Immunoglobulin for Treating Bacterial Infections: One More Mechanism of Action


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
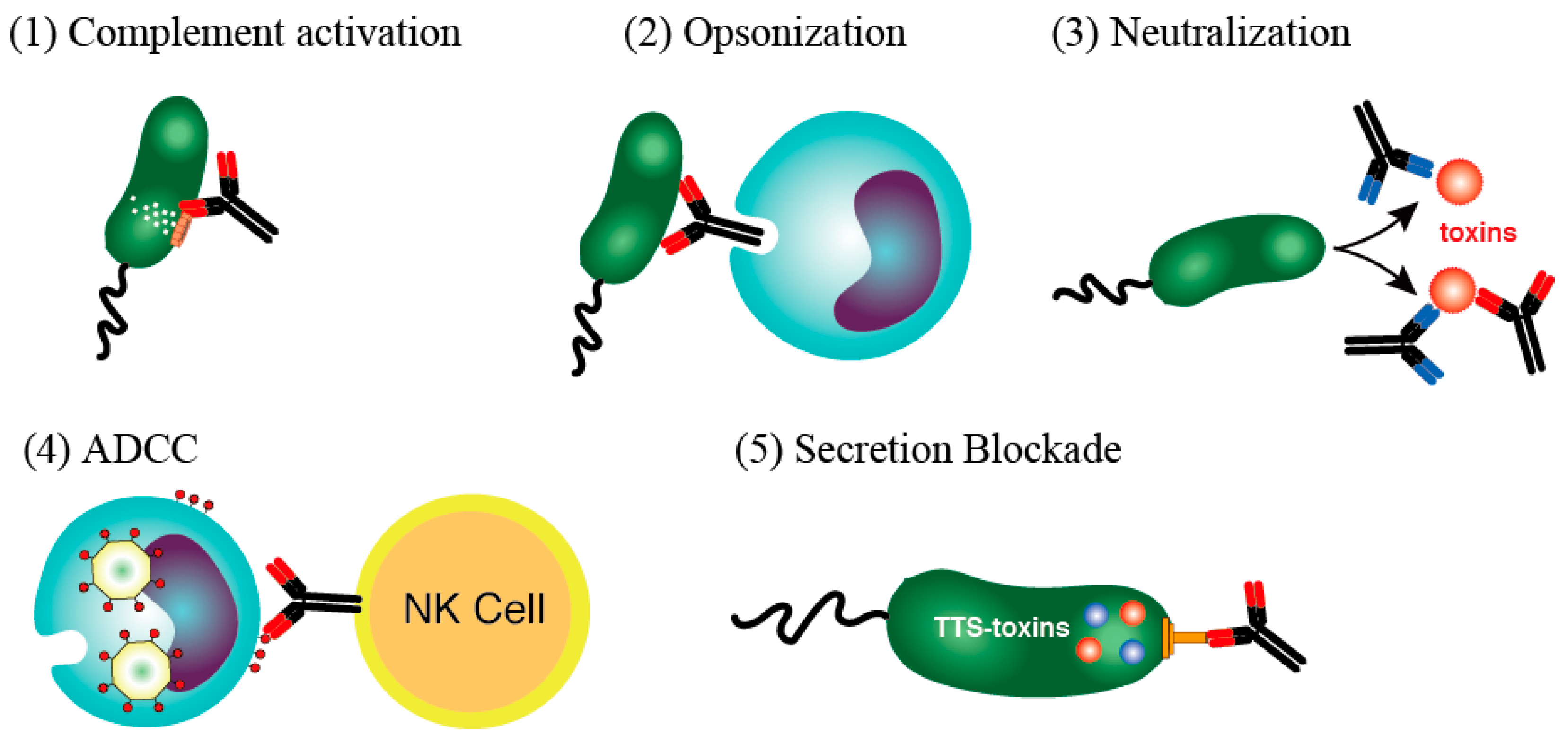
- (1)
- Complement-associated immunolysis: After the antibody binds to certain Gram-negative bacteria, spirochetes, or other types of bacteria, the complement components react, pierce the cell membrane, and destroy the bacteria through cell lysis.
- (2)
- Opsonin action (immunophagocytosis): Neutrophils and macrophages have a receptor for the Fc portion of IgG and can effectively phagocytose bacteria that are bound to IgG antibodies via this receptor. IgG bound to bacteria induces active oxygen (O2−) (a bactericidal substance) in the phagosome, which in turn acts directly on the bacterium as an oxidant and promotes phagosome to lysosome fusion. The phagocytosed bacteria are then effectively sterilized and digested by cooperation with lysosomal enzymes.
- (3)
- Toxin neutralizing action: An antibody responds by binding to a toxin produced by a bacterium and neutralizing its activity. In the case of a viral infection, the antibody can bind to a virus particle to prevent the virus from entering the target cell.
- (4)
- ADCC: When IgG antibodies bind to the virus-related antigens expressed on the surface of a virus-infected cell, a natural killer cell with Fc receptors can then bind to and damage the virus-infected cell. In doing so, it destroys the infected cell, which is the site of virus propagation, and prevents the transmission of pathogenic viruses.
2. The Bacterial Type III Secretion System and Its Toxins
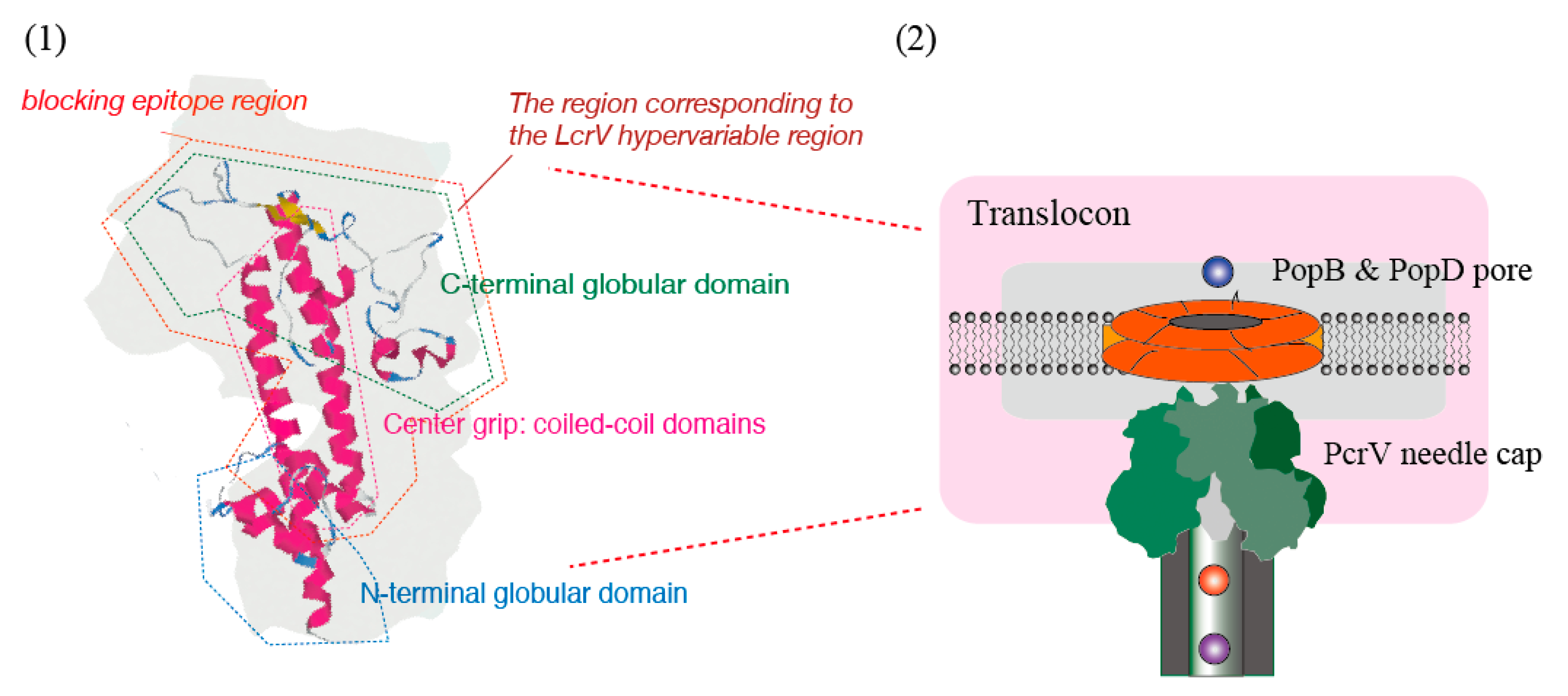
3. The V-Antigen of the Type III Secretion Apparatus and Its Specific Antibody
4. Blocking Effects of Antibodies Against the Bacterial Type III Secretion System
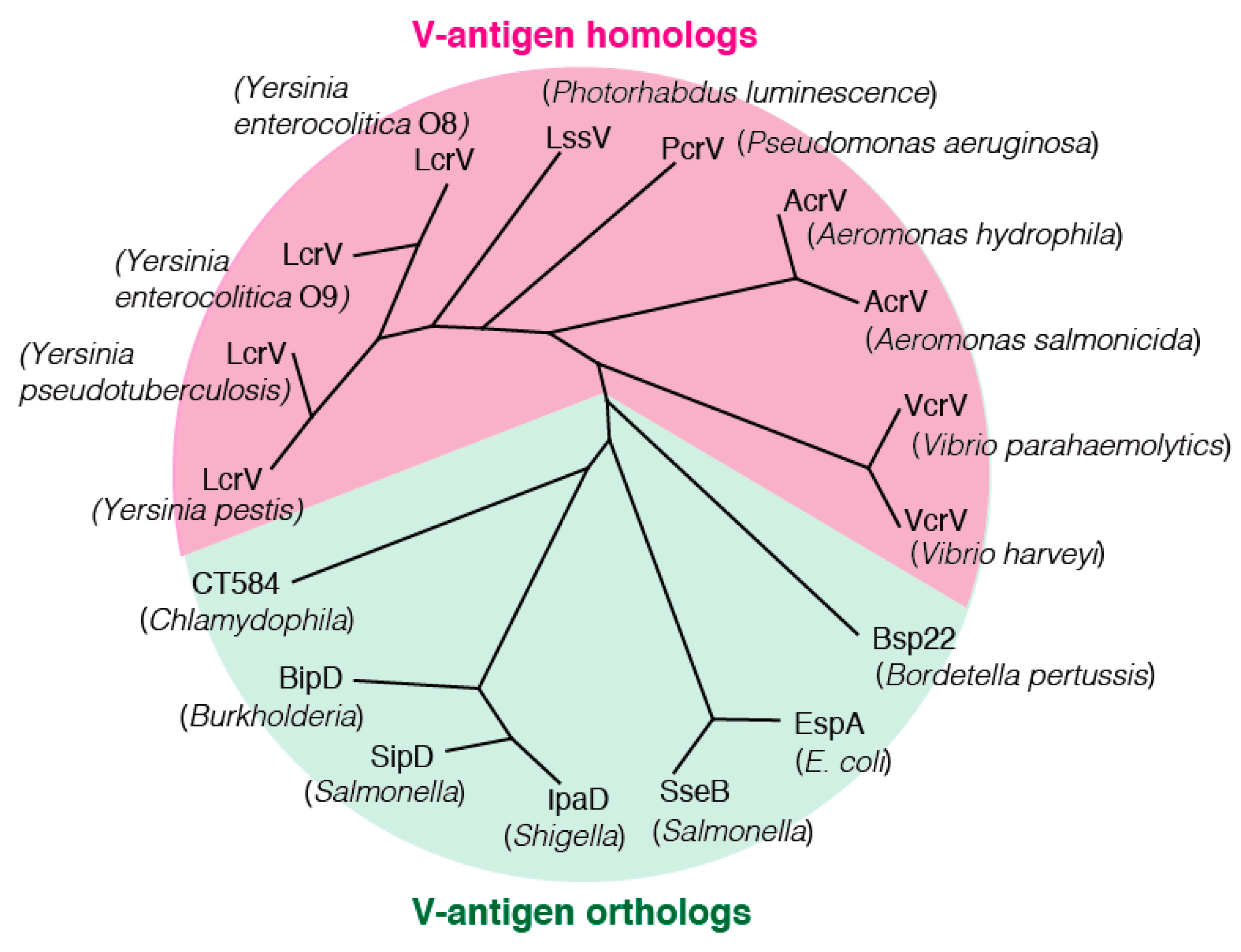
5. V-Antigen Homologs
6. The Antibody-Blocking Mechanisms against Type III Secretion
7. The Blocking Antibody Fraction in Human Serum
8. The Fifth Mechanism of Action of Immunoglobulin Therapy and Future Research
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Murphy, K.; Weaver, C. Basic concept of immunology. Antibodies protect against extracellular pathogens and their toxic products. The effector mechanisms of immunity. In Janeway’s Immunobiology, 9th ed.; Murphy, K., Weaver, C., Eds.; Taylor & Francis Group: New York, NY, USA, 2016; pp. 27–28. [Google Scholar]
- Laing, K. Immune Responses to Bacteria. Bitesized Immunlogy. British Society for Immunology. Available online: https://www.immunology.org/public-information/bitesized-immunology/patógenos-y-enfermedades/immune-responses-bacteria (accessed on 11 October 2019).
- Sewell, W.A.; Jolles, S. Immunomodulatory action of intravenous immunoglobulin. Immunology 2002, 107, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Vassilev, T.; Kazatchkine, M.D. Mechanisms of immunomodulatory action of intravenous immunoglobulin in autoimmune and systemic inflammatory diseases. Ther. Apher. 1997, 1, 38–41. [Google Scholar] [CrossRef] [PubMed]
- Zandman-Goddard, G.; Shoenfeld, Y. Intravenous immunoglobulin-customized therapy. Section IV Innovative Therapies 41. In Autoimmunity: From Bench to Bedside; Anata, J.-M., Shoenfeld, Y., Rosjas-Villarraga, A., Levy, R.A., Cervera, R., Eds.; El Rosario University Press: Bogota, Colombia, 2013; pp. 699–704. [Google Scholar]
- Afonso, A.F.B.; João, C.M.P. The production processes and biological effects of intravenous immunoglobulin. Biomolecules 2016, 6, 15. [Google Scholar] [CrossRef]
- Boros, P.; Gondolesi, G.; Bromberg, J.S. High dose intravenous immunoglobulin treatment: Mechanisms of action. Liver Transpl. 2005, 11, 1469–1480. [Google Scholar] [CrossRef] [PubMed]
- Chaigne, B.; Mouthon, L. Mechanisms of action of intravenous immunoglobulin. Transfus. Apher. Sci. 2017, 56, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Kazatchkine, M.D.; Bellon, B.; Kaveri, S.V. Mechanisms of action of intravenous immunoglobulin (IVIG). Mult. Scler. 2000, 6 (Suppl. 2), S24–S26. [Google Scholar] [CrossRef]
- Negi, V.S.; Elluru, S.; Siberil, S.; Graff-Dubois, S.; Mouthon, L.; Kazatchkine, M.D.; Lacroix-Desmazes, S.; Bayry, J.; Kaveri, S.V. Intravenous immunoglobulin: An update on the clinical use and mechanisms of action. J. Clin. Immunol. 2007, 27, 233–245. [Google Scholar] [CrossRef]
- Newland, A.C. The use and mechanisms of action of intravenous immunoglobulin: An update. Br. J. Haematol. 1989, 72, 301–305. [Google Scholar] [CrossRef]
- McCusker, C.; Warrington, R.R. Primary immunodeficiency. Allergy Asthma Clin. Immunol. 2011, 7, S11. [Google Scholar] [CrossRef]
- Parks, T.; Barrett, L.; Jones, N. Invasive Streptococcal disease: A review for clinicians. Br. Med. Bull. 2015, 115, 778–779. [Google Scholar] [CrossRef]
- Aubron, C.; Berteau, F.; Sparrow, R.L. Intravenous immunoglobulin for adjunctive treatment of severe infections in ICUs. Curr. Opin. Crit. Care 2019, 25, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Alejandria, M.M.; Lansang, M.A.; Dans, L.F.; Mantaring, J.B., 3rd. Intravenous immunoglobulin for treating sepsis, severe sepsis and septic shock. Cochrane Database Syst. Rev. 2013, 9, CD001090. [Google Scholar] [CrossRef]
- Anantharajah, A.; Mingeot-Leclercq, M.P.; Van Bambeke, F. Targeting the type three secretion system in Pseudomonas aeruginosa. Trends Pharmacol. Sci. 2016, 37, 734–749. [Google Scholar] [CrossRef] [PubMed]
- Fasciano, A.C.; Shaban, L.; Mecsas, J. Promises and challenges of the type three secretion system-injectisome as an anti-virulence target. EcoSal Plus 2019, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Rosqvist, R.; Magnusson, K.E.; Wolf-Watz, H. Target cell contact triggers expression and polarized transfer of Yersinia YopE cytotoxin into mammalian cells. EMBO J. 1994, 13, 964–972. [Google Scholar] [CrossRef] [PubMed]
- Rosqvist, R.; Persson, C.; Hakansson, S.; Nordfeldt, R.; Wolf-Watz, H. Translocation of the Yersinia YopE and YopH virulence proteins into target cells is mediated by YopB and YopD. Contrib. Microbiol. Immunol. 1995, 13, 230–234. [Google Scholar]
- Rosqvist, R.; Forsberg, A.; Wolf-Watz, H. Intracellular targeting of the Yersinia YopE cytotoxin in mammalian cells induces actin microfilament disruption. Infect. Immun. 1991, 59, 4562–4569. [Google Scholar]
- Cornelis, G.R. Type III secretion: A bacterial device for close combat with cells of their eukaryotic host. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2000, 355, 681–693. [Google Scholar] [CrossRef]
- Galan, J.E.; Collmer, A. Type III secretion machines: Bacterial devices for protein delivery into host cells. Science 1999, 284, 1322–1328. [Google Scholar] [CrossRef]
- Lee, C.A. Type III secretion systems: Machines to deliver bacterial proteins into eukaryotic cells? Trends Microbiol. 1997, 5, 148–156. [Google Scholar] [CrossRef]
- Hueck, C.J. Type III protein secretion systems in bacterial pathogens of animals and plants. Microbiol. Mol. Biol. Rev. 1998, 62, 379–433. [Google Scholar] [PubMed]
- Blocker, A.; Komoriya, K.; Aizawa, S. Type III secretion systems and bacterial flagella: Insights into their function from structural similarities. Proc. Natl. Acad. Sci. USA 2003, 100, 3027–3030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aizawa, S.I. Bacterial flagella and type III secretion systems. FEMS Microbiol. Lett. 2001, 202, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Christie, P.J. Type IV secretion: Intercellular transfer of macromolecules by systems ancestrally related to conjugation machines. Mol. Microbiol. 2001, 40, 294–305. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Coburn, J.; Collier, R.J. The eukaryotic host factor that activates exoenzyme S of Pseudomonas aeruginosa is a member of the 14-3-3 protein family. Proc. Natl. Acad. Sci. USA 1993, 90, 2320–2324. [Google Scholar] [CrossRef]
- Anderson, D.M.; Schmalzer, K.M.; Sato, H.; Casey, M.; Terhune, S.S.; Haas, A.L.; Feix, J.B.; Frank, D.W. Ubiquitin and ubiquitin-modified proteins activate the Pseudomonas aeruginosa T3SS cytotoxin, ExoU. Mol. Microbiol. 2011, 82, 1454–1467. [Google Scholar] [CrossRef]
- Lara-Tejero, M.; Galán, J.E. The injectisome, a complex nanomachine for protein injection into mammalian cells. EcoSal Plus 2019, 8, 1–18. [Google Scholar] [CrossRef]
- Dey, S.; Chakravarty, A.; Guha Biswas, P.; De Guzman, R.N. The type III secretion system needle, tip, and translocon. Protein Sci. 2019, 28, 1582–1593. [Google Scholar] [CrossRef]
- Bacon, G.A.; Burrows, T.W. The basis of virulence in Pasteurella pestis: An antigen determining virulence. Br. J. Exp. Pathol. 1956, 37, 481–493. [Google Scholar]
- Burrows, T.W. An antigen determining virulence in Pasteurella pestis. Nature 1956, 177, 426–427. [Google Scholar] [CrossRef]
- Burrows, T.W.; Bacon, G.A. The basis of virulence in Pasteurella pestis: The development of resistance to phagocytosis in vitro. Br. J. Exp. Pathol. 1956, 37, 286–299. [Google Scholar] [PubMed]
- Burrows, T.W. Virulence of Pasteurella pestis. Nature 1957, 179, 1246–1247. [Google Scholar] [CrossRef] [PubMed]
- Burrows, T.W.; Bacon, G.A. The effects of loss of different virulence determinants on the virulence and immunogenicity of strains of Pasteurella pestis. Br. J. Exp. Pathol. 1958, 39, 278–291. [Google Scholar] [PubMed]
- Burrows, T.W. Biochemical properties of virulent and avirulent strains of bacteria: Salmonella typhosa and Pasteurella pestis. Ann. NY Acad. Sci. 1960, 88, 1125–1135. [Google Scholar] [CrossRef]
- Perry, R.D.; Harmon, P.A.; Bowmer, W.S.; Straley, S.C. A low-Ca2+ response operon encodes the V antigen of Yersinia pestis. Infect. Immun. 1986, 54, 428–434. [Google Scholar]
- Goguen, J.D.; Yother, J.; Straley, S.C. Genetic analysis of the low calcium response in Yersinia pestis mu d1(Ap lac) insertion mutants. J. Bacteriol. 1984, 160, 842–848. [Google Scholar]
- Motin, V.L.; Nakajima, R.; Smirnov, G.B.; Brubaker, R.R. Passive immunity to yersiniae mediated by anti-recombinant V antigen and protein A-V antigen fusion peptide. Infect. Immun. 1994, 62, 4192–4201. [Google Scholar]
- Roggenkamp, A.; Geiger, A.M.; Leitritz, L.; Kessler, A.; Heesemann, J. Passive immunity to infection with Yersinia spp. mediated by anti-recombinant V antigen is dependent on polymorphism of V antigen. Infect. Immun. 1997, 65, 446–451. [Google Scholar]
- Pettersson, J.; Holmstrom, A.; Hill, J.; Leary, S.; Frithz-Lindsten, E.; von Euler-Matell, A.; Carlsson, E.; Titball, R.; Forsberg, A.; Wolf-Watz, H. The V-antigen of Yersinia is surface exposed before target cell contact and involved in virulence protein translocation. Mol. Microbiol. 1999, 32, 961–976. [Google Scholar] [CrossRef]
- Fields, K.A.; Nilles, M.L.; Cowan, C.; Straley, S.C. Virulence role of V antigen of Yersinia pestis at the bacterial surface. Infect. Immun. 1999, 67, 5395–5408. [Google Scholar]
- Yahr, T.L.; Goranson, J.; Frank, D.W. Exoenzyme S of Pseudomonas aeruginosa is secreted by a type III pathway. Mol. Microbiol. 1996, 22, 991–1003. [Google Scholar] [CrossRef] [PubMed]
- Finck-Barbancon, V.; Goranson, J.; Zhu, L.; Sawa, T.; Wiener-Kronish, J.P.; Fleiszig, S.M.; Wu, C.; Mende-Mueller, L.; Frank, D.W. ExoU expression by Pseudomonas aeruginosa correlates with acute cytotoxicity and epithelial injury. Mol. Microbiol. 1997, 25, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Frank, D.W. The exoenzyme S regulon of Pseudomonas aeruginosa. Mol. Microbiol. 1997, 26, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Yahr, T.L.; Mende-Mueller, L.M.; Friese, M.B.; Frank, D.W. Identification of type III secreted products of the Pseudomonas aeruginosa exoenzyme S regulon. J. Bacteriol. 1997, 179, 7165–7168. [Google Scholar] [CrossRef]
- Sawa, T.; Yahr, T.L.; Ohara, M.; Kurahashi, K.; Gropper, M.A.; Wiener-Kronish, J.P.; Frank, D.W. Active and passive immunization with the Pseudomonas V antigen protects against type III intoxication and lung injury. Nat. Med. 1999, 5, 392–398. [Google Scholar] [CrossRef]
- Hakansson, S.; Bergman, T.; Vanooteghem, J.C.; Cornelis, G.; Wolf-Watz, H. YopB and YopD constitute a novel class of Yersinia Yop proteins. Infect. Immun. 1993, 61, 71–80. [Google Scholar]
- Sarker, M.R.; Neyt, C.; Stainier, I.; Cornelis, G.R. The Yersinia Yop virulon: LcrV is required for extrusion of the translocators YopB and YopD. J. Bacteriol. 1998, 180, 1207–1214. [Google Scholar]
- Wager, B.; Faudry, E.; Wills, T.; Attree, I.; Delcour, A.H. Current fluctuation analysis of the PopB and PopD translocon components of the Pseudomonas aeruginosa type III secretion system. Biophys. J. 2013, 104, 1445–1455. [Google Scholar] [CrossRef]
- Tang, Y.; Romano, F.B.; Breña, M.; Heuck, A.P. The Pseudomonas aeruginosa type III secretion translocator PopB assists the insertion of the PopD translocator into host cell membranes. J. Biol. Chem. 2018, 293, 8982–8993. [Google Scholar] [CrossRef]
- Mueller, C.A.; Broz, P.; Muller, S.A.; Ringler, P.; Erne-Brand, F.; Sorg, I.; Kuhn, M.; Engel, A.; Cornelis, G.R. The V-antigen of Yersinia forms a distinct structure at the tip of injectisome needles. Science 2005, 310, 674–676. [Google Scholar] [CrossRef]
- Mota, L.J. Type III secretion gets an LcrV tip. Trends Microbiol. 2006, 14, 197–200. [Google Scholar] [CrossRef] [PubMed]
- Shime, N.; Sawa, T.; Fujimoto, J.; Faure, K.; Allmond, L.R.; Karaca, T.; Swanson, B.L.; Spack, E.G.; Wiener-Kronish, J.P. Therapeutic administration of anti-PcrV F(ab’) (2) in sepsis associated with Pseudomonas aeruginosa. J. Immunol. 2001, 167, 5880–5886. [Google Scholar] [CrossRef] [PubMed]
- Neely, A.N.; Holder, I.A.; Wiener-Kronish, J.P.; Sawa, T. Passive anti-PcrV treatment protects burned mice against Pseudomonas aeruginosa challenge. Burns 2005, 31, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Imamura, Y.; Yanagihara, K.; Fukuda, Y.; Kaneko, Y.; Seki, M.; Izumikawa, K.; Miyazaki, Y.; Hirakata, Y.; Sawa, T.; Wiener-Kronish, J.P.; et al. Effect of anti-PcrV antibody in a murine chronic airway Pseudomonas aeruginosa infection model. Eur. Respir. J. 2007, 29, 965–968. [Google Scholar] [CrossRef] [PubMed]
- Frank, D.W.; Vallis, A.; Wiener-Kronish, J.P.; Roy-Burman, A.; Spack, E.G.; Mullaney, B.P.; Megdoud, M.; Marks, J.D.; Fritz, R.; Sawa, T. Generation and characterization of a protective monoclonal antibody to Pseudomonas aeruginosa PcrV. J. Infect. Dis. 2002, 186, 64–73. [Google Scholar] [CrossRef]
- Faure, K.; Fujimoto, J.; Shimabukuro, D.W.; Ajayi, T.; Shime, N.; Moriyama, K.; Spack, E.G.; Wiener-Kronish, J.P.; Sawa, T. Effects of monoclonal anti-PcrV antibody on Pseudomonas aeruginosa-induced acute lung injury in a rat model. J. Immune Based Ther. Vaccines 2003, 1, 2. [Google Scholar] [CrossRef]
- Baer, M.; Sawa, T.; Flynn, P.; Luehrsen, K.; Martinez, D.; Wiener-Kronish, J.P.; Yarranton, G.; Bebbington, C. An engineered human antibody fab fragment specific for Pseudomonas aeruginosa PcrV antigen has potent antibacterial activity. Infect. Immun. 2009, 77, 1083–1090. [Google Scholar] [CrossRef]
- Francois, B.; Luyt, C.E.; Dugard, A.; Wolff, M.; Diehl, J.L.; Jaber, S.; Forel, J.M.; Garot, D.; Kipnis, E.; Mebazaa, A.; et al. Safety and pharmacokinetics of an anti-PcrV PEGylated monoclonal antibody fragment in mechanically ventilated patients colonized with Pseudomonas aeruginosa: A randomized, double-blind, placebo-controlled trial. Crit. Care Med. 2012, 40, 2320–2326. [Google Scholar] [CrossRef]
- Milla, C.E.; Chmiel, J.F.; Accurso, F.J.; VanDevanter, D.R.; Konstan, M.W.; Yarranton, G.; Geller, D.E.; Group, K.B.S. Anti-PcrV antibody in cystic fibrosis: A novel approach targeting Pseudomonas aeruginosa airway infection. Pediatr. Pulmonol. 2014, 49, 650–658. [Google Scholar] [CrossRef]
- Jain, R.; Beckett, V.V.; Konstan, M.W.; Accurso, F.J.; Burns, J.L.; Mayer-Hamblett, N.; Milla, C.; VanDevanter, D.R.; Chmiel, J.F.; Group, K.A.S. KB001-A, a novel anti-inflammatory, found to be safe and well-tolerated in cystic fibrosis patients infected with Pseudomonas aeruginosa. J. Cyst. Fibros. 2018, 17, 484–491. [Google Scholar] [CrossRef]
- Moriyama, K.; Wiener-Kronish, J.P.; Sawa, T. Protective effects of affinity-purified antibody and truncated vaccines against Pseudomonas aeruginosa V-antigen in neutropenic mice. Microbiol. Immunol. 2009, 53, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Hamaoka, S.; Naito, Y.; Katoh, H.; Shimizu, M.; Kinoshita, M.; Akiyama, K.; Kainuma, A.; Moriyama, K.; Ishii, K.J.; Sawa, T. Efficacy comparison of adjuvants in PcrV vaccine against Pseudomonas aeruginosa pneumonia. Microbiol. Immunol. 2017, 61, 64–74. [Google Scholar] [CrossRef]
- Naito, Y.; Hamaoka, S.; Kinoshita, M.; Kainuma, A.; Shimizu, M.; Katoh, H.; Moriyama, K.; Ishii, K.J.; Sawa, T. The protective effects of nasal PcrV-CpG oligonucleotide vaccination against Pseudomonas aeruginosa pneumonia. Microbiol. Immunol. 2018, 62, 774–785. [Google Scholar] [CrossRef] [PubMed]
- Thanabalasuriar, A.; Surewaard, B.G.; Willson, M.E.; Neupane, A.S.; Stover, C.K.; Warrener, P.; Wilson, G.; Keller, A.E.; Sellman, B.R.; DiGiandomenico, A.; et al. Bispecific antibody targets multiple Pseudomonas aeruginosa evasion mechanisms in the lung vasculature. J. Clin. Investig. 2017, 127, 2249–2261. [Google Scholar] [CrossRef] [PubMed]
- Tabor, D.E.; Oganesyan, V.; Keller, A.E.; Yu, L.; McLaughlin, R.E.; Song, E.; Warrener, P.; Rosenthal, K.; Esser, M.; Qi, Y.; et al. Pseudomonas aeruginosa PcrV and Psl, the molecular targets of bispecific antibody MEDI3902, are conserved among diverse global clinical isolates. J. Infect. Dis. 2018, 218, 1983–1994. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.O.; Yu, X.Q.; Robbie, G.J.; Wu, Y.; Shoemaker, K.; Yu, L.; DiGiandomenico, A.; Keller, A.E.; Anude, C.; Hernandez-Illas, M.; et al. Phase 1 study of MEDI3902, an investigational anti-Pseudomonas aeruginosa PcrV and Psl bispecific human monoclonal antibody, in healthy adults. Clin. Microbiol. Infect. 2019, 25, 629. e1–629. e6. [Google Scholar] [CrossRef]
- Le, H.N.; Tran, V.G.; Vu, T.T.T.; Gras, E.; Le, V.T.M.; Pinheiro, M.G.; Aguiar-Alves, F.; Schneider-Smith, E.; Carter, H.C.; Sellman, B.R.; et al. Treatment efficacy of MEDI3902 in Pseudomonas aeruginosa bloodstream infection and acute pneumonia rabbit models. Antimicrob. Agents Chemother. 2019, 63, 710–719. [Google Scholar] [CrossRef]
- Sawa, T.; Katoh, H.; Yasumoto, H. V-antigen homologs in pathogenic gram-negative bacteria. Microbiol. Immunol. 2014, 58, 267–285. [Google Scholar] [CrossRef]
- Burr, S.E.; Stuber, K.; Wahli, T.; Frey, J. Evidence for a type III secretion system in Aeromonas salmonicida subsp. salmonicida. J. Bacteriol. 2002, 184, 5966–5970. [Google Scholar] [CrossRef]
- Bergh, P.V.; Burr, S.E.; Benedicenti, O.; von Siebenthal, B.; Frey, J.; Wahli, T. Antigens of the type-three secretion system of Aeromonas salmonicida subsp. salmonicida prevent protective immunity in rainbow trout. Vaccine 2013, 31, 5256–5261. [Google Scholar] [CrossRef]
- Sekiya, K.; Ohishi, M.; Ogino, T.; Tamano, K.; Sasakawa, C.; Abe, A. Supermolecular structure of the enteropathogenic Escherichia coli type III secretion system and its direct interaction with the EspA-sheath-like structure. Proc. Natl. Acad. Sci. USA 2001, 98, 11638–11643. [Google Scholar] [CrossRef] [PubMed]
- Kenny, B.; Lai, L.C.; Finlay, B.B.; Donnenberg, M.S. EspA, a protein secreted by enteropathogenic Escherichia coli, is required to induce signals in epithelial cells. Mol. Microbiol. 1996, 20, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Becerra, F.J.; Chen, X.; Dickenson, N.E.; Choudhari, S.P.; Harrison, K.; Clements, J.D.; Picking, W.D.; Van De Verg, L.L.; Walker, R.I.; Picking, W.L. Characterization of a novel fusion protein from IpaB and IpaD of Shigella spp. and its potential as a pan-Shigella vaccine. Infect. Immun. 2013, 81, 447064477. [Google Scholar] [CrossRef] [PubMed]
- Sawa, T.; Ito, E.; Nguyen, V.H.; Haight, M. Anti-PcrV antibody strategies against virulent Pseudomonas aeruginosa. Hum. Vaccin. Immunother. 2014, 10, 2843–2852. [Google Scholar] [CrossRef] [PubMed]
- Derewenda, U.; Mateja, A.; Devedjiev, Y.; Routzahn, K.M.; Evdokimov, A.G.; Derewenda, Z.S.; Waugh, D.S. The structure of Yersinia pestis V-antigen, an essential virulence factor and mediator of immunity against plague. Structure 2004, 12, 301–306. [Google Scholar] [CrossRef]
- Gazi, A.D.; Charova, S.N.; Panopoulos, N.J.; Kokkinidis, M. Coiled-coils in type III secretion systems: Structural flexibility, disorder and biological implications. Cell Microbiol. 2009, 11, 719–729. [Google Scholar] [CrossRef]
- Mueller, C.A.; Broz, P.; Cornelis, G.R. The type III secretion system tip complex and translocon. Mol. Microbiol. 2008, 68, 1085–1095. [Google Scholar] [CrossRef]
- Broz, P.; Mueller, C.A.; Muller, S.A.; Philippsen, A.; Sorg, I.; Engel, A.; Cornelis, G.R. Function and molecular architecture of the Yersinia injectisome tip complex. Mol. Microbiol. 2007, 65, 1311–1320. [Google Scholar] [CrossRef]
- Sato, H.; Frank, D.W. Multi-functional characteristics of the Pseudomonas aeruginosa type III needle-tip protein, PcrV; comparison to orthologs in other gram-negative bacteria. Front. Microbiol. 2011, 2, 142. [Google Scholar] [CrossRef]
- Holmström, A.; Olsson, J.; Cherepanov, P.; Maier, E.; Nordfelth, R.; Pettersson, J.; Benz, R.; Wolf-Watz, H.; Forsberg, A. LcrV is a channel size-determining component of the Yop effector translocon of Yersinia. Mol. Microbiol. 2001, 39, 620–632. [Google Scholar] [CrossRef]
- Sing, A.; Tvardovskaia, N.; Rost, D.; Kirschning, C.J.; Wagner, H.; Heesemann, J. Contribution of toll-like receptor 2 and 4 in an oral Yersinia enterocolitica mouse infection model. Int. J. Med. Microbiol. 2003, 293, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Katoh, H.; Yasumoto, H.; Shimizu, M.; Hamaoka, S.; Kinoshita, M.; Akiyama, K.; Sawa, T. IV immunoglobulin for acute lung injury and bacteremia in Pseudomonas aeruginosa pneumonia. Crit. Care Med. 2016, 44, e12–e24. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Katoh, H.; Hamaoka, S.; Kinoshita, M.; Akiyama, K.; Naito, Y.; Sawa, T. Protective effects of intravenous immunoglobulin and antimicrobial agents on acute pneumonia in leukopenic mice. J. Infect. Chemother. 2016, 22, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, H.; Katoh, H.; Kinoshita, M.; Shimizu, M.; Hamaoka, S.; Akiyama, K.; Naito, Y.; Sawa, T. Epidemiological analysis of serum anti-Pseudomonas aeruginosa PcrV titers in adults. Microbiol. Immunol. 2016, 60, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, M.; Kato, H.; Yasumoto, H.; Shimizu, M.; Hamaoka, S.; Naito, Y.; Akiyama, K.; Moriyama, K.; Sawa, T. The prophylactic effects of human IgG derived from sera containing high anti-PcrV titers against pneumonia-causing Pseudomonas aeruginosa. Hum. Vaccin. Immunother. 2016, 12, 2833–2846. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sawa, T.; Kinoshita, M.; Inoue, K.; Ohara, J.; Moriyama, K. Immunoglobulin for Treating Bacterial Infections: One More Mechanism of Action. Antibodies 2019, 8, 52. https://doi.org/10.3390/antib8040052
Sawa T, Kinoshita M, Inoue K, Ohara J, Moriyama K. Immunoglobulin for Treating Bacterial Infections: One More Mechanism of Action. Antibodies. 2019; 8(4):52. https://doi.org/10.3390/antib8040052
Chicago/Turabian StyleSawa, Teiji, Mao Kinoshita, Keita Inoue, Junya Ohara, and Kiyoshi Moriyama. 2019. "Immunoglobulin for Treating Bacterial Infections: One More Mechanism of Action" Antibodies 8, no. 4: 52. https://doi.org/10.3390/antib8040052
APA StyleSawa, T., Kinoshita, M., Inoue, K., Ohara, J., & Moriyama, K. (2019). Immunoglobulin for Treating Bacterial Infections: One More Mechanism of Action. Antibodies, 8(4), 52. https://doi.org/10.3390/antib8040052