
Exploring Parametric and Mechanistic Differences between Expi293FTM and ExpiCHO-STM Cells for Transient Antibody Production Optimization

 , , ,
, , , Highlights
- Established the most optimal transfection parameters for antibody production in the TGE cell–host Expi293FTM and ExpiCHO-STM with ExpiFectamineTM and polyethylenimine.
- Identified the mechanistic differences between Expi293FTM and ExpiCHO-STM with regard to DNA/transfection agent complexation formation and cellular uptake trafficking.
- Uncovered the expression preferences towards antibody light-chain isotypes in Expi293FTM and ExpiCHO-STM.
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cell Culture
2.3. Plasmids and Plasmid Preparation
2.4. Transient Transfection Procedures for Expi293FTM and ExpiCHO-STM
2.5. Expression Titer Determination and Quality Control for Expressed Antibodies
2.6. Liquid Chromatography (LC)–Mass Spectrometry (MS) Protein Sample Preparation
2.7. LC-MS Instrument Methods
2.8. Dynamic Light Scattering
3. Results
3.1. The Volume of Opti-MEMTM Dilution Buffer for DNAs and Transfection Reagents Was Critical for the TGE Protein Productivity in Expi293FTM and ExpiCHO-STM Cells
3.2. Expi293FTM and ExpiCHO-STM Cells Responded Differently to PEI/DNA Ratios
3.3. ExpiCHO-STM Cells Preferred a Shorter Complex Formation Time Than That of Expi293FTM Cells
3.4. The Size of the DNA Complexes with Either ExpiFectamine™CHO or PEI Increased as a Function of the Incubation Time
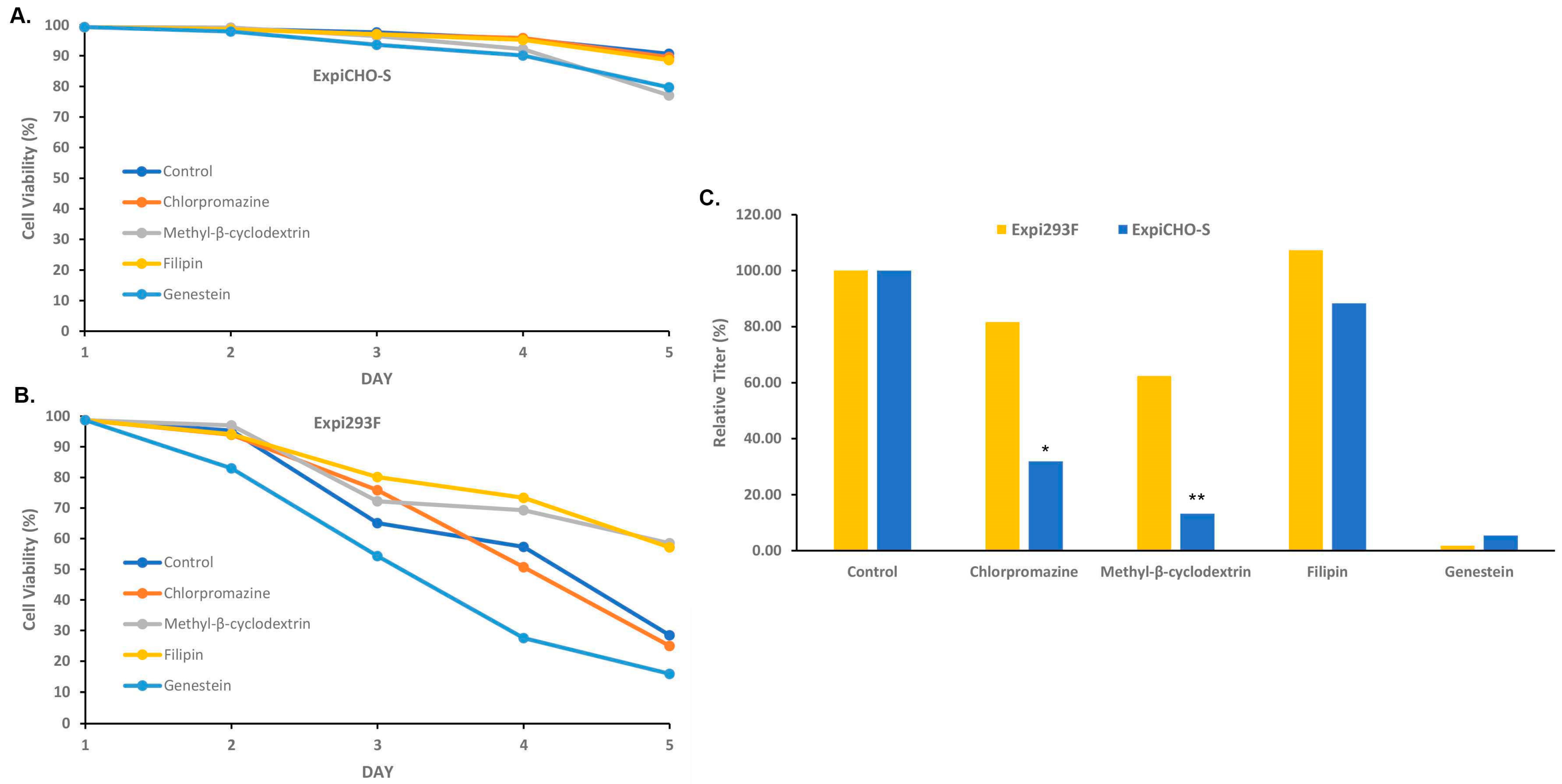
3.5. Expi293FTM and ExpiCHO-STM Cells Responded Differently to the Endocytosis Blockers
3.6. Expi293FTM and ExpiCHO-STM Cells Responded Differently to the Cotransfection of Cell Cycle Inhibitor p21 and p27
3.7. DMSO Could Enhance More Transient Expression in ExpiCHO-STM Cells Than in Expi293FTM Cells as the Concentration Increased
3.8. Expression Preferences for Kappa and Lambda Light Chain Isotype Were Detected in Expi293FTM and ExpiCHO-STM Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Urquhart, L. Top product forecasts for 2022. Nat. Rev. Drug Discov. 2022, 21, 11. [Google Scholar] [CrossRef]
- Pham, P.L.; Kamen, A.; Durocher, Y. Large-scale transfection of mammalian cells for the fast production of recombinant protein. Mol. Biotechnol. 2006, 34, 225–237. [Google Scholar] [CrossRef]
- Baldi, L.; Hacker, D.L.; Adam, M.; Wurm, F.M. Recombinant protein production by large-scale transient gene expression in mammalian cells: State of the art and future perspectives. Biotechnol. Lett. 2007, 29, 677–684. [Google Scholar] [PubMed] [Green Version]
- Geisse, S.; Voedisch, B. Transient expression technologies: Past, present, and future. Methods Mol. Biol. 2012, 899, 203–219. [Google Scholar]
- Gutiérrez-Granados, S.; Cervera, L.; Kamen, A.A.; Gòdia, F. Advancements in mammalian cell transient gene expression (TGE) technology for accelerated production of biologics. Crit. Rev. Biotechnol. 2018, 38, 918–940. [Google Scholar] [CrossRef]
- Jager, V.; Bussow, K.; Schirrmann, T. Transient Recombinant Protein Expression in Mammalian Cells. In Animal Cell Culture; Cell Engineering; Al-Rubeai, M., Ed.; Springer Interantional Publishing: Cham, Switzerland, 2015; Volume 9, pp. 27–64. [Google Scholar]
- Backliwal, G.; Hildinger, M.; Chenuet, S.; Wulhfard, S.; De Jesus, M.; Wurm, F.M. Rational vector design and multi-pathway modulation of HEK 293E cells yield recombinant antibody titers exceeding 1 g/L by transient transfection under serum-free conditions. Nucleic Acids Res. 2008, 36, e96. [Google Scholar] [CrossRef] [Green Version]
- Jain, N.K.; Barkowski-Clark, S.; Altman, R.; Johnson, K.; Sun, F.; Zmuda, J.; Liu, C.Y.; Kita, A.; Schulz, R.; Neill, A.; et al. A high density CHO-S transient transfection system: Comparison of ExpiCHO and Expi293. Protein Expr. Purif. 2017, 134, 38–46. [Google Scholar]
- Liu, C.; Dalby, B.; Chen, W.; Kilzer, J.M.; Chiou, H.C. Transient transfection factors for high-level recombinant protein production in suspension cultured mammalian cells. Mol. Biotechnol. 2008, 39, 141–153. [Google Scholar] [CrossRef]
- Ye, J.; Kober, V.; Tellers, M.; Naji, Z.; Salmon, P.; Markusen, J.F. High-level protein expression in scalable CHO transient transfection. Biotechnol. Bioeng. 2009, 103, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Daramola, O.; Stevenson, J.; Dean, G.; Hatton, D.; Pettman, G.; Holmes, W.; Field, R. A high-yielding CHO transient system: Coexpression of genes encoding EBNA-1 and GS enhances transient protein expression. Biotechnol. Prog. 2014, 30, 132–141. [Google Scholar] [PubMed]
- Rajendra, Y.; Hougland, M.D.; Alam, R.; Morehead, T.A.; Barnard, G.C. A high cell density transient transfection system for therapeutic protein expression based on a CHO GS-knockout cell line: Process development and product quality assessment. Biotechnol. Bioeng. 2015, 112, 977–986. [Google Scholar] [PubMed]
- Zhong, X.; Ma, W.; Meade, C.L.; Tam, A.S.; Llewellyn, E.; Cornell, R.; Cote, K.; Scarcelli, J.J.; Marshall, J.K.; Tzvetkova, B.; et al. Transient CHO expression platform for robust antibody production and its enhanced N-glycan sialylation on therapeutic glycoproteins. Biotechnol. Prog. 2019, 35, e2724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuible, M.; Burlacu, A.; Perret, S.; Brochu, D.; Paul-Roc, B.; Baardsnes, J.; Loignon, M.; Grazzini, E.; Durocher, Y. Optimization of a high-cell-density polyethylenimine transfection method for rapid protein production in CHO-EBNA1 cells. J. Biotechnol. 2018, 281, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, M.G.; White, R.N.; Barnard, G.C. Development of a high cell density transient CHO platform yielding mAb titers greater than 2 g/L in only 7 days. Biotechnol. Prog. 2020, 36, e3047. [Google Scholar]
- Rodriguez-Conde, S.; Inman, S.; Lindo, V.; Amery, L.; Tang, A.; Okorji-Obike, U.; Du, W.; Bosch, B.-J.; Schreur, P.J.W.; Kortekaas, J.; et al. Suitability of transiently expressed antibodies for clinical studies: Product quality consistency at different production scales. mAbs 2022, 14. [Google Scholar]
- Steger, K.; Brady, J.; Wang, W.; Duskin, M.; Donato, K.; Peshwa, M. CHO-S antibody titers >1 gram/liter using flow electroporation-mediated transient gene expression followed by rapid migration to high-yield stable cell lines. J. Biomol. Screen. 2015, 20, 545–551. [Google Scholar]
- Zhong, X.; D’Antona, A.M. Recent Advances in the Molecular Design and Applications of Multispecific Biotherapeutics. Antibodies 2021, 10, 13. [Google Scholar] [CrossRef]
- Brinkmann, U.; Kontermann, R.E. The making of bispecific antibodies. mAbs 2017, 9, 182–212. [Google Scholar] [CrossRef] [Green Version]
- Labrijn, A.F.; Janmaat, M.L.; Reichert, J.M.; Parren, P.W.H.I. Bispecific antibodies: A mechanistic review of the pipeline. Nat. Rev. Drug Discov. 2019, 18, 585–608. [Google Scholar]
- Chiou, H.C.; Vasu, S.; Liu, C.Y.; Cisneros, I.; Jones, M.B.; Zmuda, J.F. Scalable transient protein expression. Methods Mol. Biol. 2014, 1104, 35–55. [Google Scholar]
- Liu, C.Y.; Spencer, V.; Kumar, S.; Liu, J.; Chiou, H.; Zmuda, J.F. Attaining high transient titers in CHO cells: Case study involving the use of the ExpiCHOTM mammalian transient expression system. Genet. Eng. Biotechnol. News 2015, 35, 34–35. [Google Scholar] [CrossRef]
- Zhong, X.; Schenk, J.; Sakorafas, P.; Chamberland, J.; Tam, A.; Thomas, L.M.; Yan, G.; D’Antona, A.M.; Lin, L.; Nocula-Lugowska, M.; et al. Impacts of fast production of afucosylated antibodies and Fc mutants in ExpiCHO-S for enhancing FcgammaRIIIa binding and NK cell activation. J. Biotechnol. 2022, 360, 79–91. [Google Scholar] [CrossRef]
- Heng, Z.S.-L.; Yeo, J.Y.; Koh, D.W.-S.; Gan, S.K.-E.; Ling, W.-L. Augmenting recombinant antibody production in HEK293E cells: Optimizing transfection and culture parameters. Antib. Ther. 2022, 5, 30–41. [Google Scholar]
- Johari, Y.B.; Estes, S.D.; Alves, C.S.; Sinacore, M.S.; James, D.C. Integrated cell and process engineering for improved transient production of a “difficult-to-express” fusion protein by CHO cells. Biotechnol. Bioeng. 2015, 112, 2527–2542. [Google Scholar] [CrossRef] [PubMed]
- Rajendra, Y.; Hougland, M.D.; Schmitt, M.G.; Barnard, G.C. Transcriptional and post-transcriptional targeting for enhanced transient gene expression in CHO cells. Biotechnol. Lett. 2015, 37, 2379–2386. [Google Scholar] [CrossRef] [PubMed]
- Kiszel, P.; Fiesel, S.; Voit, S.; Waechtler, B.; Meier, T.; Oelschlaegel, T.; Schraeml, M.; Engel, A.M. Transient gene expression using valproic acid in combination with co-transfection of SV40 large T antigen and human p21(CIP)/p27(KIP). Biotechnol. Prog. 2019, 35, e2786. [Google Scholar] [CrossRef] [PubMed]
- Pham, P.L.; Perret, S.; Cass, B.; Carpentier, E.; St-Laurent, G.; Bisson, L.; Kamen, A.; Durocher, Y. Transient gene expression in HEK293 cells: Peptone addition posttransfection improves recombinant protein synthesis. Biotechnol. Bioeng. 2005, 90, 332–344. [Google Scholar]
- Zhong, X.; Schwab, A.; Ma, W.; Meade, C.L.; Zhou, J.; D’antona, A.M.; Somers, W.; Lin, L. Large-Scale Transient Production in ExpiCHO-S with Enhanced N-Galactosylation-Sialylation and PEI-Based Transfection. Methods Mol. Biol. 2022, 2313, 143–150. [Google Scholar]
- Nomani, A.; Hyvönen, Z.; Pulkkinen, E.; Hiekkala, M.; Ruponen, M. Intracellular gene delivery is dependent on the type of non-viral carrier and defined by the cell surface glycosaminoglycans. J. Control Release 2014, 187, 59–65. [Google Scholar] [CrossRef]
- Wang, L.H.; Rothberg, K.G.; Anderson, R.G. Mis-assembly of clathrin lattices on endosomes reveals a regulatory switch for coated pit formation. J. Cell Biol. 1993, 123, 1107–1117. [Google Scholar] [CrossRef]
- Lamaze, C.; Schmid, S.L. The emergence of clathrin-independent pinocytic pathways. Curr. Opin. Cell Biol. 1995, 7, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Parton, R.G.; Richards, A.A. Lipid rafts and caveolae as portals for endocytosis: New insights and common mechanisms. Traffic 2003, 4, 724–738. [Google Scholar] [PubMed] [Green Version]
- Khalil, I.A.; Kogure, K.; Akita, H.; Harashima, H. Uptake pathways and subsequent intracellular trafficking in nonviral gene delivery. Pharmacol. Rev. 2006, 58, 32–45. [Google Scholar] [CrossRef] [Green Version]
- Tiruppathi, C.; Song, W.; Bergenfeldt, M.; Sass, P.; Malik, A.B. Gp60 activation mediates albumin transcytosis in endothelial cells by tyrosine kinase-dependent pathway. J. Biol. Chem. 1997, 272, 25968–25975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coqueret, O. New roles for p21 and p27 cell-cycle inhibitors: A function for each cell compartment? Trends Cell Biol. 2003, 13, 65–70. [Google Scholar] [PubMed]
- Vink, T.; Oudshoorn-Dickmann, M.; Roza, M.; Reitsma, J.J.; de Jong, R.N. A simple, robust and highly efficient transient expression system for producing antibodies. Methods 2014, 65, 5–10. [Google Scholar] [CrossRef]
- Toogood, P.L.; Harvey, P.J.; Repine, J.T.; Sheehan, D.J.; VanderWel, S.N.; Zhou, H.; Keller, P.R.; McNamara, D.J.; Sherry, D.; Zhu, T.; et al. Discovery of a potent and selective inhibitor of cyclin-dependent kinase 4/6. J. Med. Chem. 2005, 48, 2388–2406. [Google Scholar] [CrossRef]
- Rajendra, Y.; Balasubramanian, S.; Kiseljak, D.; Baldi, L.; Wurm, F.M.; Hacker, D.L. Enhanced plasmid DNA utilization in transiently transfected CHO-DG44 cells in the presence of polar solvents. Biotechnol. Prog. 2015, 31, 1571–1578. [Google Scholar] [CrossRef]
- Molé, C.M.; Béné, M.C.; Montagne, P.M.; Seilles, E.; Faure, G.C. Light chains of immunoglobulins in human secretions. Clin. Chim. Acta 1994, 224, 191–197. [Google Scholar] [CrossRef]
- Popov, A.V.; Zou, X.; Xian, J.; Nicholson, I.C.; Brüggemann, M. A human immunoglobulin lambda locus is similarly well expressed in mice and humans. J. Exp. Med. 1999, 189, 1611–1620. [Google Scholar] [CrossRef]
- Arun, S.S.; Breuer, W.; Hermanns, W. Immunohistochemical examination of light-chain expression (lambda/kappa ratio) in canine, feline, equine, bovine and porcine plasma cells. Zentralbl Vet. A 1996, 43, 573–576. [Google Scholar]
- Conner, S.D.; Schmid, S.L. Regulated portals of entry into the cell. Nature 2003, 422, 37–44. [Google Scholar] [CrossRef]
- de Ménorval, M.A.; Mir, L.M.; Fernández, M.L.; Reigada, R. Effects of dimethyl sulfoxide in cholesterol-containing lipid membranes: A comparative study of experiments in silico and with cells. PLoS ONE 2012, 7, e41733. [Google Scholar]
- Tunçer, S.; Gurbanov, R.; Sheraj, I.; Solel, E.; Esenturk, O.; Banerjee, S. Low dose dimethyl sulfoxide driven gross molecular changes have the potential to interfere with various cellular processes. Sci. Rep. 2018, 8, 14828. [Google Scholar]
- Liu, C.H.; Chen, L.H. Promotion of recombinant macrophage colony stimulating factor production by dimethyl sulfoxide addition in Chinese hamster ovary cells. J. Biosci. Bioeng. 2007, 103, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.J.; Jeon, C.J.; Cho, S.M.; Lee, G.M.; Yoon, S.K. Effect of chemical chaperone addition on production and aggregation of recombinant flag-tagged COMP-angiopoietin 1 in Chinese hamster ovary cells. Biotechnol. Prog. 2011, 27, 587–591. [Google Scholar] [CrossRef]
- Lim, S.; Kaldis, P. Cdks, cyclins and CKIs: Roles beyond cell cycle regulation. Development 2013, 140, 3079–3093. [Google Scholar] [PubMed] [Green Version]
- Fox, S.R.; Patel, U.A.; Yap, M.G.; Wang, D.I. Maximizing interferon-gamma production by Chinese hamster ovary cells through temperature shift optimization: Experimental and modeling. Biotechnol. Bioeng. 2004, 85, 177–184. [Google Scholar] [PubMed]
- Kaufmann, H.; Mazur, X.; Fussenegger, M.; Bailey, J.E. Influence of low temperature on productivity, proteome and protein phosphorylation of CHO cells. Biotechnol. Bioeng. 1999, 63, 573–582. [Google Scholar] [CrossRef]
- Hao, C.; Liu, P.; Zhao, L.; Zhu, C.; Tian, X.; Wang, L.; Zhang, Z. Inhibition of leukemic cells by valproic acid, an HDAC inhibitor, in xenograft tumors. Onco Targets Ther. 2013, 6, 733–740. [Google Scholar] [CrossRef] [Green Version]
- Meyer, H.J.; Turincio, R.; Ng, S.; Li, J.; Wilson, B.; Chan, P.; Zak, M.; Reilly, D.; Beresini, M.H.; Wong, A.W. High throughput screening identifies novel, cell cycle-arresting small molecule enhancers of transient protein expression. Biotechnol. Prog. 2017, 33, 1579–1588. [Google Scholar] [CrossRef]
- Du, Z.; Treiber, D.; McCarter, J.D.; Fomina-Yadlin, D.; Saleem, R.A.; McCoy, R.E.; Zhang, Y.; Tharmalingam, T.; Leith, M.; Follstad, B.D.; et al. Use of a small molecule cell cycle inhibitor to control cell growth and improve specific productivity and product quality of recombinant proteins in CHO cell cultures. Biotechnol. Bioeng. 2015, 112, 141–155. [Google Scholar] [CrossRef]
- Moghimi, S.M.; Symonds, P.; Murray, J.C.; Hunter, A.; Debska, G.; Szewczyk, A.C. A two-stage poly(ethylenimine)-mediated cytotoxicity: Implications for gene transfer/therapy. Mol. Ther. 2005, 11, 990–995. [Google Scholar]
- Schlatter, S.; Stansfield, S.H.; Dinnis, D.M.; Racher, A.J.; Birch, J.R.; James, D.C. On the optimal ratio of heavy to light chain genes for efficient recombinant antibody production by CHO cells. Biotechnol. Prog. 2005, 21, 122–133. [Google Scholar]
- Ling, W.L.; Lua, W.H.; Poh, J.J.; Yeo, J.Y.; Lane, D.P.; Gan, S.K.E. Effect of VH-VL Families in Pertuzumab and Trastuzumab Recombinant Production, Her2 and FcgammaIIA Binding. Front. Immunol. 2018, 9, 469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lua, W.H.; Ling, W.L.; Yeo, J.Y.; Poh, J.J.; Lane, D.P.; Gan, S.K.E. The effects of Antibody Engineering CH and CL in Trastuzumab and Pertuzumab recombinant models: Impact on antibody production and antigen-binding. Sci. Rep. 2018, 8, 718. [Google Scholar] [PubMed] [Green Version]
- Gong, S.; Gautam, S.; Coneglio, J.D.; Scinto, H.B.; Ruprecht, R.M. Antibody Light Chains: Key to Increased Monoclonal Antibody Yields in Expi293 Cells? Antibodies 2022, 11, 37. [Google Scholar]
- Malm, M.; Kuo, C.-C.; Barzadd, M.M.; Mebrahtu, A.; Wistbacka, N.; Razavi, R.; Volk, A.-L.; Lundqvist, M.; Kotol, D.; Tegel, H.; et al. Harnessing secretory pathway differences between HEK293 and CHO to rescue production of difficult to express proteins. Metab. Eng. 2022, 72, 171–187. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Expi293FTM | ExpiCHO-STM |
|---|---|---|
| Viable Cell Density | 3.0 × 106/mL | 6.0 × 106/mL |
| Complexation Time (ExpiFectamineTM) | 5 min | 2.5 min |
| Complexation Time (PEI) | 15 min | 2.5 min |
| Opti-MEMTM volume (ExpiFectamineTM/PEI) | 100 mL/L | 100 mL/L |
| 1% DMDO (ExpiFectamineTM/PEI) | - | + |
| P27/p21 1:1 | - | + |
| Preferred Light Chain | Lambda/Kappa | Kappa |
| PBS as dilution buffer | + | - |
| DNA:PEI ratio | 1:8 | 1:3.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.; Yan, G.G.; Cluckey, D.; Meade, C.; Ruth, M.; Sorm, R.; Tam, A.S.; Lim, S.; Petridis, C.; Lin, L.; et al. Exploring Parametric and Mechanistic Differences between Expi293FTM and ExpiCHO-STM Cells for Transient Antibody Production Optimization. Antibodies 2023, 12, 53. https://doi.org/10.3390/antib12030053
Zhou J, Yan GG, Cluckey D, Meade C, Ruth M, Sorm R, Tam AS, Lim S, Petridis C, Lin L, et al. Exploring Parametric and Mechanistic Differences between Expi293FTM and ExpiCHO-STM Cells for Transient Antibody Production Optimization. Antibodies. 2023; 12(3):53. https://doi.org/10.3390/antib12030053
Chicago/Turabian StyleZhou, Jing, Guoying Grace Yan, David Cluckey, Caryl Meade, Margaret Ruth, Rhady Sorm, Amy S. Tam, Sean Lim, Constantine Petridis, Laura Lin, and et al. 2023. "Exploring Parametric and Mechanistic Differences between Expi293FTM and ExpiCHO-STM Cells for Transient Antibody Production Optimization" Antibodies 12, no. 3: 53. https://doi.org/10.3390/antib12030053
APA StyleZhou, J., Yan, G. G., Cluckey, D., Meade, C., Ruth, M., Sorm, R., Tam, A. S., Lim, S., Petridis, C., Lin, L., D’Antona, A. M., & Zhong, X. (2023). Exploring Parametric and Mechanistic Differences between Expi293FTM and ExpiCHO-STM Cells for Transient Antibody Production Optimization. Antibodies, 12(3), 53. https://doi.org/10.3390/antib12030053