | Adenocarcinoma | T3N0M0 | 2 | IIA | - | + |
| 31 | 94 | M | Adenocarcinoma | T2N0M0 | 2 | IB | + | + |
| 32 | 56 | F | Adenocarcinoma | T2N0M0 | 3 | IB | + | + |
| 33 | 56 | M | Adenocarcinoma | T4N1M0 | 3 | IIIA | + | + |
| 34 | 51 | F | Adenocarcinoma | T3N0M0 | 2 | IIA | + | + |
| 35 | 67 | M | Adenocarcinoma | T3N0M0 | 2 | IIA | + | + |
| 36 | 53 | M | Adenocarcinoma | T3N0M0 | 2–3 | IIA | + | + |
| 37 | 48 | F | Adenocarcinoma | T2N1M0 | 3 | IIA | ++ | ++ |
| 38 | 58 | M | Adenocarcinoma | T2N0M0 | 2 | IB | - | - |
| 39 | 61 | M | Adenocarcinoma | T2N0M0 | 3 | IB | - | + |
| 40 | 62 | M | Adenocarcinoma | T2N0M0 | 3 | IB | - | + |
| 41 | 65 | M | Adenocarcinoma | T2N0M0 | 3 | IB | + | + |
| 42 | 47 | F | Adenocarcinoma | T3N1M0 | 3 | IIB | - | - |
| 43 | 65 | M | Adenocarcinoma | T2N0M0 | - | IB | - | - |
| 44 | 52 | F | Adenocarcinoma | T2N0M0 | 3 | IB | - | - |
| 45 | 72 | M | Adenocarcinoma | T2N0M0 | 3 | IB | - | + |
| 46 | 68 | F | Adenocarcinoma | T3N0M0 | 3 | IIA | - | ++ |
| 47 | 56 | M | Adenocarcinoma | T3N0M0 | 3 | IIA | ++ | ++ |
| 48 | 59 | M | Adenocarcinoma | T3N1M0 | 3 | IIB | + | + |
| 49 | 62 | M | Adenocarcinoma | T3N1M0 | 3 | IIB | - | - |
| 50 | 60 | M | Adenocarcinoma | T3N1M0 | 3 | IIB | + | +++ |
| 51 | 64 | M | Adenocarcinoma | T2N0M0 | 3 | IB | + | + |
| 52 | 69 | M | Adenocarcinoma | T2N0M0 | 3 | IB | + | ++ |
| 53 | 75 | M | Adenocarcinoma | T2N0M0 | 3 | IB | + | ++ |
| 54 | 48 | M | Adenocarcinoma | T2N0M0 | 3 | IB | - | - |
| 55 | 59 | M | Adenocarcinoma | T2N0M0 | 3 | IB | - | - |
| 56 | 64 | M | Adenocarcinoma | T3N0M0 | 3 | IIA | + | + |
| 57 | 55 | M | Adenocarcinoma | T2N0M0 | 3 | IB | ++ | ++ |
| 58 | 58 | M | Adenocarcinoma | T3N0M0 | 3 | IIA | + | + |
| 59 | 64 | M | Adenocarcinoma | T3N0M0 | 3 | IIA | + | + |
| 60 | 67 | M | Adenocarcinoma | T3N1M0 | 3 | IIB | - | + |
| 61 | 49 | M | Adenocarcinoma | T2N0M0 | 3 | IB | - | - |
| 62 | 35 | M | Adenocarcinoma | T3N1M0 | 3 | IIB | - | - |
| 63 | 45 | F | Adenocarcinoma | T4N0M1 | 3 | IV | + | ++ |
| 64 | 43 | M | Adenocarcinoma | T2N0M0 | 3 | IB | - | + |
| 65 | 56 | M | Adenocarcinoma | T2N0M0 | 3 | IB | - | - |
| 66 | 66 | M | Adenocarcinoma | T2N0M0 | 3 | IB | + | + |
| 67 | 60 | M | Adenocarcinoma | T3N0M0 | 3 | IIA | - | - |
| 68 | 74 | M | Adenocarcinoma | T2N0M0 | 3 | IB | + | ++ |
| 69 | 58 | M | Adenocarcinoma | T2N0M0 | 3 | IB | - | - |
| 70 | 68 | M | Mucinous adenocarcinoma | T2N0M0 | 2 | IB | + | + |
| 71 | 50 | M | Mucinous adenocarcinoma | T3N0M0 | 3 | IIA | - | - |
| 72 | 51 | M | Papillary adenocarcinoma | T2N0M0 | 2 | IB | - | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, H.; Goto, N.; Tanaka, T.; Ouchida, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-CD44 Variant 8 Monoclonal Antibody C44Mab-94 against Gastric Carcinomas. Antibodies 2023, 12, 45. https://doi.org/10.3390/antib12030045
Suzuki H, Goto N, Tanaka T, Ouchida T, Kaneko MK, Kato Y. Development of a Novel Anti-CD44 Variant 8 Monoclonal Antibody C44Mab-94 against Gastric Carcinomas. Antibodies. 2023; 12(3):45. https://doi.org/10.3390/antib12030045
Chicago/Turabian StyleSuzuki, Hiroyuki, Nohara Goto, Tomohiro Tanaka, Tsunenori Ouchida, Mika K. Kaneko, and Yukinari Kato. 2023. "Development of a Novel Anti-CD44 Variant 8 Monoclonal Antibody C44Mab-94 against Gastric Carcinomas" Antibodies 12, no. 3: 45. https://doi.org/10.3390/antib12030045
APA StyleSuzuki, H., Goto, N., Tanaka, T., Ouchida, T., Kaneko, M. K., & Kato, Y. (2023). Development of a Novel Anti-CD44 Variant 8 Monoclonal Antibody C44Mab-94 against Gastric Carcinomas. Antibodies, 12(3), 45. https://doi.org/10.3390/antib12030045