
An Engineered Mouse Model That Generates a Diverse Repertoire of Endogenous, High-Affinity Common Light Chain Antibodies


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mouse Model Generation
2.2. Serum Concentration of IgG, IgM and Kappa Antibodies by ELISA
2.3. Animal Immunization
2.4. B Cell Phenotypic Analysis by Flow Cytometry
2.5. Antigen-Specific B Cell Sorting
2.6. Single-Cell Sequencing and Analysis
2.7. Heavy Chain Immune Repertoire Sequencing and Analysis
2.8. Small-Scale Antibody Expression
2.9. Enzyme-Linked Immunosorbent Assay (ELISA) to Determine Antigen Binding
2.10. Biacore
2.11. Epitope Binning of Wildtype and cLC Antibodies
2.12. Statistical Analysis
3. Results
3.1. Model Generation
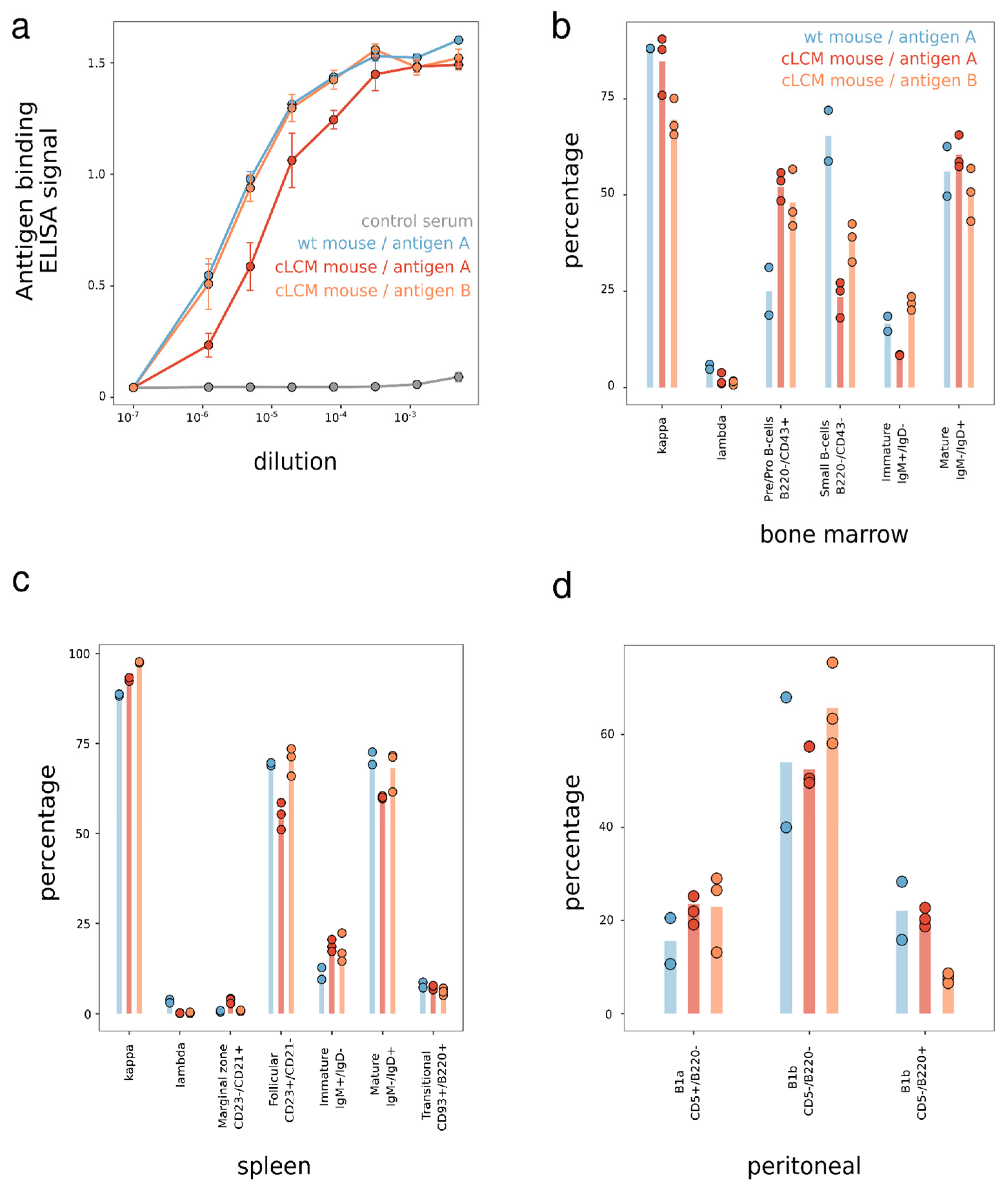
3.2. B Cell Development in the Common Light Chain Mice
3.3. Immune Repertoire—Isotype and V Gene Usage
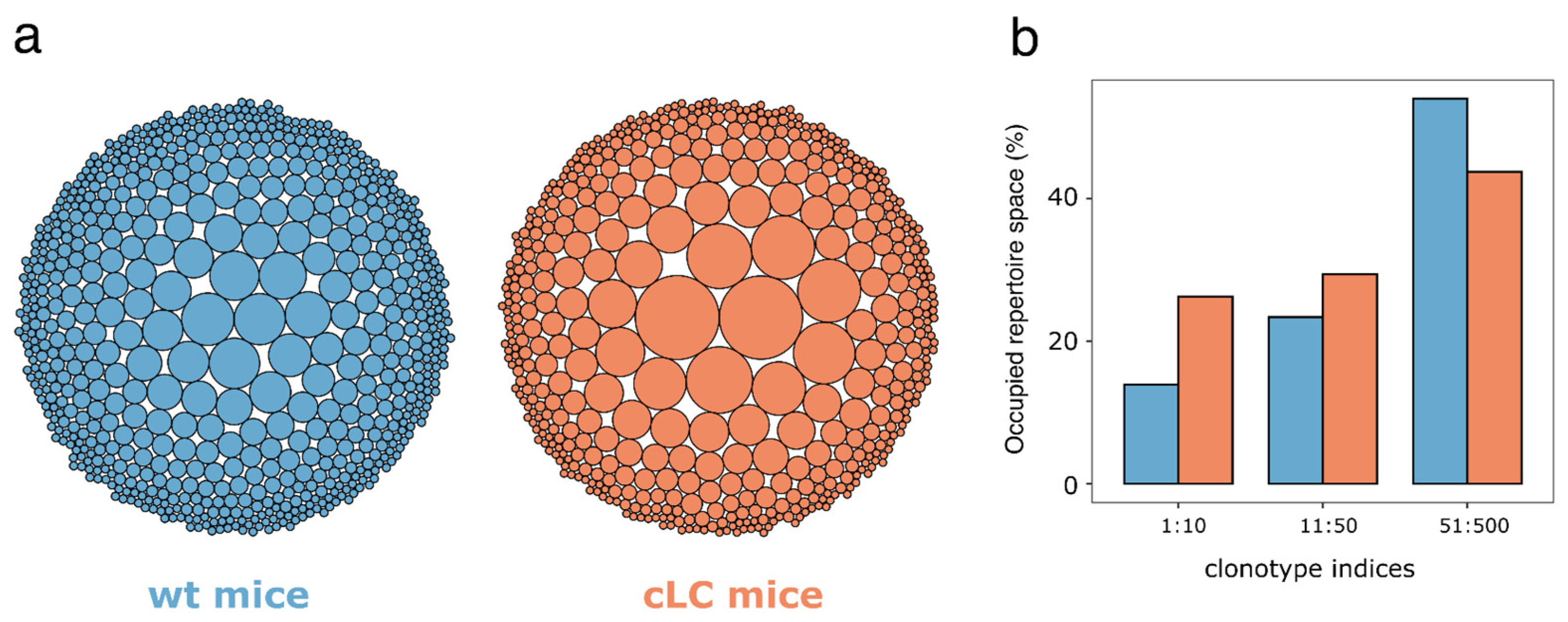
3.4. Immune Repertoire—Clonotype Diversity
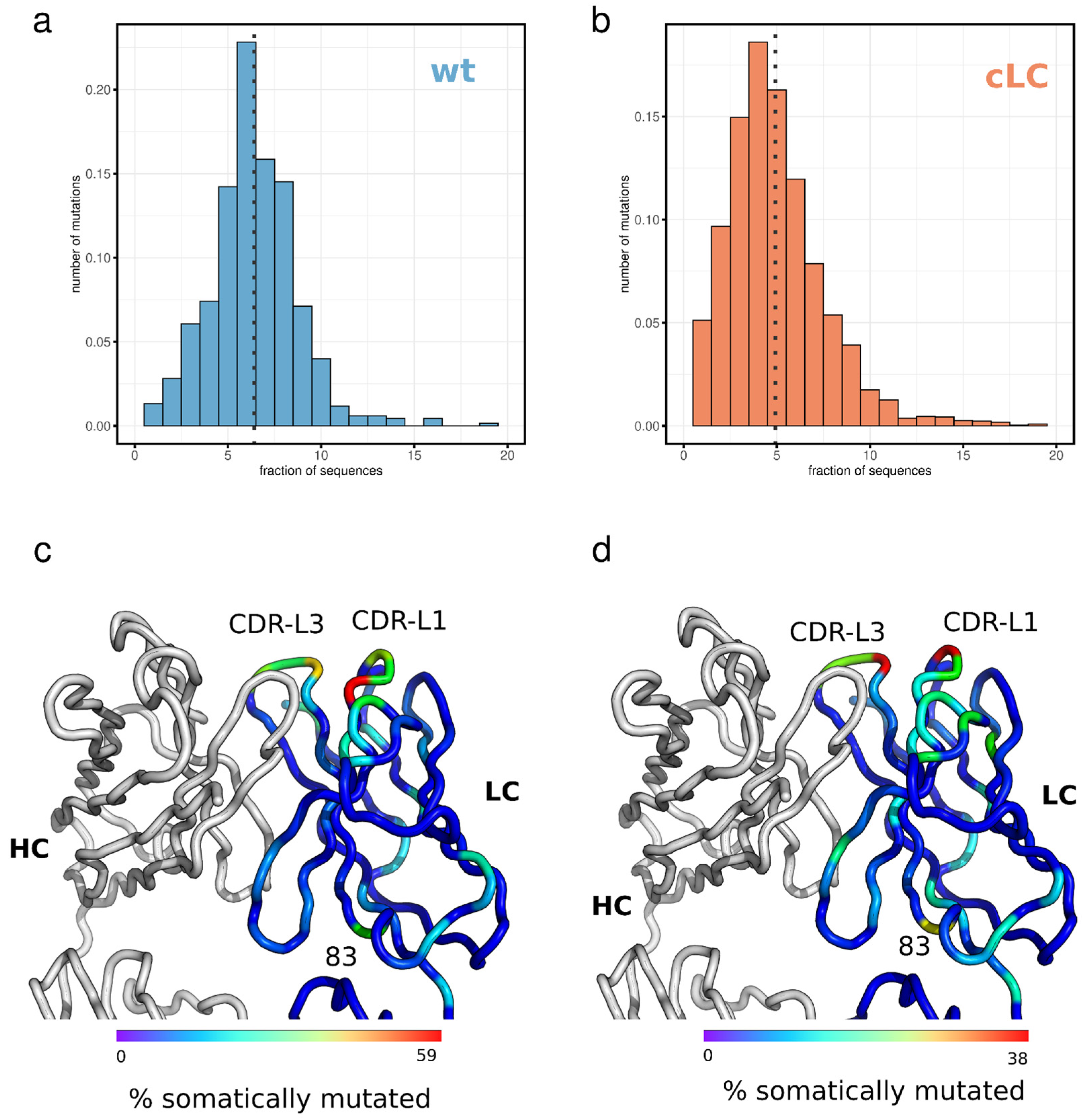
3.5. Light Chain Mutation Profile
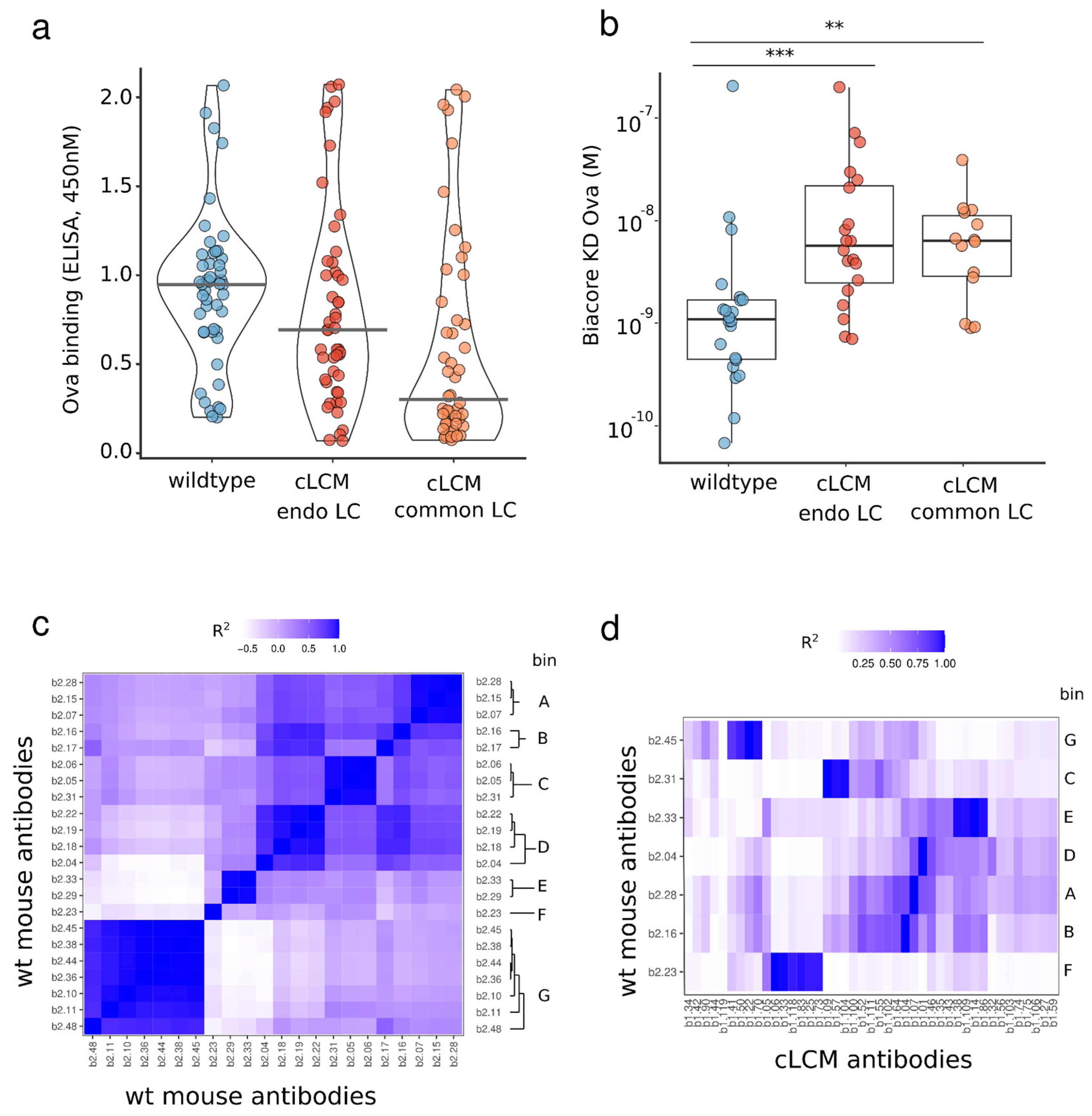
3.6. Identification of OVA-Specific Antibodies
3.7. Epitope Binning of Wildtype and cLC Antibodies
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carter, P.J.; Lazar, G.A. Next Generation Antibody Drugs: Pursuit of the ‘High-Hanging Fruit’. Nat. Rev. Drug Discov. 2018, 17, 197–223. [Google Scholar] [CrossRef]
- Spiess, C.; Zhai, Q.; Carter, P.J. Alternative Molecular Formats and Therapeutic Applications for Bispecific Antibodies. Mol. Immunol. 2015, 67, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Mo, Y.; Tang, M.; Shen, J.; Qi, Y.; Zhao, W.; Huang, Y.; Xu, Y.; Qian, C. Bispecific Antibodies: From Research to Clinical Application. Front. Immunol. 2021, 12, 626616. [Google Scholar] [CrossRef] [PubMed]
- Hust, M.; Jostock, T.; Menzel, C.; Voedisch, B.; Mohr, A.; Brenneis, M.; Kirsch, M.I.; Meier, D.; Dübel, S. Single Chain Fab (scFab) Fragment. BMC Biotechnol. 2007, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, W.; Regula, J.T.; Bähner, M.; Schanzer, J.; Croasdale, R.; Dürr, H.; Gassner, C.; Georges, G.; Kettenberger, H.; Imhof-Jung, S.; et al. Immunoglobulin Domain Crossover as a Generic Approach for the Production of Bispecific IgG Antibodies. Proc. Natl. Acad. Sci. USA 2011, 108, 11187–11192. [Google Scholar] [CrossRef] [PubMed]
- Fischer, N.; Elson, G.; Magistrelli, G.; Dheilly, E.; Fouque, N.; Laurendon, A.; Gueneau, F.; Ravn, U.; Depoisier, J.-F.; Moine, V.; et al. Exploiting Light Chains for the Scalable Generation and Platform Purification of Native Human Bispecific IgG. Nat. Commun. 2015, 6, 6113. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.; Han, J.; Wang, Y.; Li, Y. A Potential Downstream Platform Approach for WuXiBody-Based IgG-like Bispecific Antibodies. Protein Expr. Purif. 2020, 173, 105647. [Google Scholar] [CrossRef] [PubMed]
- Spiess, C.; Merchant, M.; Huang, A.; Zheng, Z.; Yang, N.-Y.; Peng, J.; Ellerman, D.; Shatz, W.; Reilly, D.; Yansura, D.G.; et al. Bispecific Antibodies with Natural Architecture Produced by Co-Culture of Bacteria Expressing Two Distinct Half-Antibodies. Nat. Biotechnol. 2013, 31, 753–758. [Google Scholar] [CrossRef]
- Labrijn, A.F.; Meesters, J.I.; de Goeij, B.E.C.G.; van den Bremer, E.T.J.; Neijssen, J.; van Kampen, M.D.; Strumane, K.; Verploegen, S.; Kundu, A.; Gramer, M.J.; et al. Efficient Generation of Stable Bispecific IgG1 by Controlled Fab-Arm Exchange. Proc. Natl. Acad. Sci. USA 2013, 110, 5145–5150. [Google Scholar] [CrossRef]
- Steinhardt, J.; Wu, Y.; Fleming, R.; Ruddle, B.T.; Patel, P.; Wu, H.; Gao, C.; Dimasi, N. Fab-Arm Exchange Combined with Selective Protein A Purification Results in a Platform for Rapid Preparation of Monovalent Bispecific Antibodies Directly from Culture Media. Pharmaceutics 2020, 12, 3. [Google Scholar] [CrossRef]
- Ridgway, J.B.B.; Presta, L.G.; Carter, P. ‘Knobs-into-Holes’ Engineering of Antibody CH3 Domains for Heavy Chain Heterodimerization. Protein Eng. Des. Sel. 1996, 9, 617–621. [Google Scholar] [CrossRef] [PubMed]
- Shiraiwa, H.; Narita, A.; Kamata-Sakurai, M.; Ishiguro, T.; Sano, Y.; Hironiwa, N.; Tsushima, T.; Segawa, H.; Tsunenari, T.; Ikeda, Y.; et al. Engineering a Bispecific Antibody with a Common Light Chain: Identification and Optimization of an Anti-CD3 Epsilon and Anti-GPC3 Bispecific Antibody, ERY974. Methods 2019, 154, 10–20. [Google Scholar] [CrossRef] [PubMed]
- De Nardis, C.; Hendriks, L.J.A.; Poirier, E.; Arvinte, T.; Gros, P.; Bakker, A.B.H.; de Kruif, J. A New Approach for Generating Bispecific Antibodies Based on a Common Light Chain Format and the Stable Architecture of Human Immunoglobulin G1. J. Biol. Chem. 2017, 292, 14706–14717. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.H.; Aperlo, C.; Li, Y.; Kurosawa, E.; Lan, Y.; Lo, K.-M.; Huston, J.S. SEEDbodies: Fusion Proteins Based on Strand-Exchange Engineered Domain (SEED) CH3 Heterodimers in an Fc Analogue Platform for Asymmetric Binders or Immunofusions and Bispecific Antibodies. Protein Eng. Des. Sel. 2010, 23, 195–202. [Google Scholar] [CrossRef]
- Gunasekaran, K.; Pentony, M.; Shen, M.; Garrett, L.; Forte, C.; Woodward, A.; Ng, S.B.; Born, T.; Retter, M.; Manchulenko, K.; et al. Enhancing Antibody Fc Heterodimer Formation through Electrostatic Steering Effects: Applications to Bispecific Molecules and Monovalent IgG. J. Biol. Chem. 2010, 285, 19637–19646. [Google Scholar] [CrossRef]
- Strop, P.; Ho, W.-H.; Boustany, L.M.; Abdiche, Y.N.; Lindquist, K.C.; Farias, S.E.; Rickert, M.; Appah, C.T.; Pascua, E.; Radcliffe, T.; et al. Generating Bispecific Human IgG1 and IgG2 Antibodies from Any Antibody Pair. J. Mol. Biol. 2012, 420, 204–219. [Google Scholar] [CrossRef]
- Lewis, S.M.; Wu, X.; Pustilnik, A.; Sereno, A.; Huang, F.; Rick, H.L.; Guntas, G.; Leaver-Fay, A.; Smith, E.M.; Ho, C.; et al. Generation of Bispecific IgG Antibodies by Structure-Based Design of an Orthogonal Fab Interface. Nat. Biotechnol. 2014, 32, 191–198. [Google Scholar] [CrossRef]
- Dillon, M.; Yin, Y.; Zhou, J.; McCarty, L.; Ellerman, D.; Slaga, D.; Junttila, T.T.; Han, G.; Sandoval, W.; Ovacik, M.A.; et al. Efficient Production of Bispecific IgG of Different Isotypes and Species of Origin in Single Mammalian Cells. MAbs 2016, 9, 213–230. [Google Scholar] [CrossRef]
- Liu, Z.; Leng, E.C.; Gunasekaran, K.; Pentony, M.; Shen, M.; Howard, M.; Stoops, J.; Manchulenko, K.; Razinkov, V.; Liu, H.; et al. A Novel Antibody Engineering Strategy for Making Monovalent Bispecific Heterodimeric IgG Antibodies by Electrostatic Steering Mechanism. J. Biol. Chem. 2015, 290, 7535–7562. [Google Scholar] [CrossRef]
- Mazor, Y.; Oganesyan, V.; Yang, C.; Hansen, A.; Wang, J.; Liu, H.; Sachsenmeier, K.; Carlson, M.; Gadre, D.V.; Borrok, M.J.; et al. Improving Target Cell Specificity Using a Novel Monovalent Bispecific IgG Design. MAbs 2015, 7, 377–389. [Google Scholar] [CrossRef]
- Sampei, Z.; Igawa, T.; Soeda, T.; Okuyama-Nishida, Y.; Moriyama, C.; Wakabayashi, T.; Tanaka, E.; Muto, A.; Kojima, T.; Kitazawa, T.; et al. Identification and Multidimensional Optimization of an Asymmetric Bispecific IgG Antibody Mimicking the Function of Factor VIII Cofactor Activity. PLoS ONE 2013, 8, e57479. [Google Scholar] [CrossRef]
- Kitazawa, T.; Igawa, T.; Sampei, Z.; Muto, A.; Kojima, T.; Soeda, T.; Yoshihashi, K.; Okuyama-Nishida, Y.; Saito, H.; Tsunoda, H.; et al. A Bispecific Antibody to Factors IXa and X Restores Factor VIII Hemostatic Activity in a Hemophilia A Model. Nat. Med. 2012, 18, 1570–1574. [Google Scholar] [CrossRef]
- Geuijen, C.A.W.; De Nardis, C.; Maussang, D.; Rovers, E.; Gallenne, T.; Hendriks, L.J.A.; Visser, T.; Nijhuis, R.; Logtenberg, T.; de Kruif, J.; et al. Unbiased Combinatorial Screening Identifies a Bispecific IgG1 That Potently Inhibits HER3 Signaling via HER2-Guided Ligand Blockade. Cancer Cell 2018, 33, 922–936.e10. [Google Scholar] [CrossRef]
- Smith, E.J.; Olson, K.; Haber, L.J.; Varghese, B.; Duramad, P.; Tustian, A.D.; Oyejide, A.; Kirshner, J.R.; Canova, L.; Menon, J.; et al. A Novel, Native-Format Bispecific Antibody Triggering T-Cell Killing of B-Cells Is Robustly Active in Mouse Tumor Models and Cynomolgus Monkeys. Sci. Rep. 2015, 5, 17943. [Google Scholar] [CrossRef]
- DiLillo, D.J.; Olson, K.; Mohrs, K.; Meagher, T.C.; Bray, K.; Sineshchekova, O.; Startz, T.; Kuhnert, J.; Retter, M.W.; Godin, S.; et al. A BCMAxCD3 Bispecific T Cell–Engaging Antibody Demonstrates Robust Antitumor Efficacy Similar to That of Anti-BCMA CAR T Cells. Blood Adv. 2021, 5, 1291–1304. [Google Scholar] [CrossRef]
- Merchant, A.M.; Zhu, Z.; Yuan, J.Q.; Goddard, A.; Adams, C.W.; Presta, L.G.; Carter, P. An Efficient Route to Human Bispecific IgG. Nat. Biotechnol. 1998, 16, 677–681. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.V.; Liang, W.-C.; Dennis, M.S.; Eigenbrot, C.; Sidhu, S.S.; Fuh, G. High-Affinity Human Antibodies from Phage-Displayed Synthetic Fab Libraries with a Single Framework Scaffold. J. Mol. Biol. 2004, 340, 1073–1093. [Google Scholar] [CrossRef] [PubMed]
- Van Blarcom, T.; Rossi, A.; Foletti, D.; Sundar, P.; Pitts, S.; Bee, C.; Melton Witt, J.; Melton, Z.; Hasa-Moreno, A.; Shaughnessy, L.; et al. Precise and Efficient Antibody Epitope Determination through Library Design, Yeast Display and Next-Generation Sequencing. J. Mol. Biol. 2015, 427, 1513–1534. [Google Scholar] [CrossRef] [PubMed]
- Krah, S.; Schröter, C.; Eller, C.; Rhiel, L.; Rasche, N.; Beck, J.; Sellmann, C.; Günther, R.; Toleikis, L.; Hock, B.; et al. Generation of Human Bispecific Common Light Chain Antibodies by Combining Animal Immunization and Yeast Display. Protein Eng. Des. Sel. 2017, 30, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Mcwhirter, J.; Macdonald, L.; Stevens, S.; Davis, S.; Murphy, A.J.; Buckler, D.R. Common Light Chain Mouse. European Patent EP2505654B2, 13 May 2020. [Google Scholar]
- Harris, K.E.; Aldred, S.F.; Davison, L.M.; Ogana, H.A.N.; Boudreau, A.; Brüggemann, M.; Osborn, M.; Ma, B.; Buelow, B.; Clarke, S.C.; et al. Sequence-Based Discovery Demonstrates That Fixed Light Chain Human Transgenic Rats Produce a Diverse Repertoire of Antigen-Specific Antibodies. Front. Immunol. 2018, 9, 889. [Google Scholar] [CrossRef] [PubMed]
- Ching, K.H.; Berg, K.; Reynolds, K.; Pedersen, D.; Macias, A.; Abdiche, Y.N.; Harriman, W.D.; Leighton, P.A. Common Light Chain Chickens Produce Human Antibodies of High Affinity and Broad Epitope Coverage for the Engineering of Bispecifics. MAbs 2021, 13, 1862451. [Google Scholar] [CrossRef]
- Hui, L.; Lu, H.; Zhang, Y.; Huang, S.; Zhang, P.; Zhao, H.; Chang, R.; Yang, B.; Zhang, F.; Yang, W.; et al. Abstract 1861: Advancing Bispecific Antibody Discovery Using Common Light Chain Immunoglobulin Humanized Mouse. Cancer Res. 2021, 81, 1861. [Google Scholar] [CrossRef]
- Logtenberg, T.; Throsby, M.; Kramer, R.A.; Pinto, R.D.; de Kruif, C.A.; Houtzager, E. Antibody Producing Non-Human Animals. U.S. Patent US9944695B2, 17 April 2018. [Google Scholar]
- Pelanda, R.; Schwers, S.; Sonoda, E.; Torres, R.M.; Nemazee, D.; Rajewsky, K. Receptor Editing in a Transgenic Mouse Model: Site, Efficiency, and Role in B Cell Tolerance and Antibody Diversification. Immunity 1997, 7, 765–775. [Google Scholar] [CrossRef]
- Prak, E.L.; Weigert, M. Light Chain Replacement: A New Model for Antibody Gene Rearrangement. J. Exp. Med. 1995, 182, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Fraenkel, S.; Mostoslavsky, R.; Novobrantseva, T.I.; Pelanda, R.; Chaudhuri, J.; Esposito, G.; Jung, S.; Alt, F.W.; Rajewsky, K.; Cedar, H.; et al. Allelic ‘choice’ Governs Somatic Hypermutation in Vivo at the Immunoglobulin κ-Chain Locus. Nat. Immunol. 2007, 8, 715–722. [Google Scholar] [CrossRef]
- Luchner, M.; Reinke, S.; Milicic, A. TLR Agonists as Vaccine Adjuvants Targeting Cancer and Infectious Diseases. Pharmaceutics 2021, 13, 142. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for Clustering the next-Generation Sequencing Data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Turchaninova, M.A.; Davydov, A.; Britanova, O.V.; Shugay, M.; Bikos, V.; Egorov, E.S.; Kirgizova, V.I.; Merzlyak, E.M.; Staroverov, D.B.; Bolotin, D.A.; et al. High-Quality Full-Length Immunoglobulin Profiling with Unique Molecular Barcoding. Nat. Protoc. 2016, 11, 1599–1616. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Pharmaceutics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Ma, N.; Madden, T.L.; Ostell, J.M. IgBLAST: An Immunoglobulin Variable Domain Sequence Analysis Tool. Nucleic Acids Res. 2013, 41, W34–W40. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer: New York, NY, USA, 2016. [Google Scholar]
- Chan, B.M.; Badh, A.; Berry, K.A.; Grauer, S.A.; King, C.T. Flow Cytometry-Based Epitope Binning Using Competitive Binding Profiles for the Characterization of Monoclonal Antibodies against Cellular and Soluble Protein Targets. SLAS Discov. Adv. Sci. Drug Discov. 2018, 23, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Kearney, J.F.; Grusby, M.J.; Benoist, C.; Mathis, D. Induction of Tolerance in Arthritogenic B Cells with Receptors of Differing Affinity for Self-Antigen. Proc. Natl. Acad. Sci. USA 2006, 103, 3734–3739. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, T.; Wabl, M. Autoreactivity and Allelic Inclusion in a B Cell Nuclear Transfer Mouse. Nat. Immunol. 2004, 5, 1282–1287. [Google Scholar] [CrossRef] [PubMed]
- Aoki-Ota, M.; Torkamani, A.; Ota, T.; Schork, N.; Nemazee, D. Skewed Primary Igκ Repertoire and V–J Joining in C57BL/6 Mice: Implications for Recombination Accessibility and Receptor Editing. J. Immunol. 2012, 188, 2305–2315. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, L.D.; Chen, Y.-J.J.; Wu, J.; Chaudhuri, S.; Hsiao, Y.-C.; Schneider, K.; Hoi, K.H.; Lin, Z.; Guerrero, S.; Jaiswal, B.S.; et al. Massively Parallel Single-Cell B-Cell Receptor Sequencing Enables Rapid Discovery of Diverse Antigen-Reactive Antibodies. Commun. Biol. 2019, 2, 304. [Google Scholar] [CrossRef]
- Rettig, T.A.; Ward, C.; Bye, B.A.; Pecaut, M.J.; Chapes, S.K. Characterization of the Naive Murine Antibody Repertoire Using Unamplified High-Throughput Sequencing. PLoS ONE 2018, 13, e0190982. [Google Scholar] [CrossRef]
- Raybould, M.I.J.; Rees, A.R.; Deane, C.M. Current Strategies for Detecting Functional Convergence across B-Cell Receptor Repertoires. MAbs 2021, 13, 1996732. [Google Scholar] [CrossRef]
- Ji, J.A.; Zhang, B.; Cheng, W.; Wang, Y.J. Methionine, Tryptophan, and Histidine Oxidation in a Model Protein, PTH: Mechanisms and Stabilization. J. Pharm. Sci. 2009, 98, 4485–4500. [Google Scholar] [CrossRef]
- Foote, J.; Winter, G. Antibody Framework Residues Affecting the Conformation of the Hypervariable Loops. J. Mol. Biol. 1992, 224, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Prihoda, D.; Maamary, J.; Waight, A.; Juan, V.; Fayadat-Dilman, L.; Svozil, D.; Bitton, D.A. BioPhi: A Platform for Antibody Design, Humanization, and Humanness Evaluation Based on Natural Antibody Repertoires and Deep Learning. MAbs 2022, 14, 2020203. [Google Scholar] [CrossRef] [PubMed]
- Koenig, P.; Lee, C.V.; Walters, B.T.; Janakiraman, V.; Stinson, J.; Patapoff, T.W.; Fuh, G. Mutational Landscape of Antibody Variable Domains Reveals a Switch Modulating the Interdomain Conformational Dynamics and Antigen Binding. Proc. Natl. Acad. Sci. USA 2017, 114, E486–E495. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.M.; Lawrence, K.R.; Moulana, A.; Dupic, T.; Chang, J.; Johnson, M.S.; Cvijovic, I.; Mora, T.; Walczak, A.M.; Desai, M.M. Binding Affinity Landscapes Constrain the Evolution of Broadly Neutralizing Anti-Influenza Antibodies. eLife 2021, 10, e71393. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Ju, B.; Shapero, B.; Lin, X.; Ren, L.; Zhang, L.; Li, D.; Zhou, Z.; Feng, Y.; Sou, C.; et al. A VH1-69 Antibody Lineage from an Infected Chinese Donor Potently Neutralizes HIV-1 by Targeting the V3 Glycan Supersite. Sci. Adv. 2020, 6, eabb1328. [Google Scholar] [CrossRef]
- Zemlin, M.; Klinger, M.; Link, J.; Zemlin, C.; Bauer, K.; Engler, J.A.; Schroeder, H.W.; Kirkham, P.M. Expressed Murine and Human CDR-H3 Intervals of Equal Length Exhibit Distinct Repertoires That Differ in Their Amino Acid Composition and Predicted Range of Structures. J. Mol. Biol. 2003, 334, 733–749. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rong, Y.; Chen, I.-L.; Larrabee, L.; Sawant, M.S.; Fuh, G.; Koenig, P. An Engineered Mouse Model That Generates a Diverse Repertoire of Endogenous, High-Affinity Common Light Chain Antibodies. Antibodies 2024, 13, 14. https://doi.org/10.3390/antib13010014
Rong Y, Chen I-L, Larrabee L, Sawant MS, Fuh G, Koenig P. An Engineered Mouse Model That Generates a Diverse Repertoire of Endogenous, High-Affinity Common Light Chain Antibodies. Antibodies. 2024; 13(1):14. https://doi.org/10.3390/antib13010014
Chicago/Turabian StyleRong, Yinghui, I-Ling Chen, Lance Larrabee, Manali S. Sawant, Germaine Fuh, and Patrick Koenig. 2024. "An Engineered Mouse Model That Generates a Diverse Repertoire of Endogenous, High-Affinity Common Light Chain Antibodies" Antibodies 13, no. 1: 14. https://doi.org/10.3390/antib13010014
APA StyleRong, Y., Chen, I. -L., Larrabee, L., Sawant, M. S., Fuh, G., & Koenig, P. (2024). An Engineered Mouse Model That Generates a Diverse Repertoire of Endogenous, High-Affinity Common Light Chain Antibodies. Antibodies, 13(1), 14. https://doi.org/10.3390/antib13010014